
Abstract
Trillions of microorganisms reside in the hosts' gut. Since diverse activities of gut microbiota affect the hosts' health status, maintenance of gut microbiota is important for maintaining human health. Green tea (GT) has multiple beneficial effects on energy metabolism with antiobesity, antidiabetic, and hypolipidemic properties. As GT contains a large amount of bioactive ingredients (e.g., catechins), which can be metabolized by microorganisms, it would be feasible that consumption of GT may cause compositional changes in gut microbiota, and that the changes in gut microbiota would be associated with the beneficial effects of GT. In this study, we demonstrated that consumption of GT extract relieves high-fat diet-induced metabolic abnormalities. Interestingly, GT administration significantly encouraged the growth of Akkermansia muciniphila (Akkermansia), a beneficial microorganism to relieve obesity and related metabolic disorders. Finally, we found that epigallocatechin gallate is the component of GT that stimulates the growth of Akkermansia. According to these data, we propose that GT could be a prebiotic agent for Akkermansia to treat metabolic syndromes.
Introduction
Gut microbiota are the resident microorganisms in the gastrointestinal tract of the hosts. Since the role of gut microbiota on diet-induced obesity was discovered, 1 –3 numerous studies have investigated the relationship between gut microbiota and human health. As a result, specific changes in gut microbiota are reported to be closely associated with metabolic syndromes. For example, the composition of gut microbiota from lean individuals is very different from that of obese 4 or diabetic individuals. 5 In addition, increased abundance of Firmicutes phylum in obese mice leads to enhanced energy harvest from the foods. 1 Finally, loss of body weight restores compositional change of Firmicutes and Bacteroidetes phyla in animals 6 and humans. 4 On the other hand, lactic acid-producing bacteria (e.g., Lactobacillus) are introduced to modulate immune function, 7,8 whereas certain types of microorganisms—mainly belonging to Proteobacteria—trigger diverse infectious diseases including diarrhea and sepsis. In addition, those microorganisms secrete extracellular vesicles (EVs), exosome particles containing donor cell-derived components such as proteins, nucleic acids, cellular organelles, and/or metabolites. As EVs carry various substances, EVs are supposed to mediate diverse actions of microorganisms that affect the host's metabolism. For example, EVs from Staphylococcus aureus cause proinflammatory responses, 9,10 whereas Lactobacillus-derived EVs can protect atopic dermatitis that is driven by S. aureus EVs. 11 As microorganisms control biological actions of the host through diverse methods including EVs, increasing beneficial microbiota in the gut are crucial to extend an individual's health span.
The composition of gut microbiota is influenced by environmental changes, such as the sort of meals, functional foods, pharmaceutical use, and other causal factors. As we consume average three meals per day, food consumption would be one of the most powerful factors for modulating the growth of gut microbiota among the causative aspects. Green tea (GT) is the representative oriental tea, which is made from Camellia sinensis. GT contains a large amount of phytochemicals, 12 and the benefits of GT consumption have been widely studied. Antioxidant, anticancer, and anti-inflammatory effects are representative properties of GT. 13 –20 However, the relationship between GT consumption and gut microbiota composition has not been researched extensively.
Recently, Akkermansia muciniphila (Akkermansia), a mucin-degrading bacterium, 21 has been shown to relieve various metabolic disorders. 22 –28 Interestingly, Akkermansia is also reported to secrete EVs, and Akkermansia-derived EVs mediate beneficial effect of Akkermansia, at least in part. 29 As Akkermansia consumes mucins in the intestine, hosts are forced to produce new mucin. Through the endless renewal of mucins in the intestinal tract, Akkermansia helps maintain barrier and immune function in the gut. Chronic inflammation leads to the development of insulin resistance and metabolic disorders, 30 –32 and it is feasible that Akkermansia could be used to treat diverse metabolic syndromes such as obesity, insulin resistance, and type 2 diabetes by suppressing inflammation in the gut. However, little is known about how to encourage Akkermansia growth to relieve metabolic disorders.
In this study, we confirmed that administration of GT extract prevents diet-induced obesity and obesity-related metabolic disorders. GT alleviated excessive weight gain, which was induced by high-fat diet (HFD). GT also improved blood glucose concentrations and insulin tolerance and inflammation. Interestingly, we found that consumption of GT caused a robust increase of Akkermansia, and epigallocatechin gallate (EGCG) would be the mediator of the growth of Akkermansia. Taken together, we propose that GT is a modulator of the gut environment and mitigates obesity and related metabolic diseases.
