
Abstract
Recent studies have determined that gastrointestinal function contributes to the control of Parkinson's disease (PD). Gastrointestinal dysfunction results in a leaky intestinal barrier, inducing inflammation in the gut. Korean red ginseng (KRG) is widely used for the treatment of numerous afflictions, including inflammation and neurodegenerative disease. We investigated changes in the intestinal tight junctions and proinflammatory cytokines in the colon, and alpha-synuclein (aSyn) in the colon and the substantia nigra (SN) of a PD mouse model. Eight-week-old male C57BL/6 mice were intraperitoneally administered 30 mg/kg of 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) once a day for 5 days, and orally given 100 mg/kg of KRG for 12 consecutive days. Alterations in the levels of occludin, zonula occludens-1 (ZO-1), tumor necrosis factor-alpha (TNF-α) and interleukin-1 beta (IL-1β) in the colon, and the expressions of aSyn and tyrosine hydroxylase (TH) in the colon and the SN were evaluated. Oral administration of KRG significantly prevents the MPTP-induced motor dysfunction, and suppresses the MPTP-induced disruption of occludin and ZO-1, and suppresses the increase in TNF-α and IL-1β in the colon of mice. In addition, KRG prevents accumulation of aSyn and TH in the colon and the SN. These results suggest that KRG has the potential to prevent MPTP-induced leaky gut barrier, inflammation, and accumulation of aSyn.
Introduction
Parkinson's disease (PD) is a neurodegenerative disease characterized by the loss of dopaminergic neurons in the putamen, striatum, and substantia nigra (SN). Recently, many researchers have focused on the relationship between the enteric nervous system (ENS) and gastrointestinal (GI) tract in PD. GI dysfunction can be caused by “leaky gut,” a condition attributed to a collapsed intestinal barrier. 1,2 Occurrence of the leaky gut results in small molecular metabolic components being translocated from the gut epithelial barrier to the blood–brain barrier. 3 Occludin, claudins, and zonula occludens-1 (ZO-1) are tight junction proteins and regulate the epithelial permeability; hence, the expressions of these proteins reflect the epithelial integrity and function in the colon. 4 Disruption of tight junctions and intestinal inflammation leads to neuroinflammation or systemic inflammation, initiating PD. 5,6 The inflammation increases proinflammatory cytokines, which cause neuronal death in the brain of PD patients. 7
Although alpha-synuclein (aSyn) in the synaptic membrane is involved in neuronal communication, 8 the overproduction of aSyn and the accumulation of misfolded aSyn in Lewy bodies contribute to the pathogenesis of PD, 9 Moreover, dysfunction of the GI system induces the expression of aSyn. Stolzenberg et al. reported the presence of aSyn in intestines with GI inflammation, 10 and Fitzgerald et al. reported that microbiota in intestines aggregate aSyn, which is transported to the CNS via the vagus nerve, thereby contributing to PD. 11 These results indicate that an increase of aSyn in the intestine effectually increases the levels of aSyn in the brain; hence, the intestinal aSyn levels may be used as an alternative biomarker to diagnose PD. 12,13
Korean red ginseng (KRG) is a popular traditional herb in Asia. KRG is the steamed and dried root of Panax ginseng Meyer, it can be preserved for a long time, and is known to have antiapoptotic and anti-inflammatory effects. 14 Recent studies indicate that KRG prevents the loss of dopaminergic neurons in in vivo 15 –17 and in vitro 18 PD models. The neurotoxin 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP), commonly used for inducing PD in animal studies, is known to alter the gut motility associated with dopaminergic degeneration in the ENS, impair the intestine tight junction, 4 and increase aSyn levels in the colon. 19,20 However, it remains unknown whether KRG can prevent the MPTP-induced impairment of tight junctions and increase in aSyn in the colon. This study therefore focused on the changes of tight junctions and aSyn expressions in the colon, and investigated the protective effect of KRG in the MPTP-treated PD animal model.
Materials and Methods
Animals
Male 8-week-old C57BL/6 mice (18–21 g, Orient Bio, Inc., Seongnam, Korea) were housed at 22°C ± 2°C under a standard 12 h light/dark cycle and fed ad libitum. The animal study was approved by the Pusan National University Institutional Animal Care and Use Committee (PNU-IACUC, Approval No. PNU-2016-1071) and mice were handled in accordance with the current guidelines established by the PNU-IACUC. Mice were randomly assigned to four groups (n = 4–6): control group, MPTP group, MPTP+KRG group, and KRG group.
