6-Acetyl-2,2-Dimethylchroman-4-One Isolated from Artemisia princeps Suppresses Adipogenic Differentiation of Human Bone Marrow-Derived Mesenchymal Stromal Cells via Activation of AMPK
Available accessResearch articleFirst published online March, 2020
6-Acetyl-2,2-Dimethylchroman-4-One Isolated from Artemisia princeps Suppresses Adipogenic Differentiation of Human Bone Marrow-Derived Mesenchymal Stromal Cells via Activation of AMPK
Obesity is a world-wide health concern with increasing mortality and morbidity rates. Development of novel therapeutic agents for obesity from phytochemicals may lead to the effective prevention and control of obesity and obesity-related complications. 6-acetyl-2,2-dimethylchroman-4-one (1) was isolated from a dietary plant, Artemisia princeps. The antiobesity effect of compound 1 was determined in human bone marrow-derived mesenchymal stromal cells (hBM-MSCs) induced to differentiate into adipocytes. Treatment with compound 1 resulted in decreased lipid accumulation and expression of key adipogenic markers, proliferator-activated receptor-γ, CCAAT/enhancer-binding protein-α, and sterol regulatory element-binding transcription factor 1. It was also shown that compound 1 downregulated the adipogenesis-induced p38 and JNK MAPK activation, while upregulating adipogenesis inhibitory β-catenin-dependent Wnt10b pathway. Compound 1 was also able to stimulate adenosine monophosphate-activated protein kinase phosphorylation, which was suggested to be the underlying mechanism that resulted in inhibition of adipogenesis in hBM-MSCs. In conclusion, 6-acetyl-2,2-dimethylchroman-4-one was identified as a bioactive constituent of A. princeps that exerts antiobesity properties via suppressing adipocyte formation.
Introduction
Obesity is a worldwide health problem that threatens increasing numbers of people of all ages. Besides causing mortality and being the reason for several complications that contribute to lower life quality, obesity is linked with various life-threatening diseases such as type-2 diabetes, cardiovascular diseases, osteoporosis, and cancer.1,2 Obesity, which is characterized by deteriorated adipose tissue function and increased fat accumulation, has been the subject of several preventive medicinal studies. Controlling adipocyte formation is regarded as a potential way to hinder obesity progression.3 In addition to adipocyte differentiation in adipose tissue itself, obesity-linked complications are known to meditate the other adipogenesis-related disorders, including osteoporosis, informally labeled as obesity of the bone.4 Activated adipocyte differentiation mechanisms whether due to obesity or not, results in elevated number of adipocytes in bone marrow compared to osteoblasts and osteocytes which are responsible for bone formation.5 Studies have also shown that in addition to adipose tissue-derived MSCs, MSC from other tissues such as bone marrow can also play a role in obesity-related adipocyte differentiation in adipose tissue.5,6 Human bone marrow-derived mesenchymal stromal cells (hBM-MSCs) have the ability to differentiate into different cell types, including adipocytes and osteoblasts. Cultured hBM-MSCs with differentiation inducement are therefore an important in vitro model for screening any mechanism to prevent adipocyte differentiation.7
Phytochemicals are one of the possible ways to regulate the energy metabolism and lower the risks of developing obesity-related complications.8–10 To date, studies have reported several plants with substances that can inhibit adipogenesis with different action mechanisms. In this context, Artemisia species have produced different results in terms of their effects on adipogenesis. While A. scoparia and A. santolinifolia were reported to promote adipogenesis,11Artemisia princeps extracts inhibited the adipocyte differentiation.12A. princeps is a common edible plant native to Korea, Japan, and China, also known as mugwort or wormwood. It has been a part of daily diet as well as traditional medicine practices as it exerts beneficial effects against gastric and bleeding problems.13 Studies also reported antidiabetic,14 antimicrobial,15 antihistaminic,16 and antioxidative13 effects for A. princeps extract and constituents along their adipogenesis inhibitory properties.
As a part of ongoing research to develop antiobesity natural compounds, A. princeps was studied and 6-acetyl-2,2-dimethylchroman-4-one was isolated as one of its active phytochemicals. The current study aimed to report the effects of 6-acetyl-2,2-dimethylchroman-4-one on the adipogenic differentiation of hBM-MSCs.
Materials and Methods
Plant material, extraction, and isolation