Materials and Methods
Preparation of GT extract
GT extract was manufactured and kindly provided from Borak (Seoul, Korea). In brief, dried leaves of GT were extracted with 50% ethanol (15.78 times w/w of dried GT leaf) at 70°C for 2 h, followed by freeze drying. The content of catechins in GT was determined by HPLC (Waters Alliance 2695, Waters, Milford, MA, USA) using an octadecyl-silica (C18) column, as described earlier. 6 A gradient elution was performed with various compositions of solvent A (water–acetic acid with 96.86:3.14 w/w) to solvent B (methanol), with a flow rate of 1 mL/min as follows: 0–25 min, 80:20; 26–50 min, 70:30; 51–55 min, 80:20. The wavelength of acquisition was set in the range of 280 nm. Total content of catechin of the GT extract was ∼40% w/w, and EGCG comprised ∼47% of total catechins (Table 1).
Composition of Catechins in Green Tea
EGCG, epigallocatechin gallate; EGC, epigallocatechin; ECG, epicatechin gallate; EC, epicatechin; GCG, gallocatechin gallate; GC, gallocatechin; CG, catechin gallate; C, catechin; N.D, not detected.
Animal experiment
The animal experiment was approved by Aestura Institutional Animal Care and Use Committee (IACUC15-037). Seven-week-old male C57/BL6 mice were purchased from Samtako (Osan, South Korea) and maintained on a 12 h light/dark cycle with a controlled temperature (21–25°C) and humidity (50–60%). After a 1 week adaptation period, mice were randomly divided into four groups (normal diet (ND)/vehicle (−), ND/GT, HFD/(−), and HFD/GT, n = 16 per group). The HFD/(−) and HFD/GT groups were fed a diet of 60% calories from fat (Research Diet, New Brunswick, NJ, USA), and ND/(−) and ND/GT groups were fed AIN-76A diet (Research diet) for 12 weeks. During the experiment, GT (400 mg/kg dissolved in water) or vehicle (water) was orally administered daily. Body weight and food intake were measured weekly.
Glucose tolerance test and insulin tolerance test
Glucose tolerance test (GTT) and insulin tolerance test (ITT) were performed as described previously. 33 In brief, mice were fasted overnight. After overnight fasting, plasma glucose concentrations were measured by using Therasense FreeStyle Meter kit (Therasense, Uppsala, Sweden). For GTT assay, glucose (1 g/kg; Sigma-Aldrich, St. Louis, MO, USA) was administered orally. For ITT assay, 1 U of insulin (Sigma-Aldrich) was injected intraperitoneally. After 15, 30, 60, and 120 min, blood sample was prepared from tail, and plasma glucose level was measured as described previously.
Plasma analysis
Plasma concentrations of triglyceride, total cholesterol, and free fatty acid (FFA) were analyzed by using Triglyceride Reagent (Sigma-Aldrich), Cholesterol/Cholesteryl Ester Assay Kit (Abcam, Cambridge, United Kingdom), and Free Fatty Acid Assay kit (Abcam), respectively. Plasma concentration of proinflammatory cytokines (tumor necrosis factor α [TNFα], interleukin-1β [IL-1β], and IL-6) was determined by using ELISA kits (R&D Systems, Minneapolis, MN, USA), following the manufacturer's protocol.
Determination of gene expression
mRNA was isolated by use of Trizol™ reagents (Thermo Fisher Scientific, Waltham, MA, USA). One microgram of each RNA was used to synthesize cDNA using RevertAid™ 1st Strand cDNA Synthesis Kit (Thermo Fisher Scientific). Relative mRNA level was analyzed by using quantitative real-time polymerase chain reaction (qPCR) device (CFX96; Bio-Rad, Hercules, CA, USA) with appropriate primers (Bioneer, Daejeon, South Korea). The sequences of primers used in the qPCR assay are listed in Supplementary Table S1.