MPTP and KRG treatment
After an adaptation period of 1 week, mice in the MPTP and the MPTP+KRG groups were intraperitoneally administered MPTP-HCl (30 mg/kg; Sigma, St. Louis, MO, USA) at 24 h intervals for 5 days. Mice in the control and the KRG groups were intraperitoneally administered sterilized normal saline instead of MPTP. Two hours after the first MPTP or saline injection, mice in the MPTP+KRG and KRG groups were orally administrated 100 mg/kg of KRG (Korea Ginseng Corporation, Daejeon, Korea) at 24 h intervals for 12 consecutive days. In our previous study, this dose showed the best effect in MPTP-treated mice. 16
Pole test
The pole test was conducted on day 0 (1 day before the first MPTP injection) and day 12 (after the final KRG treatment). Mice were located near the top of a wooden square pole (55 cm in height and 1 cm in diameter), and the time taken to arrive at the bottom of the pole was measured. After repeating the test three times, the mean time was used in the final analysis. 21
Rotarod test
Mice were placed, facing forward, on a rotating cylinder with a bumpy surface (Harvard Apparatus, Holliston, MA, USA) and the time for which the mice stayed on the cylinder was measured. The start speed was 4 RPM, which was accelerated to 40 RPM for 2 min, with a cutoff time of 3 min. The test was repeated three times, maintaining an interval of at least 30 min between each trial. After 3 days of training, the test was conducted on day 0, day 5 (the last day of MPTP injection), and day 12.
Immunofluorescence analysis
After euthanasia, the colon lengths of the mice were measured and colon tissues were fixed and the frozen sections (4 μm thickness) were prepared on a cryostat (Leica Microsystems, Wetzlar, Germany). The sections were incubated with primary antibody occludin (1:200; Santa Cruz Biotechnology, Santa Cruz, CA, USA) and subsequently reacted with the secondary antibody (Alexa-488 IgG; Molecular Probes, Eugene, OR, USA) for 1.5 h at RT in the dark. The sections stained were mounted with VECTASHIELD (Vector, Burlingame, CA, USA), and a LSM 700 confocal laser scanning microscope (Zeiss, Oberkochen, Germany) was used to detect the fluorescence intensity.
Western blot analysis
Brain and colon tissues were subjected to western blot analysis after lysis with RIPA lysis buffer (LPS Solution, Daejeon, Korea) containing 1 mM phenylmethanesulfonyl fluoride (Enzo Life Sciences, Farmingdale, NY, USA). After the separation of total protein (40 μg by 10% or 12% sodium dodecyl sulfate–polyacrylamide gel electrophoresis), it was transferred to a 0.2 μm nitrocellulose blotting membrane (GE Health care UK Ltd., Little Chalfont, UK). The membrane was incubated with primary antibodies: aSyn (1:500), interleukin-1 beta (IL-1β; 1:500), occludin (1:500) and tyrosine hydroxylase (TH; 1:500; Santa Cruz biotechnology), tumor necrosis factor-alpha (TNF-α; 1:1000; Cell Signaling Technology, Beverly, MA, USA), and ZO-1 (1:1000; Abcam, Cambridge, UK). The probed membranes were subsequently incubated with secondary antibody and washed with TBS-T three times. The protein bands were detected by Image Quant LAS 4000 (Fujifilm, Tokyo, Japan). The protein expression levels were quantified by ImageJ.
Statistical analysis
All data are presented as means ± standard deviations. Behavior tests were analyzed by two-way analysis of variance (ANOVA) with Bonferroni post hoc test. The data from immunofluorescence and western blot were analyzed by one-way ANOVA with Neuman–Keuls post hoc test. All statistical analyses were conducted using Prism 5 for Windows (GraphPad Software, Inc., La Jolla, CA, USA), and the differences are considered statistically significant at P < .05.
Results
KRG improves motor function of MPTP-injected mice
The pole test was performed on days 0 and 12. On day 0, no significant differences were observed between groups, for the time taken to arrive at the bottom. On day 12, the time in the MPTP group showed significantly increased compared to that in the control group (P < .05), and the time in the MPTP+KRG group was significantly decreased compared to the MPTP group (P < .001, Fig. 1A).

