
Abstract
Mulberry (Morus alba L.) fruits have long been used in traditional medicine and as edible berries in many countries. This study investigated the antiadipogenic effect of high hydrostatic pressure mulberry fruit extract (MFE) during 3T3-L1 adipocyte differentiation. MFE decreased lipid and triglyceride accumulation and glycerol-3-phosphate dehydrogenase activity. The mRNA expression levels of genes related to adipogenesis, such as the adipocyte protein 2, proliferator-activated receptor-γ, and CCAAT/enhancer binding protein-α, were suppressed by MFE. They also reduced microRNA (miR)-21 and miR-143 expression, which are involved in adipogenesis. In contrast, adenosine monophosphate-activated protein kinase (AMPK) activity was increased by MFE. These results suggested that MFE may suppress adipogenesis through modulating miR-21/143 expression and AMPK activity in 3T3-L1 adipocytes, which may be useful as antiobesity food agents.
Introduction
Adipocytes regulate energy homeostasis in the body by the release of free fatty acids and neutral fat. 1 Excessive fat accumulation causes obesity, which is one of the causes of various diseases including cardiovascular disease, diabetes, hypertension, and hyperlipidemia. 2 Therefore, there is a growing interest in natural substances that inhibit adipogenesis to prevent obesity.
Mulberries (Morus alba L.) have long been used as conventional medicine in Asian countries. Mulberry is rich in carbohydrates, proteins, fats, fibers, minerals, and vitamins, along with their precursors, and has a low calorie content. 3 Mulberry fruits have been commonly eaten as jams, juices, and fresh fruits, and contains biologically active components that are beneficial for health. 4,5 Mulberry fruit has also been reported to have various pharmacological effects such as hypolipidemia, antidiabetic, antioxidant, and anti-inflammatory effects. 6 –8 These pharmacological effects can be due to polyphenolic compounds, such as flavonoids and anthocyanins. Mulberry fruit has antioxidative properties, mainly due to the presence of polyphenols. 9 In particular, anthocyanins in mulberry fruit have been shown to inhibit the oxidation of free radicals. 10 The freeze-dried mulberry fruit powder improved antioxidant status of blood and hepatic superoxide dismutase and glutathione peroxidase activity in rats fed high-fat diet. 6 The ethyl acetate extract of the mulberry fruit exhibited radical scavenging activity and α-glucosidase inhibitory activity in vitro, and increased antioxidant enzyme activity in streptozotocin-induced diabetic mice. 7
Recently, high hydrostatic pressure (HHP) extract of mulberry fruit have been shown to exhibit anti-inflammatory effects by inhibiting inflammatory mediator synthesis in LPS-stimulated RAW 264.7 cells. 8 HHP belongs to the category of nonthermal treatment techniques and has been reported as an appropriate technology for food processing to extract active compounds of plants by preserving heat-sensitive active compounds to improve their biological activity. 11,12 In the HHP process, temperature and pressure combinations of different conditions can be used for a beneficial effect on the taste, color, and texture of the food. 13 In particular, it is possible to extract at low temperatures without the destruction of heat-sensitive biologically active components. 14
In this study, we examined the antiadipogenic effect of HHP mulberry fruit extract (MFE) in 3T3-L1 adipocytes. Furthermore, we investigated adenosine monophosphate-activated protein kinase (AMPK) activity as well as microRNA (miR)-21/143 expression in 3T3-L1 adipocytes.
Materials and Methods
Materials
MFE preparation
The HHP MFE was kindly supplied by the Korea Food Research Institute (KFRI, Wanju, Korea) using mulberry fruit (Sang-ju Silkworm Farming Association, Sang-ju, Korea) collected from May to June. 8
Mulberry fruits (500 g) were homogenized and mixed with enzymes of the Pectinex ultra color and Pectinex BE XXL. High-pressure enzyme treatment was carried out using a high-pressure apparatus (TFS-50L; Innoway Co., Bucheon, Korea) at 100 MPa for 4 h at 50°C.
The mixtures were transferred to a high-pressure apparatus (TFS-50L; Innoway Co., ) and extracted at 100 MPa for 4 h at 50°C. The enzyme was inactivated by heating at 100°C. for 10 min. The extract was filtered and then freeze-dried.