Analysis of gut microbiota and EVs
Isolation of bacterial DNA and EVs from fecal samples and analysis of composition of gut microbiota and gut microbiota-derived EVs were conducted at MD Healthcare Corporation (Seoul, South Korea). 34 In brief, collected stool samples were dissolved in phosphate-buffered saline and centrifuged at 10,000 g for 10 min at 4°C. EVs were collected from supernatant fraction, and bacterial fraction was remained in pellet. DNA from microorganisms and EVs was extracted with PowerSoil DNA isolation kit (MO BIO Laboratory, Carlsbad, CA, USA) following the manufacturer's protocol. Bacterial 16s rDNA V3/V4 region from isolated DNA was amplified by PCR, and sequences were further analyzed by using MiSeq system (Illumina, San Diego, CA, USA). Taxonomic assignment was profiled with MDx-Pro ver.1 (MD Healthcare, Inc., Seoul, Korea).
Statistical analysis
Error bars represent SE. A P-value <.05, calculated by two-way ANOVA followed by Tukey honest significant difference, was regarded as statistically significant. Principle component analysis of gut microbiota was conducted by use of R ver. 3.6.3 (R Foundation, Vienna, Austria), and 3D scatter plot was generated using Chartstudio (Plotly, Waltham, MA, USA).
Results
GT relieves diet-induced obesity and hyperlipidemia
To determine whether GT relieves diet-induced obesity, we administered GT to mice fed a HFD. As expected, GT blunted excessive weight gain (Fig. 1A, B) without changing food intake (Fig. 1C). Concomitant with the retardation of body weight increase, the mass of white adipose tissue (WAT) was also reduced partially (Fig. 1D). Plasma lipid levels were also reduced by GT administration (Fig. 1E). During the animal experiment, plasma level of glutamic oxaloacetic transaminase (GOT) and glutamic-pyruvate transaminase (GPT), indicators of liver damage, was induced by HFD (Supplementary Fig. S1). However, treatment of GT did