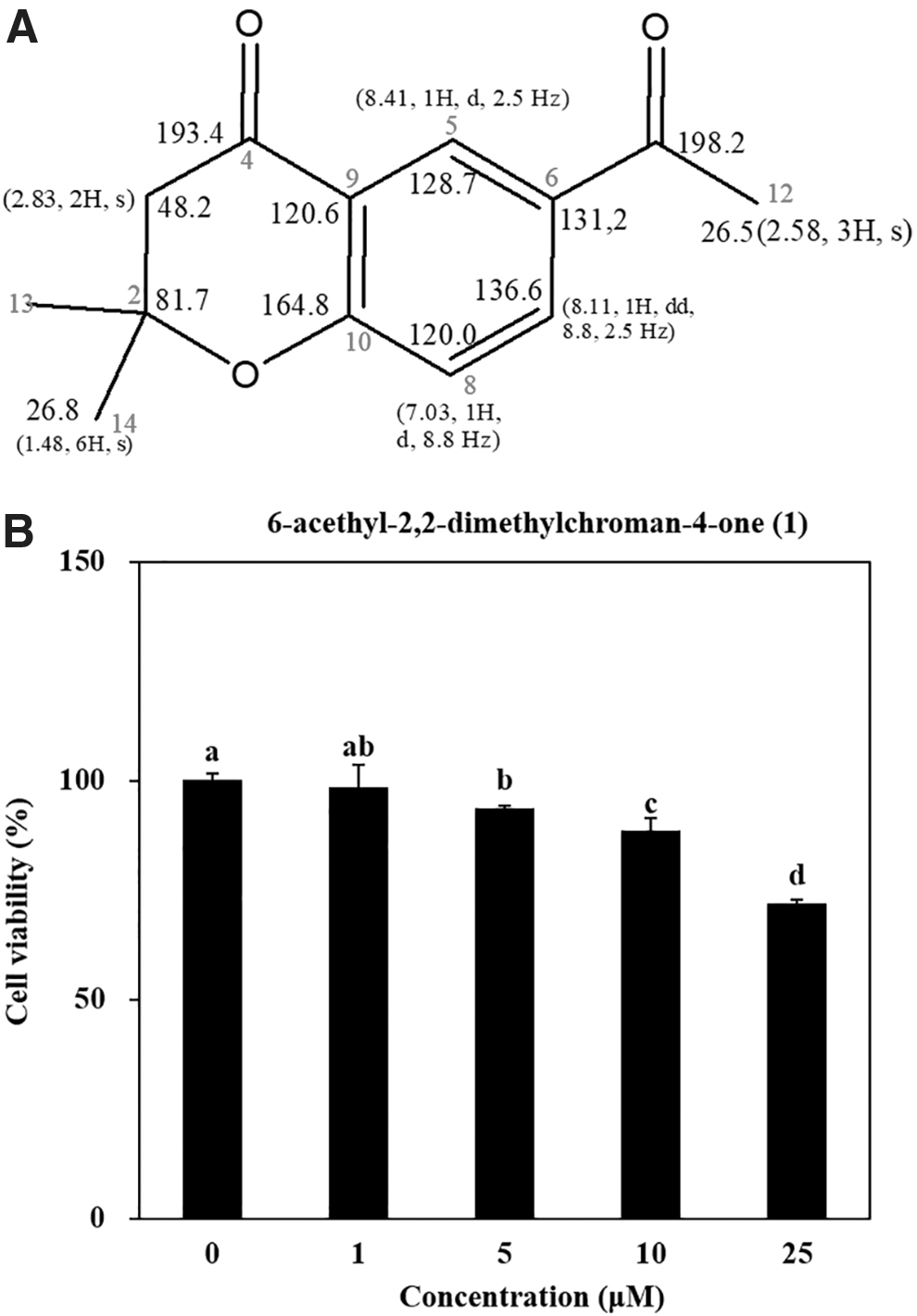
6-acetyl-2,2-dimethylchroman-4-one (1) was isolated from 85% aq. MeOH solvent fraction of A. princeps extract was prepared as described earlier.12 Chemical structure and characterization of the compound 1 were carried out by1H and13C NMR, and comparison of spectral data with published literature was also carried out (Fig. 1A).17,18
Chemical structure of 6-acethyl-2,2-dimethylchroman-4-one (1) (A) with 1H and 13C NMR readings for representative numbering and cytotoxicity of compound 1 in hBM-MSCs (B). Cells were seeded (3 × 103) in 96-well plates and incubated for 48 h untreated or treated with different concentrations (1, 5, 10, 25 μM) of compound 1. Viability of the cells was measured by quantification of MTT dye removed from cells. Cell viability was given as the percentage of untreated control group (0 μM). Different letters (a–d) indicate statistically significant difference (P < .05) by Duncan's multiple range test. hBM-MSCs, human bone marrow-derived mesenchymal stromal cells; MTT, 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide.
Cell culture and viability assay
hBM-MSCs obtained from PromoCell (hBM-MSC, C-12974) were cultured in 6-well plates unless otherwise noted. hBM-MSCs were fed with mesenchymal stromal cell growth medium (C-28009, PromoCell) and incubated at 37°C with 5% CO2. For adipogenic differentiation, following 100% confluence, hBM-MSCs were adipo-induced by MSC adipogenic differentiation medium 2 (C-28016, PromoCell) and incubated for 14 days unless otherwise noted for the specific analysis. Compound 1 was introduced with initial differentiation medium change and included in consequent fresh media (every third day). The same procedure for adipogenic differentiation of hBM-MSC was followed, except for replacing differentiation-inducing medium with MSC.
Effects of compound 1 on the viability of hBM-MSCs was evaluated by 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay as previously described.12 Cell culture medium was aspired after 48 h incubation with different concentrations of compound 1, and 100 μL of MTT reagent (1 mg/mL) was added to the wells before 4 h incubation. Formazan salt was dissolved with the addition of 100 μL DMSO to the wells. Viability and proliferation of the cells were calculated by measuring the absorbance values of the formed formazan salts for each well at 540 nm using a Multiskan GO microplate reader (Tecan Austria GmbH, Grodig, Austria).
Oil Red O staining
Staining of intracellular lipid droplets was carried out with common Oil Red O staining protocols. In brief, adipo-induced hBM-MSCs were fixed on six-well plates at day 14 of incubation via addition of 10% fresh formaldehyde (in PBS, v/v) to the wells for 1 h at room temperature. One milliliter of 5% Oil Red O solution (m/v, 60% isopropanol and 40% water) was then introduced to the wells after the fixing solution was removed and cells were washed once with PBS. After 1 h of incubation, staining solution was removed from wells before being dried. Images of lipid droplets in adipo-induced hBM-MSCs were taken using an optical microscope (Olympus, Tokyo, Japan). For the quantification of the dye bound to lipid droplets, Oil Red O was eluted from cells with the addition of 100% isopropyl alcohol. The amount of dye was calculated by measurement of the absorbance at 500 nm with a microplate reader (Multiskan GO, Tecan Austria GmbH, Austria).
Total RNA (2 μg) was obtained from adipo-induced hBM-MSCs using TRIzol reagent (Invitrogen, CA, USA) on day 7 of differentiation. Next, cDNA was synthesized according to the common two-step reverse transcription-polymerase chain reaction (RT-PCR) protocols. RNase-free water containing oligo (dT), random primers, and total RNA was first subjected to denaturation at 70°C for 5 min. Reverse transcription occurred in a master mix containing 1 × RT buffer, 1 mM dNTPs, 500 ng oligo (dT), 140 U M-MLV reserve transcriptase, and 40 U RNase inhibitor with cycles of 42°C for 60 min and 72°C for 5 min using an automatic T100 Thermal Cycler (Bio-Rad, UK). Previously described12 gene-specific sense and antisense primers were used for the amplification of the target cDNA via ready to use commercial master mix (Bio-Rad). The master mix containing synthesized cDNA was subjected to 30 cycles of 95°C for 45 sec, 60°C for 1 min, and 72°C for 45 sec using T100 Thermal Cycler (Bio-Rad). Amplified target products were separated by gel electrophoresis on 1.5% agarose gel for 30 min at 100 V. Bands were then observed following the staining with 1 mg/mL ethidium bromide under UV light using CAS-400SM Davinch-Chemi imager™ (Seoul, Korea).
