
Abstract
The aim of this study was to investigate the effect of a high-fat diet (HFD) on energy substrate utilization during long-term endurance exercise in mice. Male ICR mice (n = 32; 6 weeks old) were divided into two groups: low-fat diet (LFD, n = 16) and HFD (n = 16) and acclimatized to LFD or HFD feeding over 12 weeks. After 12 weeks, the two dietary groups were each divided into two groups with or without exercise (EX): LF-CON, LF-EX, HF-CON, and HF-EX groups. The exercise groups were trained to run on a treadmill for 12 weeks. At the end of the experimental protocol, energy metabolism in the whole body was measured at rest for 24 h and during exercise for 1 h using respiratory gas analysis. Furthermore, molecules involved in skeletal muscle fat metabolism were analyzed. Substrate utilization for energy metabolism in the whole body indicated that fat utilization was high in HFD intake. Notably, when HFD intake and exercise were combined, fat utilization was markedly increased during endurance exercise. In contrast, exercise showed no effect when combined with LFD intake. The gene expressions of Fat/Cd36, Fatp1, Fabp-pm, and Cpt1 were upregulated by HFD intake, with Fat/Cd36 and Cpt1 considerably elevated during long-term endurance exercise. In contrast, exercise showed no effect when combined with LFD intake. These results suggest that HFD intake effectively increased fat utilization as an energy substrate during long-term endurance exercise.
Introduction
During physical activity, the skeletal muscle uses the highest amount of energy, with fats and carbohydrates serving as the major fueling substrates for energy production. 1,2 Glycogen is a major energy substrate that is used during high-intensity physical activity. However, the accumulation of glycogen in the skeletal muscle is prevented by the efficient feedback-mediated inhibition of glycogen synthase, leading to rapid depletion and fatigue. 3 –5 Notably, 92–98% of the total endogenously stored energy is stored in the form of fat. 6,7 In energy metabolism, when fat is preferentially used as an energy source over glycogen, depletion of muscle glycogen is prevented. Furthermore, fat may generate a larger amount of energy to serve as a long-term source. Hence, it is crucial to increase fat utilization as an energy substrate to sustain long-term physical activity.
During fat oxidation in skeletal muscle, circulating long-chain fatty acids (LCFAs) primarily enter the cells through plasma membrane transporters, such as fatty acid translocase/cluster of differentiation 36 (FAT/CD36), plasma membrane-associated fatty acid binding protein (FABP-pm), and fatty acid transport protein 1 (FATP1). 8 –10 LCFAs are subsequently transported to the inner mitochondria through the carnitine system (CPT1/CACT/CPT2) and oxidized using the β-oxidation pathway, resulting in energy production. 11 These transporters, along with carnitine palmitoyltransferase 1 (CPT1), play key roles in fatty acid oxidation in skeletal muscle. 12
Several studies have shown that mRNA or/and protein expression of FAT/CD36 and CPT1 in the human skeletal muscle increases after short-term training on a cycle ergometer with a normal diet. 13,14 During endurance exercise, fatty acid transporters may be enhanced to facilitate the entry of fat into the skeletal muscle cells for fatty acid oxidation. Reportedly, the mRNA or/and protein levels of FAT/CD36 in the skeletal muscles of rats and humans increased with short-term feeding of a high-fat diet (HFD). 15,16 These studies demonstrated that the molecules involved in energy metabolism may be regulated by diet and exercise.
Researchers have proposed several dietary protocols to alter the patterns of energy substrate utilization, increasing the rate of fatty acid utilization and, thereby, reducing the rate of glycogen utilization during endurance exercise. Previously, studies have shown that short-term changes in diet, exercise, or both can produce changes in the molecules associated with fat metabolism. 13 –18 However, there is insufficient evidence available to confirm that changes in fat metabolism can be induced by long-term changes in diet or/and exercise.