not alter GOT and GPT level compared with vehicle groups (Supplementary Fig. S1). Furthermore, the GOT/GPT ratio was not changed significantly either (data not shown), indicating that GT administration did not provoke liver toxicity. According to these data, it is feasible that GT exhibits an antiobesity and hypolipidemic effect without the induction of hepatotoxicity.
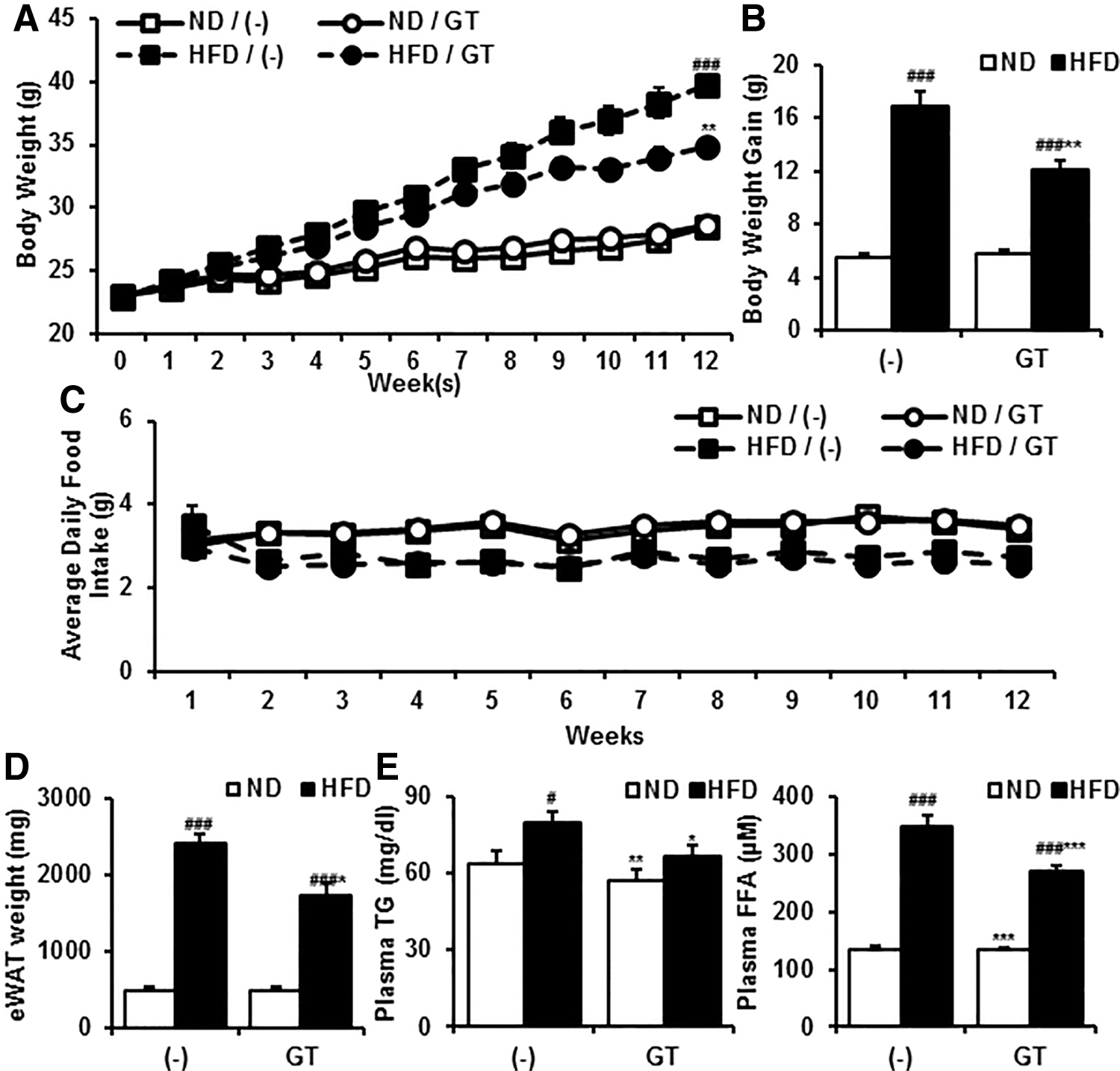
Antiobesity effect of GT.
GT improves glucose and insulin tolerance
Obesity is considered as a causal factor of various metabolic disorders by provoking glucose and insulin intolerance. 31,32 Therefore, we conducted GTT and ITT assays to confirm whether GT improves glucose and insulin tolerance. In the normal diet condition, treatment with GT did not affect glucose and insulin tolerance (Fig. 2). However, interestingly, GT alleviated glucose and insulin resistance in diet-induced obese mice (Fig. 3). Taken together, it is likely that GT relieves glucose and insulin intolerance as well as diet-induced obesity.

GT does not improve glucose and insulin tolerance in normal diet. Mice were subjected to GTT and ITT assays under normal diet condition. GTT result is shown in
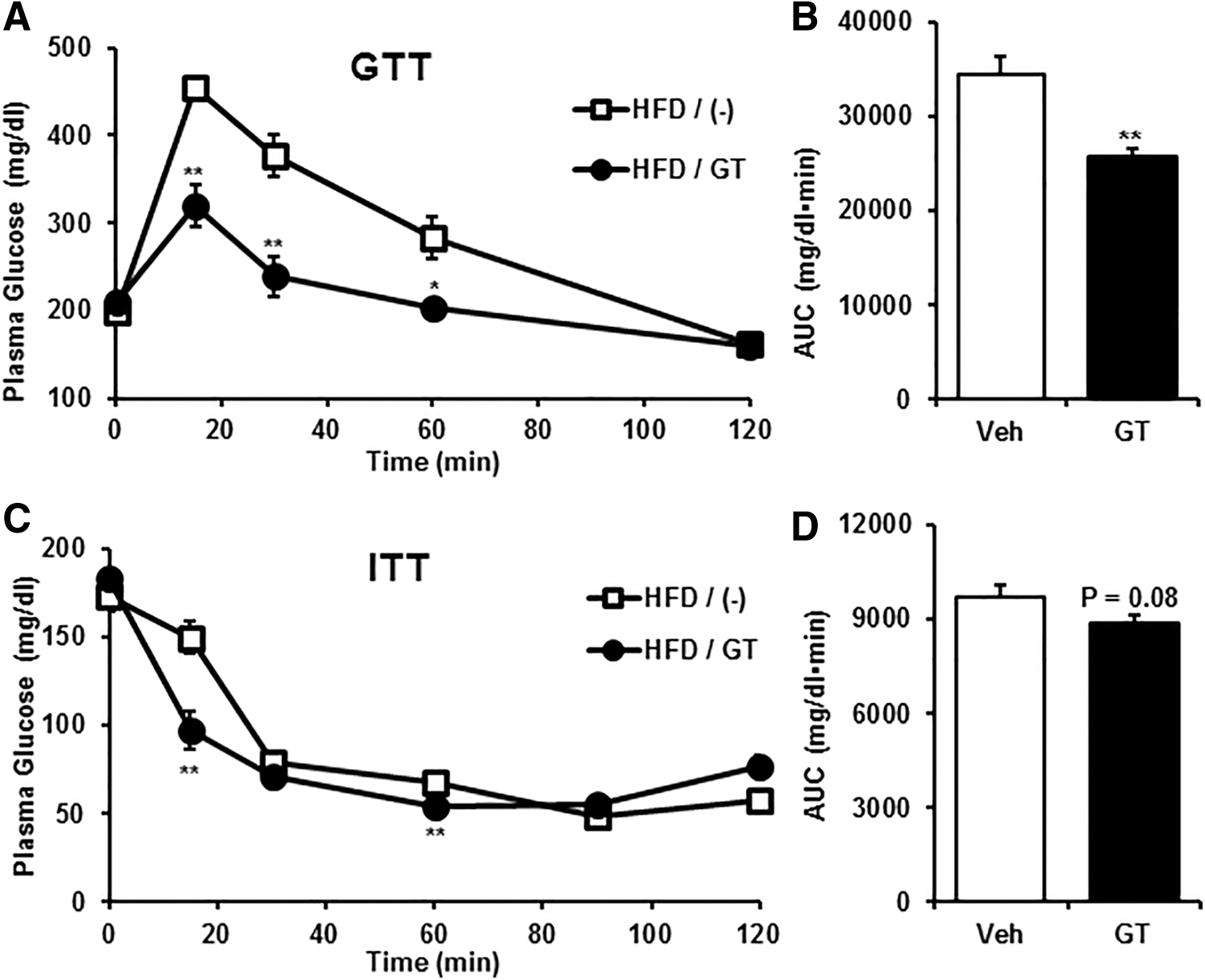
GT relieves glucose intolerance and insulin sensitivity in HFD-induced obese mice. High-fat-fed obese mice were subjected to GTT
GT reduces expression of inflammatory cytokines
By provoking insulin resistance, chronic inflammation plays an important role in the development of metabolic diseases. 30 As GT improved glucose and insulin tolerance in HFD-fed animals (Fig. 3), we further measured protein and mRNA levels of proinflammatory genes to find out whether GT exerts anti-inflammatory effect. As shown in Figure 4A, plasma concentration of proinflammatory cytokines was decreased by GT administration. Parallel with the circulating level, mRNA expression of proinflammatory genes in WAT was