Western blotting
Protein immunoblotting was performed with standard western blotting procedures. At day 7 of differentiation, adipo-induced hBM-MSCs were harvested by pipetting with 1 mL of RIPA lysis buffer (Sigma-Aldrich) at 4°C for 30 min and the lysate (25 μg) was used for western blot analysis. Nuclear fractions were obtained by using a commercial nuclear extraction kit (NE-PER™ Nuclear Extraction Kit, Catalog No. #78835; Thermo Fisher Scientific, Waltham, MA, USA) according to manufacturer's instructions. hBM-MSCs were treated with adenosine monophosphate-activated protein kinase (AMPK) inhibitor compound C at the beginning of differentiation with or without compound 1 and the harvesting was performed before differentiation and at 6, 12, and 24 h of differentiation for western blotting. Separation of proteins was achieved by sodium dodecyl sulfate-polyacrylamide gel electrophoresis on 4% stacking and 12% separating gels. Proteins separated on gels were electrotransferred to a polyvinylidene fluoride membrane (Amersham Pharmacia Biosciences, UK). Membrane was blocked with 5% skim milk in TBST buffer (v/v) for 4 h. Blocked membrane was hybridized at 4°C overnight with primary antibodies diluted (1:1000) with 1 × TBST buffer containing 5% bovine serum albumin (v/v). Hybridized membrane was incubated with horseradish-peroxidase-conjugated secondary antibody at room temperature for 2 h. Immunoreactive protein bands were visualized by a commercial chemiluminescence assay kit based on luminol (Amersham Pharmacia Biosciences) following the manufacturer's instructions. Images of membranes with protein bands were obtained by CAS-400SM Davinch-Chemi imager™ (Seoul, Korea).
Statistical analysis
The data were presented as mean of three independent experiments ± SD run in triplicates unless otherwise noted. Statistically significant differences among the means of the individual test groups were assessed by one-way analysis of variance (ANOVA) with post hoc Duncan's multiple range test using SAS v9.1 software (SAS Institute, Cary, NC, USA), P < .05 being the defining level for the significance of differences.
Results
Before the evaluation of compound 1 for antiadipogenic effects, its cytotoxicity was assessed in hBM-MSCs via MTT assay. As seen in Figure 1B, doses higher than 10 μM decreased the viability of the cells to below 90% and were therefore deemed too toxic to be used in subsequent assays. The safe doses were determined to be 1, 5, and 10 μM of compound 1.
Oil Red O staining
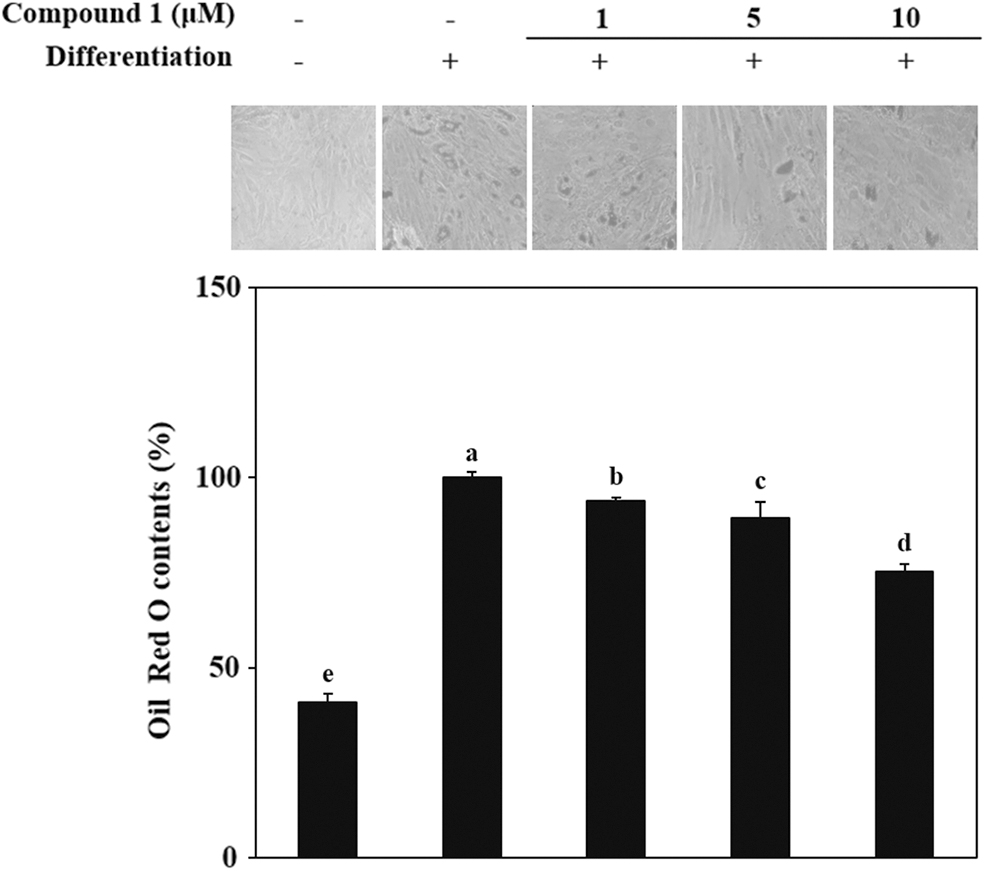
The level of adipogenesis in adipo-induced hBM-MSCs was determined at day 14 of differentiation via accumulation of lipid droplets in the cytoplasm. Most of the adipo-induced cells without compound 1 treatment accumulated high amounts of lipids, as revealed by staining with Oil Red O (Fig. 2). Compared to the untreated adipo-induced control group, cells treated with compound 1 accumulated significantly less lipid. It is also shown that the inhibition of lipid accumulation by compound 1 was in a concentration-dependent manner. Quantification of the Oil Red O stain from cells by measurement of optical density further confirmed the results. At 5 and 10 μM concentrations, compound 1 treatment decreased the lipid accumulation of adipo-induced hBM-MSCs by 19% and 25%, respectively, compared to control.
Effect of compound 1 on the lipid accumulation of adipo-induced hBM-MSCs. Cells were seeded in six-well plates and induced to differentiate with adipocyte differentiation medium in the absence or presence of compound 1 (1, 5, and 10 μM). Following 14 days of incubation, intracellular lipid droplets of mature adipocytes were stained with Oil Red O. Images of cells show stained lipid droplets (upper panel). Lipid accumulation levels were calculated by the colorimetric quantification of the dye removed from the wells (lower panel) and given as percentage of adipo-induced untreated control group. Different letters (a–e) indicate statistically significant difference (P < .05) by Duncan's multiple range test.