Thus, we investigated the effect of a HFD on energy substrate utilization during long-term endurance exercise in mice. In the present study, mice were acclimatized to a high-fat/low-carbohydrate or low-fat/high-carbohydrate diet for 12 weeks and were then trained to run on a treadmill for 12 weeks while maintaining the same diet. We then used an open-circuit calorimetry system to measure whole-body energy expenditure and substrate utilization for mice resting for 24 h and for mice exercising for 1 h to determine the effect of a HFD on fat utilization during long-term endurance training. Furthermore, we also analyzed the gene expression related to fatty acid transport in the skeletal muscle.
Materials and Methods
Animals and diets
Thirty-two male ICR mice (6 weeks old) were purchased from Orient Bio Inc., (Seongnam, Republic of Korea). The mice were individually housed in standard cages, maintained on a 12-h light/12-h dark cycle at 23°C ± 1°C with 50% humidity, and allowed free access to food and water. After acclimation for 1 week, the mice were randomly divided into two groups: low-fat diet (LFD, n = 16) and HFD (n = 16) groups. The LFD group was fed a diet in which 10% of calories were obtained from fat and 70% of calories were obtained from carbohydrate (D12450B; Research Diets, Inc., USA). The HFD group was fed a diet in which 60% of calories were obtained from fat and 20% of calories were obtained from carbohydrate (D12492; Research Diets, Inc.,). After acclimation to each diet for a 12-week period, the mice were divided into two groups each with the inclusion of exercise (EX): the LF-CON (n = 8, low-fat diet), LF-EX (n = 8, low-fat diet+EX), HF-CON (n = 8, high-fat diet), and HF-EX (n = 8, high-fat diet+EX) groups. All mice were maintained on the same diet, and the exercise groups were trained by running on a treadmill for 12 weeks. The body weight and food intake were monitored daily throughout the experimental period. After 24 weeks, mice were sacrificed on the day after the end of the experimental protocol. All experimental protocols were approved by the Animal Care and Use Committee of the Konkuk University and conducted in accordance with the ethical guidelines at the Animal Experiment Research Center (Approval No. KU13031).
Running training
The mice in the exercise groups were trained by running on a treadmill. Running mice were adapted to treadmill training at a fixed intensity (10 m/min speed, 8° slope) for 5 days. Then all mice were run five times per week for 12 weeks using the following protocol: week 1: 8° slope, 10 m/min speed, 30 min/day running; weeks 2 and 3: 8°–10° slope, 12 m/min speed, 40 min/day running; weeks 4 and 5: 11° slope, 13 m/min speed, 45 min/day running; weeks 6 and 7: 12° slope, 14 m/min speed, 50 min/day running; and weeks 8–12: 12° slope, 15 m/min speed, 60 min/day running.
Respiratory gas analysis
Energy metabolism at resting
At the end of the experimental protocol, the resting metabolic rate was measured by indirect calorimetry, using an open-circuit apparatus as previously described. 19 –22 The flow rate was kept constant at 1.2 L/min and measured for 24 h. Mice had free access to food and water. O2 uptake and CO2 production were analyzed using a mass spectrometer (RL-600; Alco System, Chiba, Japan) and switching system (ANI6-A-S; Alco System). O2 uptake and CO2 production were used to calculate the respiratory exchange ratio (RER, VCO2/VO2) and fat and carbohydrate oxidation.
Energy metabolism during exercise
After 12 weeks of endurance training, energy metabolism was measured during 1 h of running using an exercise metabolism chamber 20 under the training conditions of 15 m/min and a 12° slope.
Muscle tissue collection
The mice were sacrificed on the day following the end of the protocol after 24 weeks. The gastrocnemius muscle was surgically removed, frozen in liquid nitrogen, and stored at −80°C for subsequent total RNA extraction.