reduced by GT consumption (Fig. 4B). Considering the role of inflammatory response on the development of insulin resistance, it is likely that anti-inflammatory property of GT might contribute to the alleviation of HFD-induced metabolic abnormalities such as elevated blood glucose concentrations and insulin intolerance, at least in part.

GT blunts expression of inflammatory genes. Concentration of inflammatory cytokines in plasma
GT alters composition of gut microbiota and encourages the growth of Akkermensia
Since the development of obesity and the compositional change of gut microbiota are closely related, 2,3 it is of interest to examine proportional change of gut microbiota induced by GT consumption. As shown in Figure 5, β-diversity of gut microbiota composition seems to be distinguished by the food source. Interestingly, administration of GT was not related to the changes of gut microbiota in normal diet. Instead, GT reformed composition of microorganisms in the gut, which was induced by HFD (Fig. 5). Our results suggest the possibility that GT could be a causal factor for the compositional change of gut microbiota, in concordance with dietary change.

PCA plot of gut microbiota. The signification of taxonomic variation was determined by PCA as described in Materials and Methods section and shown as 3D scatter plots in phylum
As we confirmed that GT is available to induce changes of gut microbiota, we further analyzed the composition of gut microbiota. At the phylum level, composition of gut microbiota was altered by HFD, especially in the reduction of Verrucomicrobia and TM7 (Fig. 6A). In contrast, the proportion of Firmicutes and Bacteroidetes phyla was increased (Fig. 6A). Interestingly, decreased abundance of Akkermansia by HFD was dramatically reversed by GT administration (Fig. 6B), suggesting that Akkermansia would be a major microbiome target of GT to alleviate metabolic disorders.