Total phenolic, flavonoid, and anthocyanin contents
The total phenolic content was analyzed by the method of Folin-Denis 16 and expressed in milligram gallic acid equivalent per 100 g of sample. Total flavonoid content was analyzed by the aluminum chloride method 17 and expressed in milligram quercetin equivalent per 100 g of sample. The total anthocyanin content was analyzed by ultraperformance liquid chromatography analysis 8 and expressed as milligram of cyanidin 3-O-glucoside and cyanidin 3-O-rutinoside equivalents per 100 g of sample.
Cell culture
Mouse 3T3-L1 cells (American Type Culture Collection, Manassas, VA, USA) were cultured in Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% (v/v) bovine calf serum, 100 units/mL penicillin, 100 μg/mL streptomycin, and 2 mM glutamine under conditions of 37°C and 5% CO2. To induce adipocyte differentiation, confluent 3T3-L1 cells (day 0, d0) were treated with DMEM supplemented with 10% fetal bovine serum (FBS) and differentiation medium (DM) containing 3-isobutyl-1-methylxanthine (IBMX; 0.5 mM), dexamethasone (1 μM), and insulin (5 μg/mL) for 2 days (d2). The cells were incubated with insulin (5 μg/mL) for an additional 2 days (d4), then in DMEM containing 10% FBS for 5 days (d9). MFE was treated for 7 days (d2 to d9).
Cytotoxicity
The adipocyte toxicity of MFE was evaluated using a commercial cell count kit-8 (CCK-8; Dojindo Laboratories, Kumamoto, Japan). 18 3T3-L1 cells were treated with 0 (control), 10, 50, 100, or 500 ng/mL of MFE, and incubated for 1, 2, 5, or 7 days. The absorbance was read using a Varioskan plate reader (Thermo Electron, Waltham, MA, USA) at 450 nm.
Intracellular lipid and triglyceride contents
Lipid content was measured by Oil Red O staining (Sigma-Aldrich, St. Louis, MO, USA). 18 Triglyceride (TG) level was analyzed using a commercial TG assay kit (EMBIEL Co., Ltd., Gunpo, Korea). 18 Protein concentrations were analyzed using a bicinchoninic acid protein assay kit (Thermo Scientific, Pittsburgh, PA, USA), and then normalized to protein content.
Glycerol-3-phosphate dehydrogenase and AMPK activity
Glycerol-3-phosphate dehydrogenase (GPDH) activity was analyzed using a commercial kit (Takara, Kyoto, Japan), and measured by monitoring the decrease of nicotinamide adenine dinucleotide (NADH) in the presence of dihydroxyacetone phosphate. 18 Absorbance was measured at 340 nm. AMPK activity was performed using an AMPK Kinase Assay kit (CycLex, Nagano, Japan), 18 and measured by monitoring phosphorylation of Ser 789 in IRS-1, using an antimouse phospho-Ser 789 IRS-1 monoclonal antibody and peroxidase-coupled antimouse IgG. Absorbance was measured at 450 nm.
mRNA and miR expression
The mRNA and miR expression levels were analyzed by quantitative real-time PCR (qRT-PCR) as described previously. 19 Total RNA from 3T3 adipocytes was extracted using TRIzol reagent (GeneAll Biotechnology, Seoul, Korea). cDNA for mRNA and miR was synthesized using M-MLV Reverse Transcriptase (Bioneer, Daejeon, Korea) and miRNA cDNA Synthesis kit with Poly (A) Polymerase Tailing (ABM, Inc., Richmond, BC, Canada), respectively. The qRT-PCR was performed using Rotor Gene 3000 (Corbett Research, Mortlake, NSW, Australia) and AccuPower 2X Greenstar qPCR Master Mix (Bioneer). Primers used for qRT-PCR analysis are described in Table 1. Quantification of miRs was carried out using miR-21, miR-143, and U6 specific primers (ABM, Inc.,). Data analysis was conducted by the 2−ΔΔCt method. β-actin for mRNA and U6 snRNA for miR were used as the reference gene for normalization, respectively.