Reverse transcription–polymerase chain reaction
Total RNA was extracted from 20 mg frozen muscle using TRIzol reagent (Ambion Inc., Austin, TX, USA), and the concentration was quantified using a spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA). First-strand complementary DNA (cDNA) was synthesized from 1 μg total RNA using amfiRivert cDNA Synthesis Platinum Master Mix (GenDEPOT, CA, USA) and used as a template. The cDNA was stored at −20°C for subsequent analysis. Primers for each gene were designed from gene sequences obtained from GenBank (National Center for Biotechnology Information). A BLAST search for each primer confirmed the homologous binding to other mRNAs of the skeletal muscle. Gene expression levels of Fat/Cd36, Fatp1, Fabp-pm, Cpt1, and Gapdh were measured by reverse transcription–polymerase chain reaction (RT-PCR) using the following primers: Fat/Cd36 forward 5′-ATGGGCTGTGATCGGAACTG-3′, reverse 5′-TGTCTGTACACAGTGGTGCC-3′; Fatp1 forward 5′-TCCTAAGGCTGCCATTGTGG-3′, reverse 5′-ACACAGTCATCCCAGAAGCG-3′; Fabp-pm forward 5′-CACAACTGGCAGCACATCAC-3′, reverse 5′-TTCCAGATACCAGCCGAGGA-3′; Cpt1 forward 5′-TAGGCCTCAACACCGAACAC-3′, reverse 5′-TGCCTTGGCTACTTGGTACG-3′; and Gapdh forward 5′-GACCACAGTCCATGCCATCA-3′, reverse 5′-CTCAGATGCCTGCTTCACCA-3′. The amplified PCR products of the target mRNA were then separated by 1% agarose gel electrophoresis and visualized by ethidium bromide staining. The gels were then photographed with a Molecular Imager Gel Doc™ XR System (Bio-Rad Laboratories, Inc., Hercules, CA, USA), and the product bands were quantified by ImageJ software (National Institutes of Health, Bethesda, MD, USA). The mRNA levels were normalized to the Gapdh mRNA level.
Statistical analyses
All data are presented as means ± standard deviations (SD). Effect of diet (LF vs. HF), exercise (EX vs. non-EX), and diet-by-exercise interaction were analyzed using two-way ANOVA. A comparison among the four groups was performed using one-way ANOVA. For all analyses, the values were considered significantly different at P < .05. The data were analyzed using JMP version 5 (SAS Institute Inc., USA).
Results
Changes in body weight and food intake
Changes in body weight and food intake in the four groups are shown in Table 1. During the training period, the daily energy intakes of the HFD (HF-CON and HF-EX) fed groups were significantly higher than the LFD (LF-CON and LF-EX) fed groups (P < .0001). Compared with each control, the HF-EX group demonstrated no significant difference in food intake, whereas the LF-EX group reported a higher food intake (Table 1).
Changes in Body Weight and Food Intake
Initial body weight indicates the weight before the start of the exercise program, after 12 weeks of a LFD or HFD. Values are shown as means ± SD. Values labeled with different superscript letters differ significantly among the four groups by one-way ANOVA (P < .05).
HFD, high-fat diet; LFD, low-fat diet; SD, standard deviation.
The final body weight differed significantly between the LFD and HFD fed groups (diet effect, P < .0001). Furthermore, an exercise-related effect (EX [LF-EX and HF-EX] vs. non-EX [LF-CON and HF-CON]) was observed (P = .0301), and a diet-by-exercise interaction was not significantly different for the final body weight (P = .1493) (Fig. 1). The final body weight of the two HFD groups, HF-CON and HF-EX (76.2 ± 2.2 g and 67.4 ± 2.0 g, respectively) differed significantly; however, this was not observed in the two LFD groups, LF-CON and LF-EX (56.3 ± 2.4 g and 54.4 ± 2.7 g, respectively). Exercise training exerted a significant effect on the final body weight in the HFD fed groups, but no statistically significant effects were observed in the LFD fed groups (Fig. 1). In addition, although the LF-EX group demonstrated a higher food intake than the LF-CON group, the observed loss in body weight verified the effect of exercise.