The mRNA and protein expression levels of key adipogenesis markers
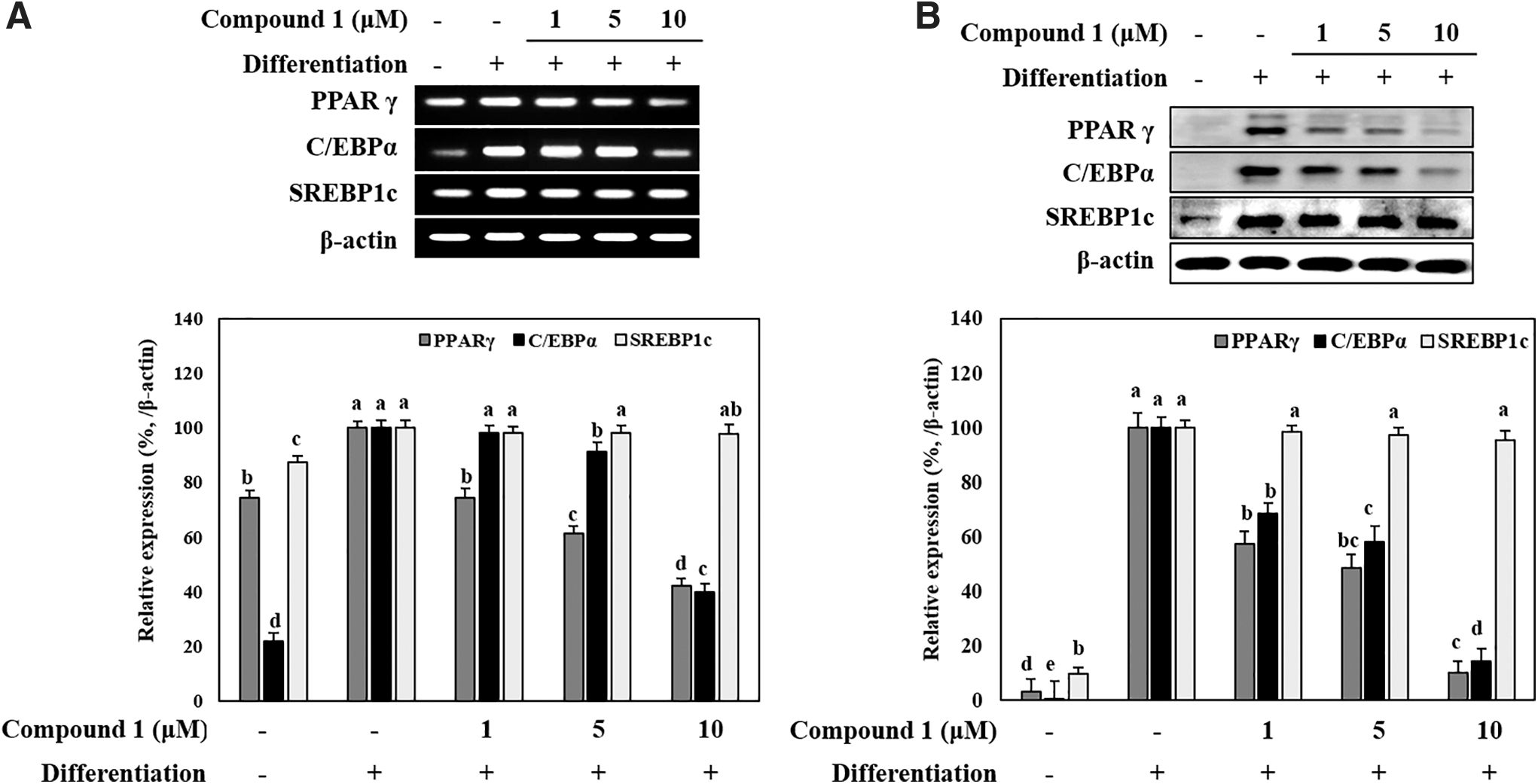
The mRNA and protein expressions of the adipogenic markers proliferator-activated receptor-γ (PPARγ), CCAAT/enhancer-binding protein-α (C/EBPα) and sterol regulatory element-binding transcription factor 1 (SREBP1c) were observed using RT-PCR and immunoblotting, respectively. Adipo-induced hBM-MSCs expressed higher amounts of adipogenic markers as expected (Fig. 3A). At day 7 of differentiation, hBM-MSCs treated with compound 1 expressed less adipogenic marker mRNA decreasing in a concentration-dependent manner. Consistent with these results, protein expression of adipogenic markers was also decreased by compound 1 treatment. Western blotting showed that the protein levels of PPARγ, C/EBPα, and SREBP1c were notably increased by adipogenesis inducement and dose-dependently decreased by compound 1 for PPARγ and C/EBPα (Fig. 3B). Although a slight decrease in SREBP1c was observed by compound 1 treatment, it was not significant, indicating that compound 1 suppressed adipogenesis via a relatively SREBP1c-independent mechanism.
Effect of compound 1 on the mRNA (A) and protein (B) expression levels of key adipogenic differentiation markers, PPARγ, C/EBPα, and SREBP1c in adipo-induced hBM-MSCs. Cells were seeded in six-well plates and induced with adipocyte differentiation medium in the absence or presence of compound 1 (1, 5 and 10 μM). Following 14 days of incubation, cell lysates were used for the detection of PPARγ, C/EBPα, and SREBP1c. β-actin was used as internal loading control. Expression levels were densiometrically quantified and normalized against the levels of housekeeping control, β-actin. Effect of compound 1 on expression levels was given as the relative percentage of the adipo-induced untreated control cells. Different letters (a–e) indicate statistically significant difference (P < .05) by Duncan's multiple range test. C/EBPα, CCAAT/enhancer-binding protein-α; PPARγ, proliferator-activated receptor-γ; SREBP1c, sterol regulatory element-binding transcription factor 1.