Primers used for Quantitative Real-Time Polymerase Chain Reaction
Ap2, adipocyte protein 2; Cebp-α, CCAAT/enhancer binding protein-α; Ppar-γ, proliferator-activated receptor-γ.
Statistical analysis
Values are expressed as mean ± standard error of the mean and statistical analysis was performed using SPSS software version 25 (IBM Corporation, Armonk, NY, USA). Significant differences between different treatment concentrations were analyzed using one-way analysis of variance (ANOVA) followed by Tukey's post hoc analysis for multiple comparisons. P < .05 was determined to be statistically significant.
Results
Total phenolic, flavonoid, anthocyanin contents of MFE
Total phenolics, flavonoids, and anthocyanin contents of MFE are given in Table 2. The total phenolic and flavonoid contents of MFE were 5.16 ± 0.05 and 9.73 ± 0.17 mg per 100 g, respectively. The total anthocyanin content of MFE was 168.86 ± 8.97 mg per 100 g. Cyanidin 3-O-glucoside and cyanidin 3-O-rutinoside in anthocyanins were 97.68 ± 1.27 and 71.18 ± 7.73 mg per 100 g, respectively.
Bioactive Compounds Found from High-Hydrostatic Pressure Extract of Mulberry Fruit
Data are expressed as mean ± SEM. The contents of total phenolics and flavonoid are expressed as milligram of GAE and QE equivalents per 100 g of dry powder. The contents of anthocyanin are expressed as milligram of cyanidin 3-O-glucoside and cyanidin 3-O-rutinoside equivalents per 100 g of dry powder. GAE, gallic acid equivalent; QE, quercetin equivalent.
Effect of MFE on 3T3-L1 cell viability
To evaluate the potential cytotoxicity of MFE on 3T3-L1 cells, cells were incubated with MFE at different concentrations (0 [control], 10, 50, 100, or 500 ng/mL) for 1, 2, 5, or 7 days. After 7 days of incubation, cytotoxicity was unaffected by 0, 10, 50, and 100 ng/mL of MFE (Fig. 1). But, at 500 ng/mL of MFE after incubation for 5 and 7 days, cell viability significantly decreased by 19.7% and 25.0%, respectively, in comparison with control. At treatment concentrations of 10 to 100 ng/mL MFE, cytotoxicity was not observed. Therefore, in future experiments, a nontoxic concentration range <100 μg/mL of MFE was used.
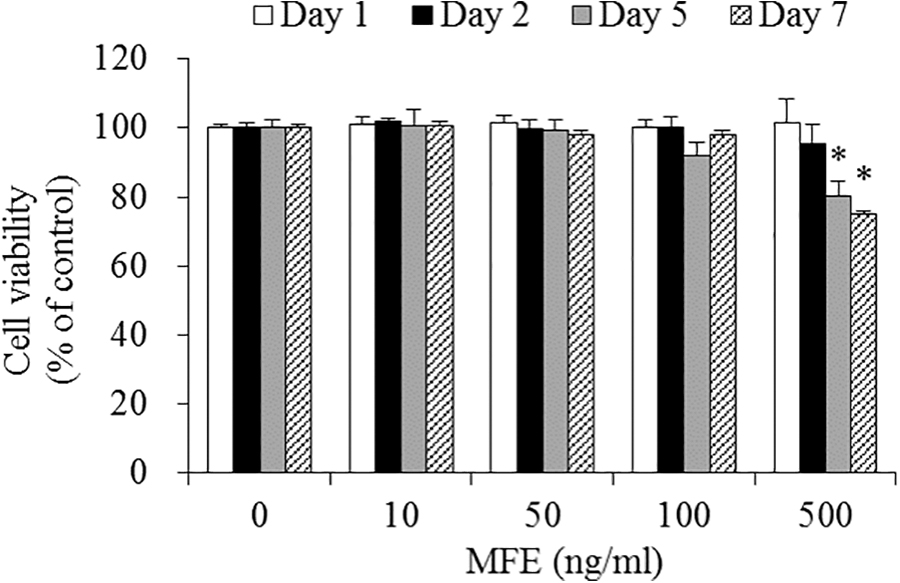
Effects of MFE on viability of 3T3-L1 cells. Cells were treated with 0 (control), 10, 50, 100, or 500 ng/mL of MFE, and incubated for 1, 2, 5, or 7 days. Cell viability was determined using WST-8 assay. Values expressed are averages ± SEM. Statistical analysis was performed using one-way ANOVA, with differences considered significant at *P < .05 versus untreated control. ANOVA, analysis of variance; MFE, mulberry fruit extract; SEM, standard error of the mean; WST, water-soluble tetrazolium salt.