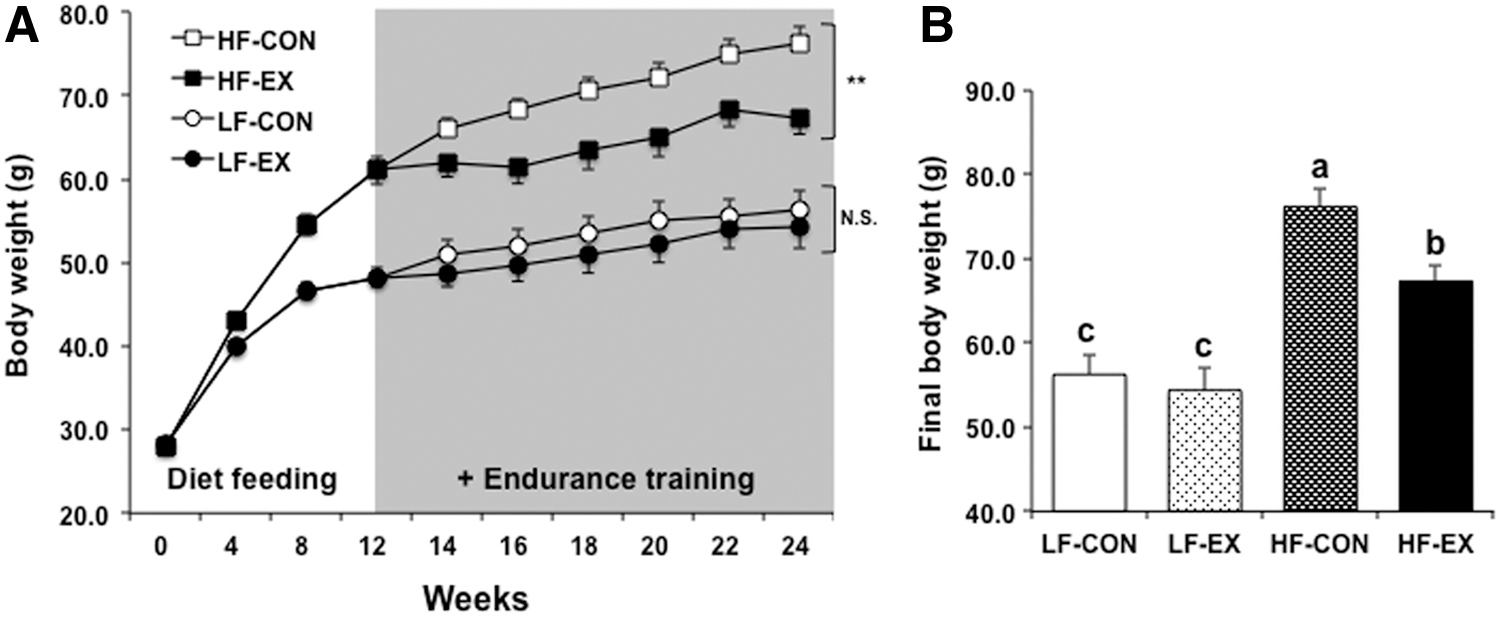
Changes in body weight in the four groups.
Energy metabolism at rest over 24 h
The resting energy expenditure (REE) for 24 h differed significantly between the LFD and HFD groups (diet effect, P < .0001). The exercise-related effect and diet-by-exercise interaction did not differ significantly (P = .284 and .292, respectively). The REE was ∼21% lower in the HF-CON group and 13% lower in the HF-EX group compared to the LF-CON group. The HFD groups reported lower REE than the LFD groups. Notably, the REE was 11% higher in the HF-EX group than the HF-CON group, with the LFD groups demonstrating no difference in exercise between the groups (Fig. 2A, B).