Effect of KRG on MPTP-induced behavior dysfunction.
The rotarod test was performed on days 0, 5, and 12. Before MPTP administration, no significant differences were observed among the groups, in the time for which the mice stayed on the cylinder. On day 5, the times in all the groups were not significantly different, although the mice in all groups stayed longer than on day 0. On day 12, the mice in all groups stayed longer than on day 5, and the time in the MPTP+KRG group stayed significantly longer than that in the MPTP group (P < .05, Fig. 1B).
KRG prevents MPTP-induced disruption of tight junction in the colon
Occludin, the main component in tight junctions, is important for functioning of the intestinal barrier and tight junction stability. 22 ZO-1, one of the tight junction-associated proteins, is responsible for linking the transmembrane protein and actin. 23 Occludin and ZO-1 regulate the permeability of epithelial cells in the intestine. In the present study, occludin immunofluorescence staining demonstrated that MPTP significantly decreases the expression of occludin in the colon (P < .001), and KRG administration significantly alleviates the decrease (P < .01, Fig. 2A and Supplementary Fig. S1). These alterations were further confirmed by western blotting. MPTP caused a significant decrease in ZO-1 expression in the colon (P < .05), and KRG partially alleviated the MPTP-induced decrease, but not significantly (Fig. 2B).

Effect of KRG on MPTP-induced disruption of intestinal tight junction in the colon.
KRG regulates inflammatory responses in the colon of MPTP-injected mice
To confirm the occurrence of intestinal inflammation, we evaluated the levels of TNF-α and IL-1β in the colon by western blotting and the length of the colon. MPTP administration significantly increased the expressions of TNF-α (P < .01) and IL-1β (P < .01) in the colon, and KRG administration significantly suppressed the MPTP-induced increase in TNF-α (P < .01) and IL-1β (P < .001). The length of the colon in the MPTP group was significantly shorter than that in the control group (P < .05), however, the length in the MPTP+KRG group was not significantly different from that in the control group (Fig. 3 and Supplementary Fig. S2).

Effect of KRG on MPTP-induced increase of proinflammatory cytokines in the colon. MPTP induces an increase in the levels of TNF-α and IL-1β in the colon and a decrease in the colon length, which are suppressed by the KRG. Data are presented as the means ± standard deviations (n = 4–6). *P < .05 and **P < .01 compared with the control group. ## P < .01 and ### P < .001 compared with the MPTP group. IL-1β, interleukin-1 beta; TNF-α, tumor necrosis factor-alpha.
KRG regulates the expressions of TH and aSyn in the colon and the SN
Intestinal barrier dysfunction induces the increase of intestinal permeability and the loss of enteric dopaminergic neurons, mostly in the myenteric plexus in the ENS. 20,24 In addition, the microbiota in the colon play a role in the formation of aSyn in both ENS and CNS in PD. 11 In this study, MPTP significantly decreases the expression of TH in the colon (P < .001), whereas KRG administration alleviated it (P < .05). Similar expressions are observed in the SN, after MPTP administration (P < .01). Furthermore, exposure to MPTP increases aSyn expression in the colon (P < .001) and the SN (P < .001), which are significantly suppressed by KRG in the colon (P < .001) and the SN (P < .001, Fig. 4 and Supplementary Fig. S2). These results suggest that the SN and colon have a similar tendency for regulating the expressions of TH and aSyn.