Effects of MFE on lipid and TG contents
To measure the effect of MFE on lipid accumulation during 3T3-L1 adipocyte differentiation, intracellular lipid content was measured using Oil Red O staining. Cells were incubated for 7 days at concentrations of 0 (DM-treated control), 50, and 100 ng/mL MFE. Changes in adipocyte differentiation were observed at day 7 (Fig. 2A). The intracellular lipid content decreased by 10.3% and 20.7% in 50 and 100 ng/mL of MFE, respectively, in comparison with DM-treated control (Fig. 2B). To investigate intracellular TG content, cells were incubated for 4 or 7 days with 0 (DM-treated control) and 100 ng/mL of MFE. At 7 days, MFE reduced the intracellular TG content by 20.6%, in comparison with DM-treated control (Fig. 2C).

Effects of MFE on intracellular lipid and TG accumulation during adipocyte differentiation.
Effect of MFE on GPDH activity
To determine whether MFE affects lipid accumulation inhibition mechanisms, GPDH activity was investigated in 3T3-L1 adipocytes. Cells were incubated for 7 days at concentrations of 0 (DM-treated control), 50, and 100 ng/mL of MFE. GPDH activity was decreased by 17.8% at a concentration of 100 μg/mL MFE, in comparison with DM-treated control (Fig. 3).

Effects of MFE on GPDH activity in adipocytes. 3T3-L1 cells were treated with 0 (DM-treated control), 50, and 100 μg/mL of MFE, and incubated for 7 days. GPDH activity was determined using a GPDH assay kit. Values expressed are averages ± SEM. Statistical analysis was performed using one-way ANOVA, with differences considered significant at *P < .05 and **P < .01 versus DM-treated control. GPDH, glycerol-3-phosphate dehydrogenase.
Effects of MFE on expression of genes related to adipogenesis
We investigated whether MFE reduces lipid accumulation through regulation related to adipogenesis, including, proliferator-activated receptor-γ (Ppar-γ), CCAAT/enhancer binding protein-α (Cebp-α), and adipocyte protein 2 (Ap2) in 3T3-L1 cells. Cells were incubated for 7 days at concentrations of 0 (DM-treated control), 50, and 100 ng/mL of MFE. The mRNA levels of Ppar-γ, Cebp-α, and Ap2 were reduced by 51.3%, 45.0%, and 60.3%, respectively, at a concentration of 100 ng/mL MFE, in comparison with DM-treated control (Fig. 4). In addition, the mRNA level of aP2 decreased by 28.0% at a concentration of 50 ng/mL MFE in comparison with DM-treated control (Fig. 4C).