The protein expression and activation of MAPK, Wnt, and AMPK pathways
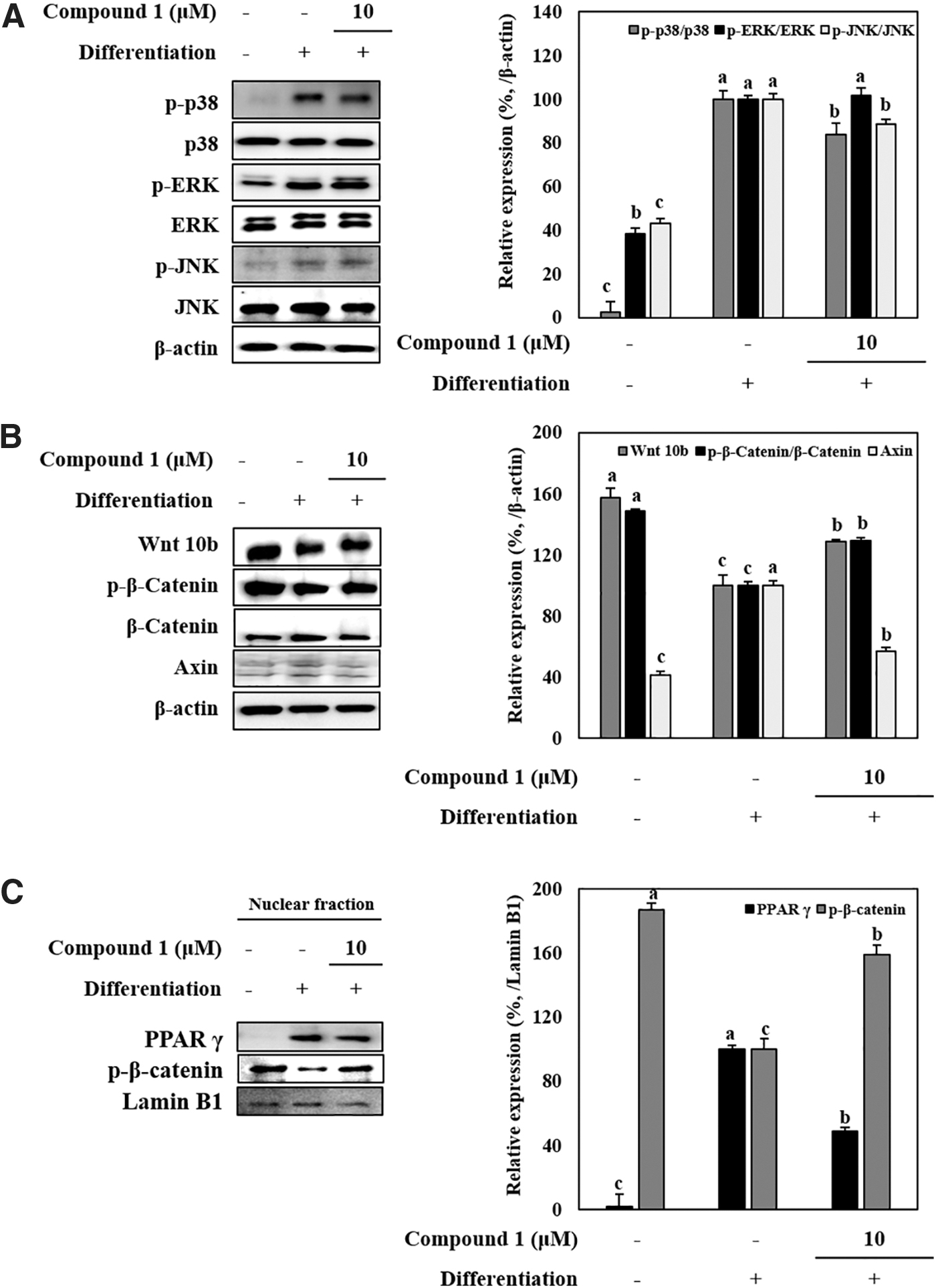
The effects of compound 1 on the activation of MAPK pathway were detected by comparing the protein levels of phosphorylated (p-) and total forms of p38, ERK, and JNK MAPK proteins. Adipo-induced hBM-MSCs showed increased activation for MAPK signaling shown by increased p-p38, p-ERK, and p-JNK levels, whereas total protein levels for all three MAPKs stayed at the same level (Fig. 4A). Presence of compound 1 (10 μM) resulted in a significant decrease for phosphorylation of p38 as well as slight decrease in p-JNK levels. However, compound 1 did not exert any significant decrease in ERK phosphorylation. In addition to MAPK levels, Wnt/β-catenin pathway was also observed under compound 1 treatment. Adipogenesis inhibitory Wnt pathway markers Wnt10b, β-catenin, and Axin levels were detected by western blotting. Inducement of hBM-MSCs toward adipogenesis caused a decrease in the Wnt10b protein levels as well as phosphorylation of β-catenin (Fig. 4B). Compound 1 (10 μM) alleviated the adipogenesis-linked decrease in Wnt10b and p-β-catenin levels. Axin levels were increased by adipogenesis, but they were also was decreased by compound 1 treatment. The adipogenic transcription factor PPARγ, and Wnt pathway transcription factor β-catenin were investigated in nuclear fractions of adipo-induced hBM-MSCs by western blotting to provide insights into their transcriptional activities. Parallel to previous results, adipocyte differentiation resulted in increased translocation of PPARγ to the nucleus as opposed to decreased levels of β-catenin (Fig. 4C). Compound 1 (10 μM) treatment strongly increased the β-catenin levels in nuclei while suppressing the PPARγ.
Effect of compound 1 on the protein levels of MAPK and Wnt pathway proteins in adipo-induced hBM-MSCs. Cells were seeded in six-well plates and induced with adipocyte differentiation medium in the absence or presence of compound 1 (10 μM). Following 14 days of incubation, cell lysates were used for the detection of total and phosphorylated (p-) protein levels of p38, ERK, and JNK for MAPK pathway analysis (A), Wnt10b, β-catenin and Axin for Wnt pathway analysis (B). β-actin was used as internal loading control. Nuclear fractions of adipo-induced hBM-MSCs at day 14 incubation were used for the detection of PPARγ and β-catenin transcription factors (C). Lamin B1 was used as nuclear loading control. Expression levels were densiometrically quantified and normalized against the levels of housekeeping control, β-actin. Expression levels of p38, ERK, JNK, and β-catenin (A, B) were given as their phosphorylation rate relative to their nonphosphorylated levels. Effect of compound 1 on expression levels was given as the relative percentage of the adipo-induced untreated control cells. Different letters (a–c) indicate statistically significant difference (P < .05) by Duncan's multiple range test.