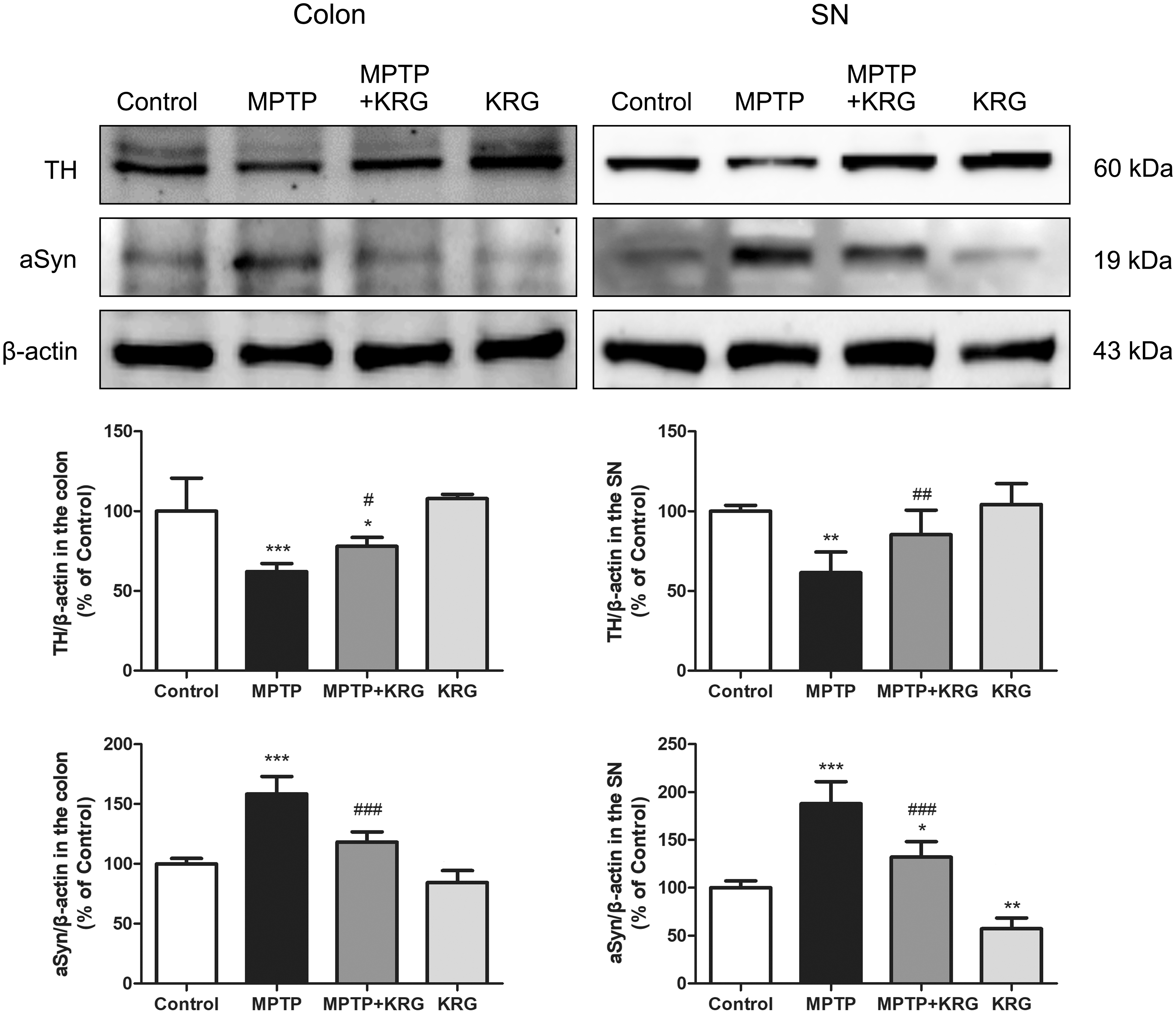
Effect of KRG on MPTP-induced suppression of TH and aSyn in the colon and the SN. MPTP suppresses the TH levels and increases aSyn levels in the colon and SN, and KRG exposure restores to the original levels. Data are presented as the means ± standard deviations (n = 4–6). *P < .05, **P < .01, and ***P < .001 compared with the control group. # P < .05, ## P < .01, and ### P < .001 compared with the MPTP group. aSyn, alpha-synuclein; SN, substantia nigra; TH, tyrosine hydroxylase.
Discussion
The present study demonstrates that the administration of KRG alleviates the MPTP-induced behavioral impairment, protects against the damage of tight junctions in the colon, and suppresses the increase in TNF-α, IL-1β, and aSyn in the colon and SN.
The loss of dopaminergic neurons in the SN, the deposition of Lewy bodies, and the accumulation of aSyn are the characteristics of PD. The main symptoms of PD include motor dysfunctions such as tremors, rigidity, and slow movement. In this study, we used the pole and the rotarod tests to confirm whether KRG alleviates the motor dysfunctions in the PD animal model. The pole test is a simple method for evaluating motor functions and is widely applied for PD rodent studies, 25 whereas the rotarod test evaluates the motor coordination, grip strength, and balance of rodents. 26 Previous studies have reported that KRG prevents the MPTP-induced motor dysfunction, 15 –17 and the current study similarly confirms the outcome.