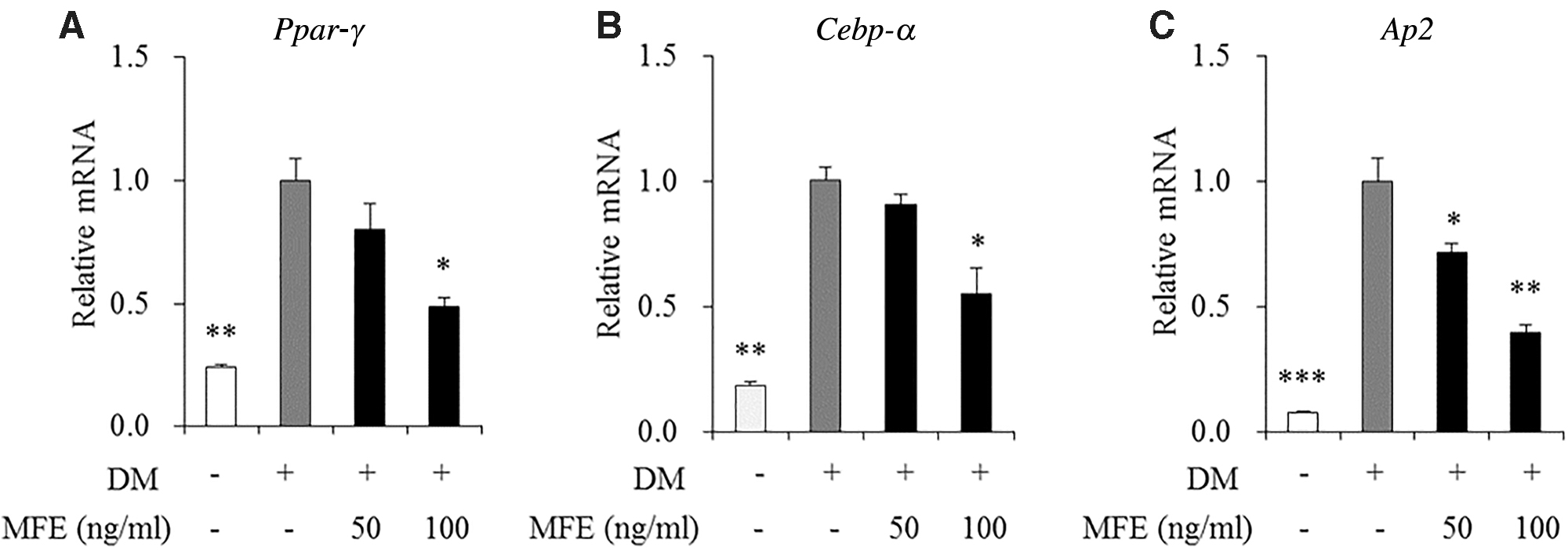
Effects of MFE on the expression of genes involved in adipogenesis in adipocytes. 3T3-L1 cells were treated with 0 (DM-treated control), 50, and 100 μg/mL of MFE, and incubated for 7 days. The mRNA levels of Ppar-γ
Effects of MFE on miR-21 and miR-143 expression
To elucidate the levels of miR-21 and miR-143 expression by MFE in 3T3-L1 cells, cells were incubated for 7 days at concentrations of 0 (DM-treated control), 50, and 100 ng/mL of MFE. The level of miR-21 in 50 and 100 ng/mL of MFE-treated cells significantly decreased by 19.0% and 42.3%, respectively, in comparison with DM-treated control (Fig. 5A). The level of miR-143 in 50 and 100 ng/mL of MFE-treated cells also significantly decreased by 37.3% and 56.7%, respectively, in comparison with DM-treated control (Fig. 5B).

Effects of MFE on miR-21
Effects of MFE on AMPK activity
To assess whether MFE affects the AMPK activity regulation of lipid metabolism in 3T3-L1 adipocytes, cells were incubated for 7 days at concentrations of 0 (DM-treated control), 50, and 100 ng/mL of MFE. The AMPK activity in 100 ng/mL of MFE-treated cells increased by 1.64-fold in comparison with DM-treated control (Fig. 6).
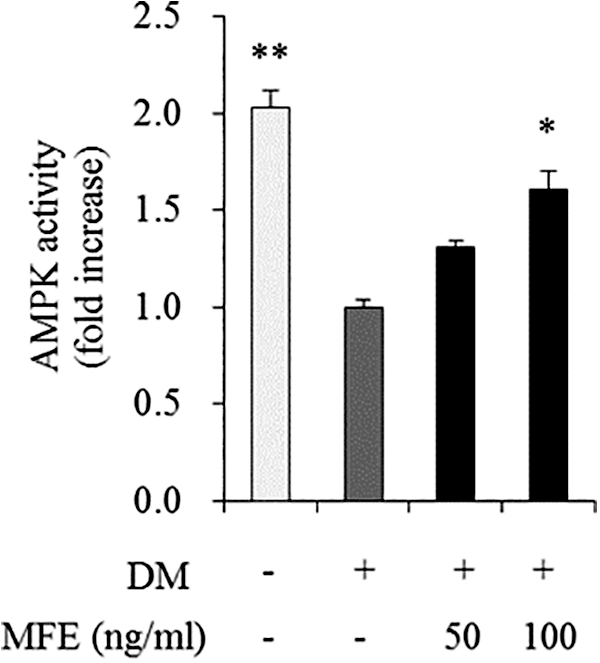
Effects of MFE on AMPK activity in adipocytes. 3T3-L1 cells were treated with 0 (DM-treated control), 50, and 100 ng/mL of MFE, and incubated for 7 days. Values expressed are averages ± SEM. Statistical analysis was performed using one-way ANOVA, with differences considered significant at *P < .05 and **P < .01 versus DM-treated control. AMPK, adenosine monophosphate-activated protein kinase.