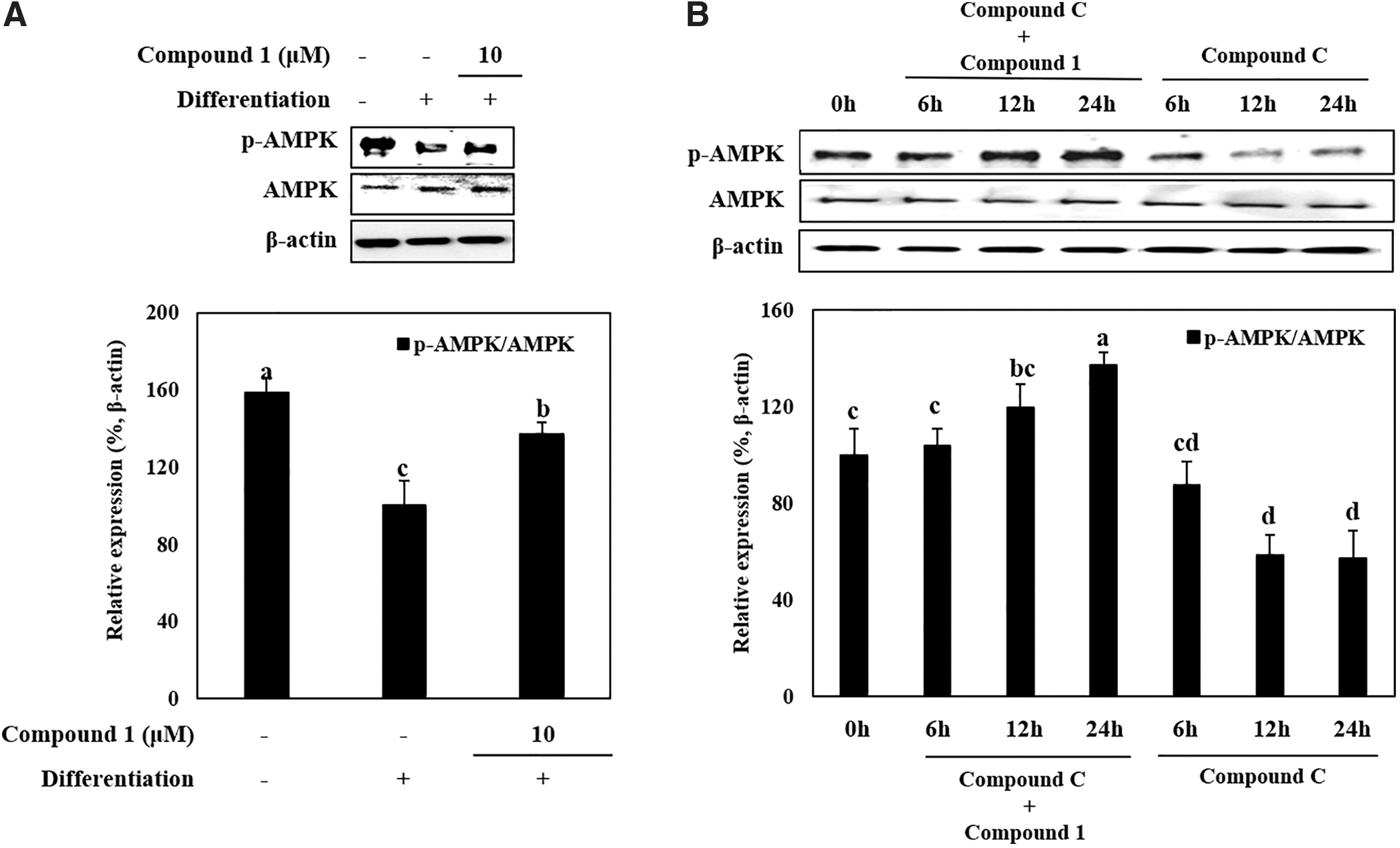
AMPK phosphorylation in adipo-induced hBM-MSCs were downregulated by adipo-inducement shown by decreased p-AMPK protein levels compared to increased total AMPK levels (Fig. 5A). The protein level of p-AMPK was notably increased by 10 μM compound 1. The stimulatory effect of compound 1 on AMPK activation was further shown using a specific AMPK phosphorylation inhibitor, compound C. Presence of compound C time dependently decreased the p-AMPK levels in differentiating adipo-induced hBM-MSCs (Fig. 5B). Treating cells with compound 1 along with compound C exerted a strong time-dependent increase in the levels of active AMPK indicating that compound 1 intervened in the AMPK phosphorylation pathway. Gradually increasing AMPK phosphorylation despite treatment with compound C indicated that the enhancement effect of compound 1 on AMPK activation was independent of compound C-mediated AMPK inhibition.
Effect of compound 1 on the total and phosphorylated (p-) levels of AMPK in adipo-induced hBM-MSCs; at day 14 of incubation (A) and after treatment with AMPK activation inhibitor compound C at 6, 12, and 24 intervals (B). Cells were seeded in six-well plates and induced with adipocyte differentiation medium in the absence or presence of compound 1 (10 μM). Following 14 days of incubation, cell lysates were used for the detection using western blotting. β-actin was used as internal loading control. Expression levels were densiometrically quantified and normalized against the levels of housekeeping control, β-actin. Expression levels of AMPK (A, B) were given as its phosphorylation rate relative to their nonphosphorylated levels. Effect of compound 1 on expression levels was given as the relative percentage of the adipo-induced untreated control cells. Different letters (a–d) indicate statistically significant difference (P < .05) by Duncan's multiple range test. AMPK, adenosine monophosphate-activated protein kinase.
Discussion
Progression of obesity is closely linked with the formation of new adipocytes via adipogenic differentiation. Mesenchymal stromal cells are the main sources of adipocytes in the human body, either in adipose tissue or bone.19 Obesity-linked increases in adipogenic differentiation affects whole body and can be counted among causes of several other diseases such as osteoporosis.1 Obesity-mediated changes in differential tendencies of bone marrow MSCs are the main cause of the lack of bone formation as the MSCs predominantly differentiate into adipocytes rather than osteoblasts.20 Therefore, the current study used hBM-MSCs as the in vitro model for determining the effects of compound 1 on adipogenesis dysregulation.
Previous studies showed that A. princeps extract exerted antiadipogenesis effects in 3T3-L1 murine preadipocyte cell line.12,13 It was reported that A. princeps inhibited the adipocyte differentiation via antioxidative activity and MAPK pathway suppression. These studies have supported the current results although they were conducted in immortalized mouse preadipocytes. Use of hBM-MSCs in the current study provided additional evidence that A. princeps had potential antiobesity effects.
On the contrary, compound 1 was previously reported only for its cytotoxic activity in cancer cells21 and antiplatelet aggregation activity,22 being isolated from Eupatorium betonicaeforme and Gynura japonica, respectively. To the best of our knowledge, this is the first report of compound 1 isolation from A. princeps and the first report of its antiadipogenesis effect. It was shown that compound 1 significantly suppressed the adipogenic differentiation of hBM-MSCs by inhibiting the lipid accumulation and the expression of adipogenesis markers (PPARγ and C/EBPα). PPARγ activation is a well-recognized pathway for adipogenic stimulation of the MSCs and it can subsequently hinder the osteogenic differentiation, which is the reason behind the lack of bone formation and excessive lipid accumulation.23 C/EPBα and SREBP1c are downstream genes of PPARγ signaling and have important roles in lipid metabolism.24 Compound 1 inhibited transcriptional and translational expression of C/EPBα suggesting an intracellular intervention for hindering adipogenic differentiation of MSCs.