The intestinal barrier absorbs nutrients and water from the lumen and prevents the entry of toxic luminal substances and pathogenic microorganisms. The functions of intestinal barrier are preserved by a monolayer and a mucus layer of epithelial cells, and these cells are interconnected by tight junctions. 27 Tight junctions are multiprotein complexes comprising transmembrane proteins, including occludin, ZO-1, and claudin, and play an important role in the intestinal barrier and regulation of epithelial permeability. 4,28 Disruption of the intestinal tissue or exposure of substances provoking strong immune reaction induces inflammation, and consequently weaken the intestinal barrier and the tight junctions. 5 Disruption of the tight junctions leads to neuro-inflammation or systemic inflammation initiating PD. 5,6 Hence, protecting the integrity of the tight junctions possibly prevents inflammatory reactions in the GI tract of PD patients. In this study, MPTP administration induced a reduction in the levels of occludin and ZO-1 and increases the expressions of TNF-α and IL-1β in the colon. However, KRG exposure suppresses the MPTP-induced changes, thereby indicating that KRG prevents the MPTP-induced increase of proinflammatory cytokines and destruction of the tight junctions in the colon.
Ginseng and its components have been known to suppress intestinal damage and inflammation in the colitis animal models. Fermented ginseng protects intestinal epithelial barrier and inhibits the activation of proinflammatory cytokines and nuclear factor (NF)-κB in the colon of dextran sulfate sodium salt (DSS)-treated mouse, 29 red ginseng suppresses crypt destruction, surface epithelial loss, and inflammatory cell infiltration in the colon of DSS-treated mouse 30 and trinitro-benzene-sulfonic acid (TNBS)-treated rat, 31 and red ginseng oil reduces proinflammatory cytokines through inhibiting the NF-κB pathway in the colon of DSS-treated mouse. 32 Ginsenoside Rg1 reduces colonic damage and proinflammatory cytokines in the colon of DSS 33 or TNBS-treated mice, 34 ginsenoside Rb1 alleviates colon shortening, myeloperoxidase activity, and proinflammatory cytokines by blocking IRAK1 activation in the colon of TNBS-treated mice, 35,36 ginsenoside Re suppresses inflammation by inhibiting the binding of lipopolysaccharide to TLR4 on macrophages, 37 and ginsenoside Rk3 protects colon barrier and inhibits NLRP3 inflammasome expression in the colon of DSS-treated mice. 38 Moreover, ginseng oligopeptides protect intestinal barrier against irradiation by suppressing inflammation. 39 Taken together, the effect of KRG against inflammation and damage of the tight junctions in the colon may be due to the multiple action of various ingredients, including ginsenosides and oligopeptides in KRG.
Abnormal gut barrier and inflammatory bacterial products trigger an inflammatory response in the gut, which initiates aSyn accumulation in the ENS, 40 which, in turn, induces GI dysfunction. 41,42 This is commonly observed in PD patients afflicted with GI dysfunction. Moreover, inflammation in the colon elicits increased expression of aSyn in the brain as well as the gut. 6 The accumulated aSyn in the colon is transported to the CNS via the vagus nerve, 11 which consequently increases aSyn in the brain. In this study, we found that MPTP administration increases the levels of aSyn in the colon and the SN, and exposure to KRG significantly prevents the increase of aSyn; this may be due to the suppression of inflammation and the protection of tight junctions in the colon by KRG.
The TH enzyme is involved in the synthesis of dopamine. In the CNS, TH is abundant in the dopaminergic neurons, and is often used as a marker of dopaminergic neurons. In the ENS, dopamine contributes to GI motility; therefore, a reduction of dopamine in the intestine causes GI dysfunction. 20 In this study, MPTP suppressed the expression of TH in the colon and the SN, which is restored after exposure to KRG, indicating that MPTP induces GI symptoms (such as constipation) and dopaminergic neuronal destruction in the SN, but KRG subsequently alleviates them. However, changes in GI dysfunction were not investigated in the present study, and further studies are required to confirm this hypothesis.
In conclusion, our study determines that KRG alleviates the MPTP-induced motor dysfunction and increases in aSyn expression in SN, by preventing leakage of tight junctions and inflammation in the colon. These results indicate that KRG is a potential therapeutic for PD patients.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This research was supported by the Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the MSIT (No. NRF-2019R1A2C1085130).
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.