Discussion
Adipogenesis is the differentiation process by which preadipocytes change into mature adipocytes, which affects the development of obesity by increasing TG storage. 20 In this study, we investigated the antiadipogenic effect of MFE in 3T3-L1 cells. MFE reduced the intracellular lipid and TG accumulation in 3T3-L1 cells. In a previous study, polysaccharide extracted from Korean mulberry fruits reduced the number of fat cells via suppression of preadipocyte proliferation in 3T3-L1 cells. 21 The ethanolic extracts of mulberry fruits decreased 3T3-L1 adipocyte differentiation. 22 In addition, mulberry fruit water extract decreased the body weight and serum lipid profiles of hamsters fed with a high-fat diet. 23 Our results showed a decrease in intracellular lipid and TG accumulation by MFE during 3T3-L1 adipocyte differentiation, suggesting that MFE may be beneficial for inhibiting fat accumulation.
In the stage of differentiation of preadipocytes, the cells exhibit a change in gene expression pattern, and the expression level of adipogenic transcription factors also changes depending on the differentiation process. In particular, Ppar-γ and Cebp-α are transcriptional regulators that play an important role in adipocyte differentiation, 24,25 and adipose-specific marker Ap2 is regulated through the action of Ppar-γ and Cebp-α. 26 In previous studies, ethyl acetate MFEs have been reported to reduce adipocyte differentiation by inhibiting mRNA expression of Ppar-γ and Cebp-α in 3T3-L1 adipocytes. 22 Ethanol extract of mulberry leaf inhibited lipid accumulation and reduced protein levels of Ppar-γ and Cebp-α in 3T3-L1 adipocytes. 27 Lee et al. reported that anthocyanins isolated from grape inhibit Ppar-γ, Cebp-α, and sterol regulatory element-binding transcription factor 1 (Srebp-1c) during 3T3 adipocyte differentiation. 28 Our findings indicated that MFE suppresses the expression of Ppar-γ, Cebp-α, and Ap2 in 3T3-L1 adipocytes. Thus, it is assumed that MFE can exert antiadipogenic effects by suppressing the expression of adipogenic genes in adipocytes.
miR is a small noncoding RNA consisting of 20–24 nucleotides in various biological processes and affects adipose tissue as a major regulator of lipid metabolism in obesity and obesity-related diseases. 29,30 MiR-21 promoted adipocyte differentiation, increased expression in adipose tissue of obese animals, and miR-143 also induced adipocyte differentiation. 31 –35 We previously found that HHP extract of ginger suppressed miR-21 and miR-132 expression and increased AMPK activation in adipose tissue of obese rats. 19 In addition, Zerumbone, a sesquiterpene isolated from ginger rhizome (Zingiber zerumbet), has been shown to decrease the miR-143 expression and increase AMPK phosphorylation in adipose tissue of obese mice. 36 However, the regulatory mechanism for miRs of MFE has not yet been elucidated. In this study, we found for the first time that MFE reduced miR-21 and miR-143 expression levels in 3T3-L1 adipocytes. Thus, it can be speculated that the antiadipogenic effects of MFE may be partially related to the inhibition of miR-21 and miR-143 expression in adipocytes.