To provide insights on how compound 1 suppressed the PPARγ-mediated adipogenesis, its effects on MAPK and Wnt10b pathways were determined. MAPK is a serine/threonine protein kinase and can be found in numerous cell types. It has three main subunits, namely p38, ERK, and JNK. Activation of MAPK pathway has been linked with several cellular processes such as energy metabolism, apoptosis, differentiation, and proliferation.25 Reports have shown that MAPK activation promotes or inhibits adipogenic differentiation depending on the stimulatory signal.26 Current results showed that adipogenesis inducement resulted in overactivation of MAPK in hBM-MSCs. Presence of compound 1 was able to decrease the activation of MAPKs, mainly p38 phosphorylation, while suppressing adipogenesis. This suggested that compound 1 treatment reverted the adipogenic stimulatory signal on MAPK pathway. Previous studies confirmed that the inhibition of p38 activation suggestively blocks adipogenesis,27 which could be considered parallel to the results of this study.
In addition, Wnt/β-catenin pathway is also an important regulator of MSC differentiation.28 Activation or suppression of Wnt pathways evidently decides the fate of MSC differentiation. Among different Wnt subtypes, Wnt10b is a well-recognized mediator of MSC osteogenesis.29 Activation of Wnt10b downstream signaling inhibits adipogenesis while promoting osteoblast differentiation of MSCs through a β-catenin-dependent mechanism.30 This was also shown in the present results, as the Wnt10b and activated β-catenin levels were suppressed in adipo-induced hBM-MSCs. However, compound 1 treatment slightly increased the diminished Wnt10b and β-catenin protein levels. Axin is an inhibitory regulator for osteoblast differentiation and is subsequently expressed in differentiating MSCs during adipogenesis.31 Increased Axin levels were decreased by compound 1 treatment as well, further suggesting that compound 1 hinders adipogenesis of hBM-MSCs while consequently relieving suppressed osteogenic pathways. Effect of compound 1 on transcriptional activities of both adipogenesis and osteoblastogenic transcription factors showed consistent results. Protein levels of PPARγ and β-catenin in nuclear fractions of the adipo-induced hBM-MSCs were reversed after compound 1 treatment.
Reports have shown that AMPK activation mediates the initial differentiation of MSCs.32,33 Activation of AMPK results in enhanced osteoblast differentiation, whereas suppression results in enhanced adipogenesis. The present study showed that levels of phosphorylated AMPK in adipo-induced hBM-MSCs were significantly lower compared to the noninduced group. At 10 μM, compound 1 notably increased the phosphorylated AMPK levels. This effect of compound 1 was further confirmed using compound C, an inhibitor for AMPK activation.34 Adding compound C during the adipogenesis inducement strongly inhibited the levels of activated AMPK. Introduction of compound 1 to the adipo-induced hBM-MSCs along with compound C removed the inhibition of AMPK activation in a time-dependent manner. After 24 h of treatment, levels of activated AMPK were found to be higher than the noninduced group, despite the presence of compound C. Hence, it was suggested that compound 1 was able to both suppress PPARγ-mediated adipogenesis and enhance adipogenesis inhibitory pathways such as Wnt10b/β-catenin through the stimulation of AMPK phosphorylation.
Compound 1 is derived from the chroman-4-one scaffold, which is an important and promising base for both organic and synthetic drug development. Compound 1 has a chemical framework where both the 2,3-dihydro-γ-pyranone ring and benzene nucleus of chroman-4-one have active side chains. This scaffold is very similar to chromone, but differs from it by the absence of C2-C3 double bond in the 2,3-dihydro-γ-pyranone ring. It was suggested that this difference results in differences in its biochemistry as well as bioactivity of the compounds possessing a chroman-4-one frame.35 Several compounds were discovered or synthesized with the chroman-4-one framework, including flavanones and hydrazones. Among the numerous bioactive chroman-4-one derivatives, several of them were regarded as promising pharmacophores for development of novel drug candidates against diabetes, obesity, and linked metabolic syndromes.36 In this context, the current results showing the antiadipogenic potential of compound 1 provides valuable insights for further studies to develop antiobesity drug candidates based on compound 1 chemical framework. In addition, this was the first instance of compound 1 being isolated from A. princeps although other chroman-4-one derivatives were reported from different Artemisia species.37
In conclusion, this study reported for the first time the antiadipogenic effect of compound 1, which was isolated from A. princeps. It was revealed that compound 1 inhibited the adipogenic differentiation of hBM-MSCs via AMPK activation. The effects of compound 1 were suggested to be an important reference to further studies for phytochemical-mediated prevention and treatment of obesity-related complications.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the National Research Foundation of Korea (NRF) grant funded by the Korea government (MSIP) (No. NRF-2017R1A2B4009588).
References
1.
BlüherM: Adipose tissue dysfunction contributes to obesity related metabolic diseases. Best Pract Res Clin Endocrinol Metab, 2013; 27:163–177.
2.
CohenDH, LeRoithD: Obesity, type 2 diabetes, and cancer: The insulin and IGF connection. Endocr Relat Cancer, 2012; 19:27–45.
3.
ZaiouM, El AmriH, BakillahA: The clinical potential of adipogenesis and obesity-related microRNAs. Nutr Metab Cardiovasc Dis, 2018; 28:91–111.
4.
RoyB, CurtisME, FearsLS, NahashonSN, FentressHM: Molecular mechanisms of obesity-induced osteoporosis and muscle atrophy. Front Physiol, 2016; 7:439.
5.
AmbrosiTH, ScialdoneA, GrajaA, et al.: Adipocyte accumulation in the bone marrow during obesity and aging impairs stem cell-based hematopoietic and bone regeneration. Cell Stem Cell, 2017; 20:771–784.
6.
da SilvaSV, Renovato-MartinsM, Ribeiro-PereiraC, CitelliM, Barja-FidalgoC: Obesity modifies bone marrow microenvironment and directs bone marrow mesenchymal cells to adipogenesis. Obesity (Silver Spring), 2016; 24:2522–2532.
7.