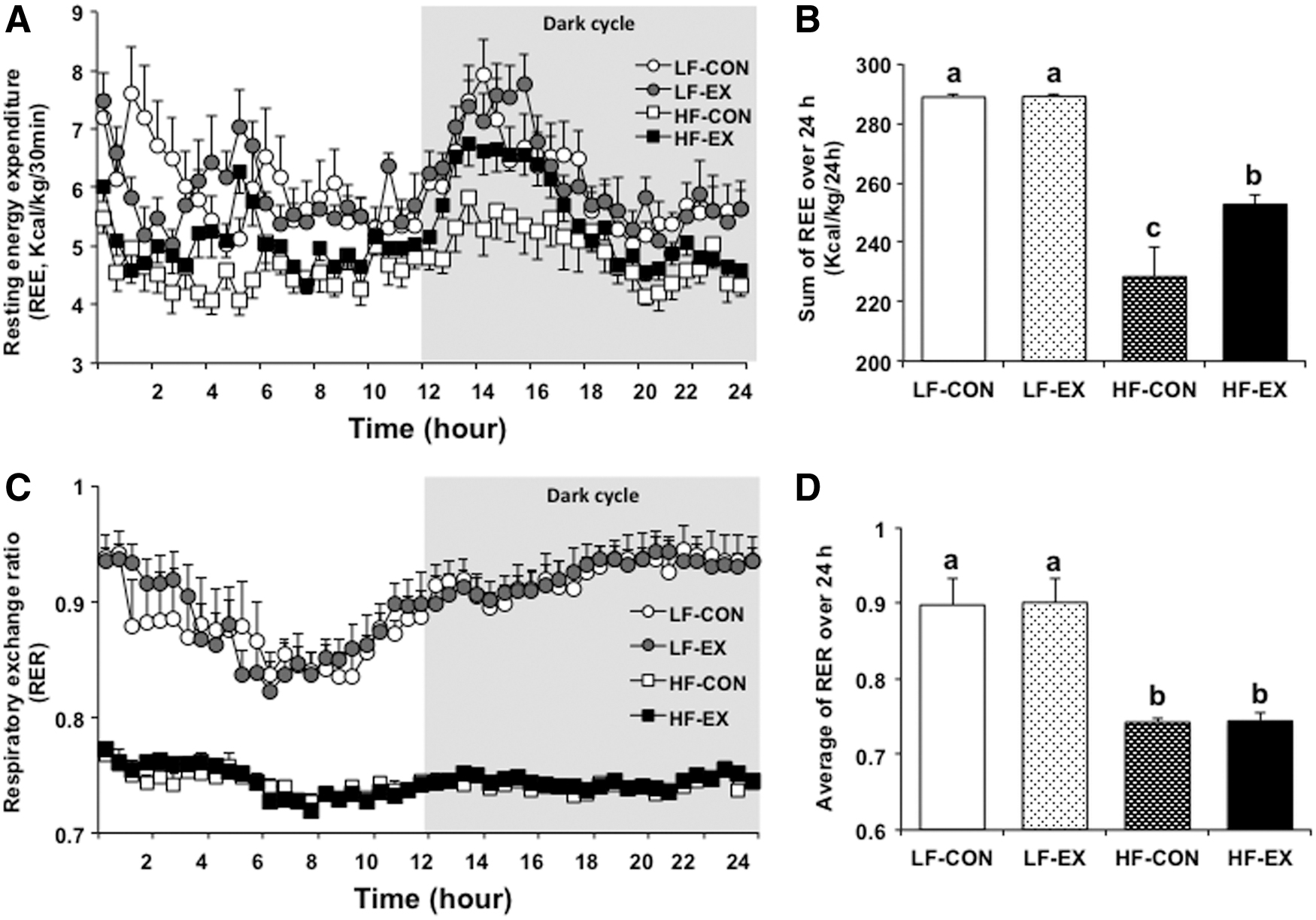
Changes in energy metabolism at rest over 24 h.
The RER, calculated as the ratio of O2 and CO2 produced for 24 h, differed significantly between the LFD and HFD groups (diet effect, P < .0001). The exercise-related effect and diet-by-exercise interaction did not differ significantly (P = .486 and .838, respectively). The LFD groups reported an REE of about 0.9, with the HFD groups reporting an REE of about 0.74 (Fig. 2C, D).
Energy metabolism during exercise for 1 h
RER during running exercise for 1 h differed significantly between the LFD and HFD groups (diet effect, P < .001). In addition, exercise-related and diet-by-exercise effects were observed (P = .0193 and .0017, respectively). LF-CON, LF-EX, HF-CON, and HF-EX groups reported RERs of about 0.86, 0.87, 0.84, and 0.78, respectively. The RER of the HF-EX group significantly differed from all the groups (Fig. 3A, B).

Changes in energy metabolism during exercise for 1 h.