AMPK is a key energy sensor important for the regulation of fat production and lipid metabolism and its activation inhibits adipocyte differentiation with inhibition of Ppar-γ, Cebp-α, and Srebp-1c. 37,38 A recent report showed that MFE containing anthocyanins ameliorated insulin sensitivity via AMPK activation in skeletal muscles of diabetic db/db mice. 39 Mulberry water extracts have been shown to prevent liver damage through hepatic AMPK activation in C57BL/6J mice. 40 Our findings indicated that MFE increased AMPK activation in 3T3-L1 adipocytes, suggesting that there is an antiadipogenic effect of MFE through AMPK activation in adipocytes.
In conclusion, our findings suggest that MFE inhibits adipogenesis in 3T3-L1 cells. MFE-mediated regulation of adipogenesis may be associated with AMPK activation as well as inhibition of the expression of the adipogenic genes and miRNAs (miR-21 and miR-143) (Fig. 7). Therefore, MFE may be used as a beneficial food agent for preventing and/or treating obesity.
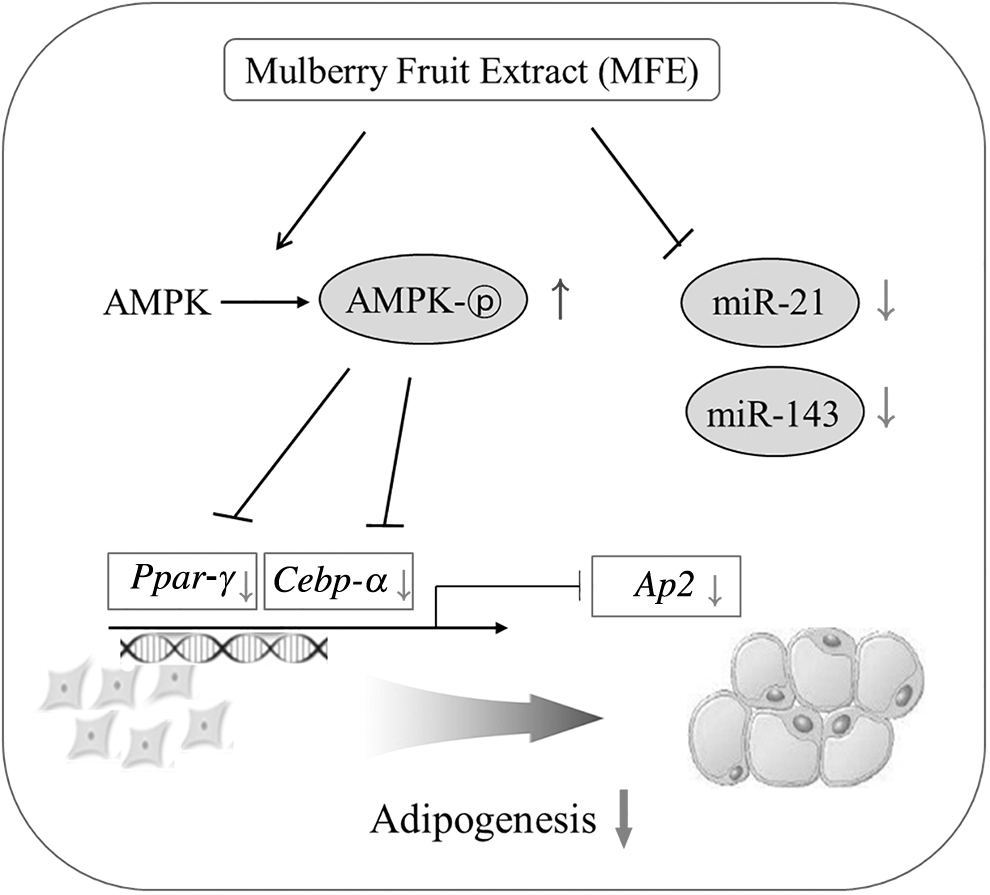
Schematic mechanistic diagram showing the potential cellular exchange effect of MFE on adipogenesis.
Footnotes
Author Disclosure Statement
The authors declare no conflict of interest.
Funding Information
This study was supported by the National Research Foundation of Korea (NRF) funded by the Korean Government (Grant Nos. 2012M3A9C4048761, 2016R1A2B4011021, and 2019R1A2C1002861).