MatsushitaK, DzauVJ: Mesenchymal stem cells in obesity: Insights for translational applications. Lab Investig, 2017; 97:1158–1166.
8.
FuC, JiangY, GuoJ, SuZ: Natural products with anti-obesity effects and different mechanisms of action. J Agric Food Chem, 2016; 64:9571–9585.
9.
SunNN, WuTY, ChauCF: Natural dietary and herbal products in anti-obesity treatment. Molecules, 2016; 21:E1351.
RichardAJ, BurrisTP, Sanchez-InfantesD, WangY, RibnickyDM, StephensJM: Artemisia extracts activate PPARγ, promote adipogenesis, and enhance insulin sensitivity in adipose tissue of obese mice. Nutrition, 2014; 30:31–36.
12.
OhJH, KaradenizF, LeeJI, SeoY, KongCS: Artemisia princeps inhibits adipogenic differentiation of 3T3-L1 pre-adipocytes via downregulation of PPARγ and MAPK pathways. Prev Nutr Food Sci, 2019; 24:299–307.
13.
KimMJ, ParkMH, JeongMK, et al.: Radical scavenging activity and anti-obesity effects in 3T3-L1 preadipocyte differentiation of ssuk (Artemisia princeps pamp.) extract. Food Sci Biotechnol, 2010; 19:535–540.
14.
JungUJ, BaekNI, ChungHG, et al.: The anti-diabetic effects of ethanol extract from two variants of Artemisia princeps pampanini in C57BL/ksj-db/db mice. Food Chem Toxicol, 2007; 45:2022–2029.
15.
YangHG, KimHJ, KimHS, ParkSN: Antioxidative and antibacterial activities of Artemisia princeps pampanini extracts. Korean J Microbiol Biotechnol, 2012; 40:250–260.
16.
LeeSH, BaeEA, ParkEK, et al.: Inhibitory effect of eupatilin and jaceosidin isolated from Artemisia princeps in IgE-induced hypersensitivity. Int Immunopharmacol, 2007; 7:1678–1684.
17.
AlbuquerqueMRJR, PiresAML, PessoaODL, SilveiraER: Terpenoids, flavonoids and other constituents of Eupatorium betonicaeforme (Asteraceae). J Braz Chem Soc, 2006; 17:68–72.
18.
BohlmannF, ZderoC, LonitzM: Neue guajen-derivate aus parthenium hysterophorus und ein weiteres pseudoguajanolid aus ambrosia cumanensis. Phytochemistry, 1977; 16:575–577.
19.
NishimuraS, ManabeI, NagasakiM, et al.: Adipogenesis in obesity requires close interplay between differentiating adipocytes, stromal cells, and blood vessels. Diabetes, 2007; 56:1517–1526.
20.
KimMJ, JangWS, LeeIK, et al.: Reciprocal regulation of adipocyte and osteoblast differentiation of mesenchymal stem cells by Eupatorium japonicum prevents bone loss and adiposity increase in osteoporotic rats. J Med Food, 2014; 17:772–781.
21.
RochaDD, DantasINF, AlbuquerqueMRJR, et al.: Studies on the cytotoxicity of miscellaneous compounds from Eupatorium betonicaeforme (d.c.) Baker (Asteraceae). Chem Biodivers, 2007; 4:2835–2844.
22.
LinWY, KuoYH, ChangYL, et al.: Anti-platelet aggregation and chemical constituents from the rhizome of Gynura japonica. Planta Med, 2003; 69:757–764.
23.
KawaiM, RosenCJ: PPARγ: A circadian transcription factor in adipogenesis and osteogenesis. Nat Rev Endocrinol, 2010; 6:629–636.
24.
SongY, KimMB, KimC, KimJ, HwangJK: 5,7-dimethoxyflavone attenuates obesity by inhibiting adipogenesis in 3T3-L1 adipocytes and high-fat diet-induced obese C57BL/6j mice. J Med Food, 2016; 19:1111–1119.
25.
JohnsonGL, LapadatR: Mitogen-activated protein kinase pathways mediated by ERK, JNK, and p38 protein kinases. Science, 2002; 298:1911–1912.
26.
BostF, AouadiM, CaronL, BinétruyB: The role of MAPKs in adipocyte differentiation and obesity. Biochimie, 2005; 87:51–56.
27.
EngelmanJA, LisantiMP, SchererPE: Specific inhibitors of p38 mitogen-activated protein kinase block 3T3-L1 adipogenesis. J Biol Chem, 1999; 273:32111–32120.
28.
ChenY, AlmanBA: Wnt pathway, an essential role in bone regeneration. J Cell Biochem, 2009; 106:353–362.
29.
BennettCN, LongoKA, WrightWS, et al.: Regulation of osteoblastogenesis and bone mass by Wnt10b. Proc Natl Acad Sci USA, 2005; 102:3324–3329.
30.
CawthornWP, BreeAJ, YaoY, et al.: Wnt6, Wnt10a and Wnt10b inhibit adipogenesis and stimulate osteoblastogenesis through a β-catenin-dependent mechanism. Bone, 2012; 50:477–489.
31.
KrishnanV, BryantHU, MacDougaldOA: Regulation of bone mass by Wnt signaling. J Clin Invest, 2006; 116:1202–1209.
32.
PantovicA, KrsticA, JanjetovicK, et al.: Coordinated time-dependent modulation of AMPK/Akt/mTor signaling and autophagy controls osteogenic differentiation of human mesenchymal stem cells. Bone, 2013; 52:524–531.
33.
ZhaoJ, YueW, ZhuMJ, SreejayanN, DuM: AMP-activated protein kinase (AMPK) cross-talks with canonical Wnt signaling via phosphorylation of β-catenin at ser 552. Biochem Biophys Res Commun, 2010; 395:146–151.
34.
GaoY, ZhouY, XuA, WuD: Effects of an AMP-activated protein kinase inhibitor, Compound c, on adipogenic differentiation of 3T3-L1 cells. Biol Pharm Bull, 2008; 31:1716–1722.
35.
EmamiS, GhanbarimasirZ: Recent advances of chroman-4-one derivatives: Synthetic approaches and bioactivities. Eur J Med Chem, 2015; 93:539–563.
36.
KeriRS, BudagumpiS, PaiRK, BalakrishnaRG: Chromones as a privileged scaffold in drug discovery: A review. Eur J Med Chem, 2014; 78:340–374.
37.
SanzJF, Garcia-LlisoV, MarcoJA, Valles-XirauJ: A cadinane derivative from Artemisia crithmifolia. Phytochem, 1991; 30:4167–4168.