Total fat oxidation during exercise differed significantly between the LFD and HFD groups (diet effect, P = .0012). The exercise-related effect and diet-by-exercise interaction were not significantly different (P = .0971 and .3375, respectively). However, total fat oxidation was 11% higher in the HF-CON group and 46% higher in the HF-EX group, compared to the LF-CON group. Furthermore, fat oxidation was ∼31% higher in the HF-EX group than the HF-CON group. The LFD groups demonstrated no difference in exercise between the groups (Fig. 4A, B).
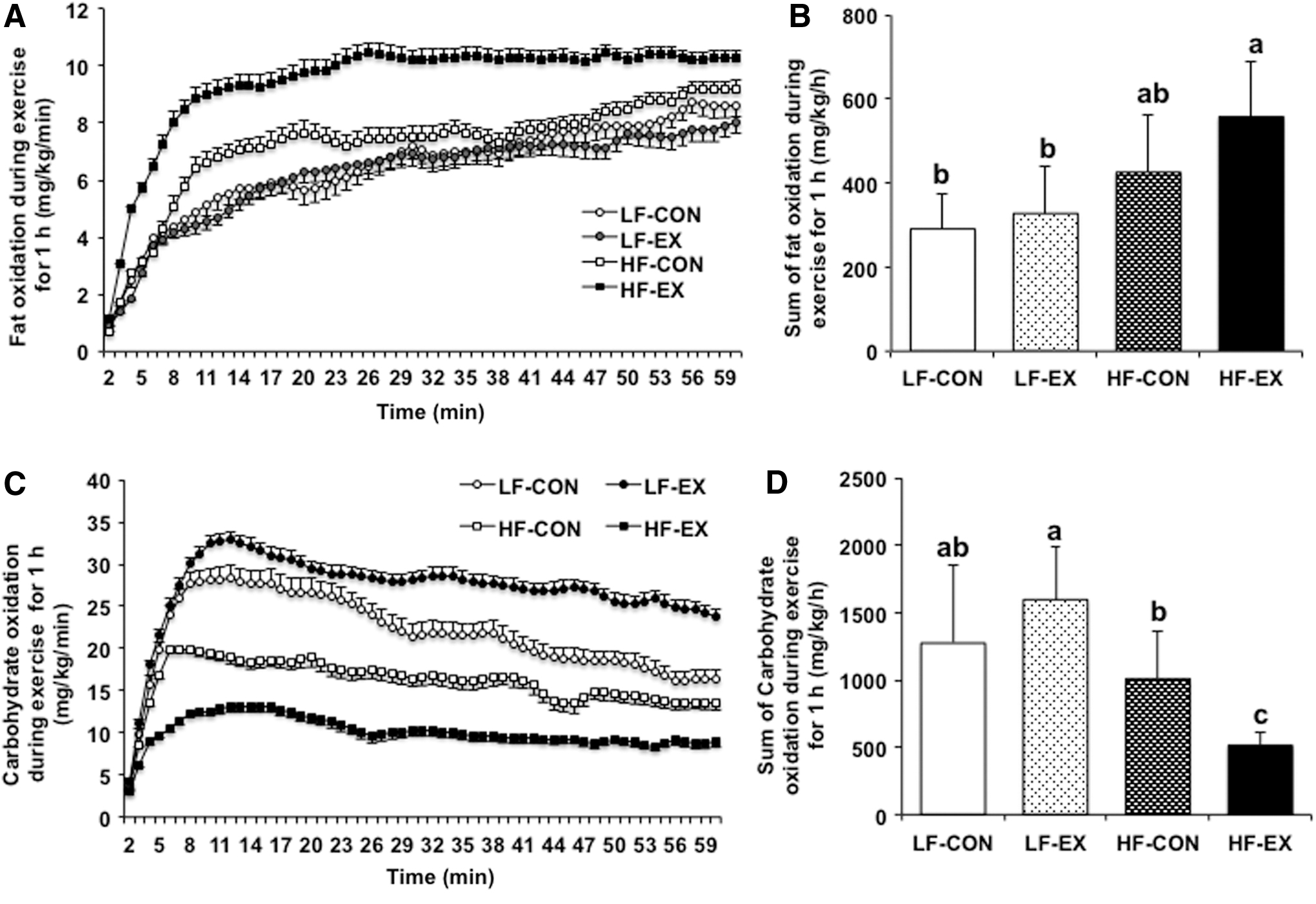
Changes in fat and carbohydrate oxidation during exercise for 1 h.
Total carbohydrate oxidation during exercise differed significantly between the LFD and HFD groups (diet effect, P = .0002). The exercise-related effect was not significantly different (P = .5818), and a diet-by-exercise interaction was observed (P = .0139). The total carbohydrate oxidation was 25% higher in the LF-EX group, 21% lower in the HF-CON group, and 60% lower in the HF-EX group, all compared to the LF-CON group. Carbohydrate oxidation was significantly lower in the HFD groups than in the LFD group; the HF-EX group demonstrated a 49% reduction compared to the HF-CON group (Fig. 4C, D).
Effect of a HFD on the expression of molecules involved in fatty acid transport
The Fat/Cd36 mRNA level differed significantly between the LFD and HFD groups (diet effect, P < .0001). In addition, exercise-related and diet-by-exercise effects were observed (P = .0860 and .0084, respectively). The Fat/Cd36 mRNA level was ∼12% higher in the HF-CON group and 23% higher in the HF-EX group compared to the LF-CON group and did not differ significantly between the LF-EX and the LF-CON groups (Fig. 5A).

The mRNA expression levels of Fat/Cd36
The Fatp1 mRNA level differed between the LFD and HFD groups (diet effect, P = .0245), and the exercise-related effect and diet-by-exercise interaction were not significantly different (P = .6179 and .2437, respectively). The Fatp1 mRNA level was ∼12% greater in the HF-CON group and 27% higher in the HF-EX group compared to the LF-CON group, with no significant difference observed between the LF-EX and the LF-CON groups (Fig. 5B). The Fatp1 mRNA level tended to increase in the HF-EX group compared to the HF-CON group, with no significant difference observed between the groups.
The Fabp-pm mRNA level differed significantly between the LFD and HFD groups (diet effect, P < .0001). The exercise-related effect and diet-by-exercise interaction were not significantly different (P = .5895 and .0897, respectively). The Fabp-pm mRNA level was ∼7% higher in the HF-CON group and 8.8% higher in the HF-EX group compared to the LF-CON group, with no significant difference observed between the two HFD groups. Moreover, the two LFD groups did not differ significantly (Fig. 5C).
The mRNA expression of Cpt1 differed significantly between the LFD and HFD groups (diet effect, P = .0001), an exercise-related effect was not significant (P = .1073), and a diet-by-exercise interaction was observed (P = .0247). The Cpt1 mRNA level was ∼9% greater in the HF-CON group and 17% higher in the HF-EX group compared to the LF-CON group; no significant difference was observed between the LF-EX and LF-CON groups (Fig. 5D).
Discussion
During endurance training, an improvement in exercise capacity can be mediated through changes in metabolism such as the increased utilization of fat and the conservation of glycogen. 23 In this study, we reported that the HFD intake effectively increased fat utilization as an energy substrate during long-term endurance exercise, with LFD intake demonstrating no efficacy, as assessed through the measurement of whole-body energy metabolism. Notably, fat oxidation was optimum when HFD and exercise were combined. Furthermore, we observed changes in the expression of genes involved in fatty acid transport in the skeletal muscle.
In a previous study, Arkinstall et al. reported that Fat/Cd36 mRNA levels in the human skeletal muscle decreased after consuming a high-carbohydrate diet (10 g/kg body mass CHO) for 48 h, compared to a low-carbohydrate diet, after performing an exhaustive exercise bout to deplete the stored muscle glycogen. 17 Cheng et al. reported the effect of dietary glycemic index (GI) on the gene expression of Glut4 and Fat/Cd36 in the human skeletal muscle after a single bout of exercise. After the completion of a 60-min cycling exercise (75% VO2 max), subjects were immediately fed either high-GI or low-GI food. 18 Fat/Cd36 mRNA levels decreased in the high-GI group compared to the low-GI group when tested 3 h after exercise. 18 Most previous studies on the transient effects of postexercise diet have been conducted using short-term exercise. During exercise following a high-fat/low-carbohydrate dietary intake, elevated blood concentrations of nonesterified fatty acids and ammonia may result in the early development of central fatigue. 24 Therefore, an adequate adaptation to a high-fat/low-carbohydrate diet is essential to induce metabolic changes and restore the muscle glycogen. 24 In our study, the mice were adapted to the change in diet for 12 weeks, followed by a 12-week endurance exercise program, while maintaining the same diet for a total of 24 weeks.
After 24 weeks, the substrate utilization in energy metabolism indicated that carbohydrate utilization was high in LFD intake and fat utilization was high in HFD intake, as assessed by the respiratory gas analysis of whole body at rest for 24 h. However, the total energy expenditure was lower in the HFD groups than in the LFD groups and increased markedly with exercise. In contrast, exercise demonstrated no effect when combined with LFD.
Substrate utilization in energy metabolism during exercise for 1 h indicated that carbohydrate utilization was high during LFD intake, while fat utilization was high with HFD intake. Notably, when HFD intake and exercise were combined, carbohydrate utilization was decreased and fat utilization was increased. These energy metabolism results in the whole body were consistent with the same trends in fatty acid transport associated gene expressions in skeletal muscle.
The mRNA expression levels of Fat/Cd36, Fatp1, Fabp-pm, and Cpt1 were higher in the HF-CON group than in the LF-CON group. In particular, the Fat/Cd36 and Cpt1 mRNA expressions increased significantly in the HF-EX group compared to that in the HF-CON group, with no significant differences observed between the LF-EX and LF-CON groups. These results indicated that the gene expression levels of all transporters were upregulated by the HFD intake for 24 weeks, and Fat/Cd36 and Cpt1 mRNA were markedly elevated with the addition of long-term endurance exercise. Thus, FAT/CD36 and CPT1 play key roles in fatty acid oxidation during endurance exercise in the skeletal muscle. In contrast, exercise showed no effect when combined with the LFD intake. These data suggest that the HFD may promote the utilization of fat as an energy source and increase the efficiency of fat oxidation during endurance exercise.
Previously, the important role of FAT/CD36 in fatty acid transport was demonstrated using either selective chemical inhibition or muscle-specific gene overexpression. 25,26 Furthermore, it has been reported that muscle contraction increases FAT/CD36 expression. 27 Several studies have also reported that mRNA or/and protein expression of FAT/CD36 and CPT1 was increased by combining short-term endurance exercise with a normal diet. 13,14
Endurance exercise induces an increase in capillary density, expression of substrate transport proteins, and mitochondrial volume density in skeletal muscles, resulting in greater fat oxidation. 13,14,28 –30 It was also reported that after endurance exercise training, CPT1 activity increased at the same rate as that of the mitochondrial content. 14 Endurance exercise may stimulate the upregulation of factors involved in fat metabolism. However, in the exercise group fed the LFD, the mRNA expression levels of Fat/Cd36 and Cpt1 were comparable to those in the nonexercising LFD group. Furthermore, the respiratory gas analysis in whole body did not affect the fat metabolism in the exercise group fed a LFD. These results suggest that a HFD may explain the observed increase in fat oxidation during exercise.
Our data indicate that an increase in dietary fatty acid intake results in upregulation of Fat/Cd36 and that the FAT/CD36 transporter plays a role in facilitating uptake of LCFAs in skeletal muscle. Furthermore, oxidation of LCFAs may be facilitated by an increase in CPT1 during endurance exercise.
Further studies are crucial to investigate the molecular changes in carbohydrate metabolism and assess clinical observations based on these results.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the National Research Foundation of Korea Grant funded by the Korean Government (NRF-2012S1A5B5A01025457) and by the Ministry of Agriculture, Food and Rural Affairs (714001-07), Republic of Korea.