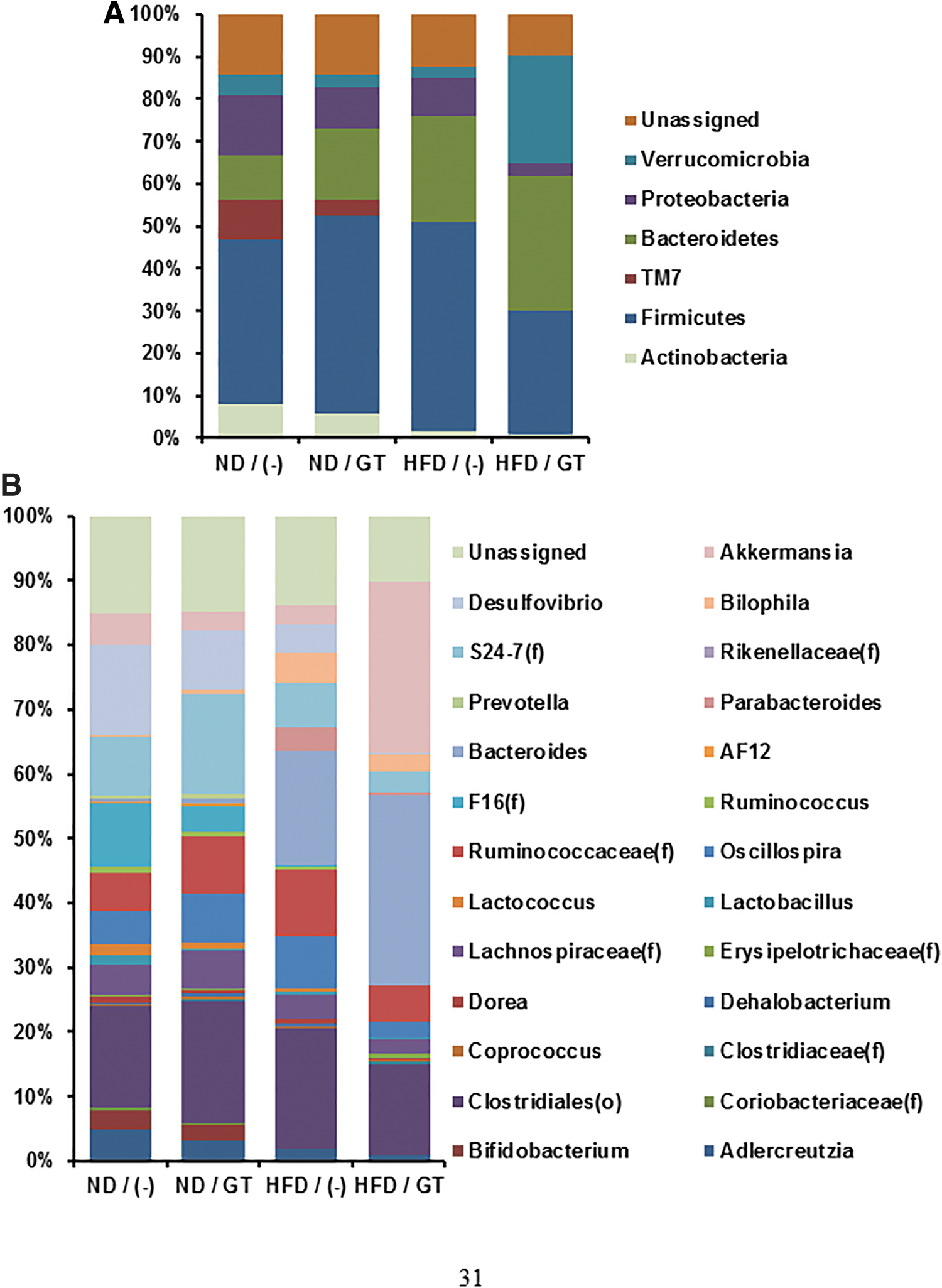
Analysis of gut microbiota composition. Analytical data of gut microbiota composition in phylum
Recently it was reported that secreted vesicles from bacteria mediate the effects of microorganisms. 29,35,36 Thus, we also analyzed EVs from the cecum-derived stools. Concomitant with the compositional data of gut microbiota, abundance of EVs from Akkermansia was decreased by HFD and restored by GT administration (Fig. 7). Interestingly, proportion of proteobacteria-derived EVs was reduced by GT regardless of dietary composition (Fig. 7). Since a number of pathogenic microorganisms belong to proteobacteria, a decreased proportion of EVs from proteobacteria would reflect a GT-mediated anti-inflammatory response (Fig. 4), at least in part. Collectively, we suggest that increased proportion of Akkermansia and its derived EVs would facilitate beneficial effects of GT to relieve obesity and relative complications.
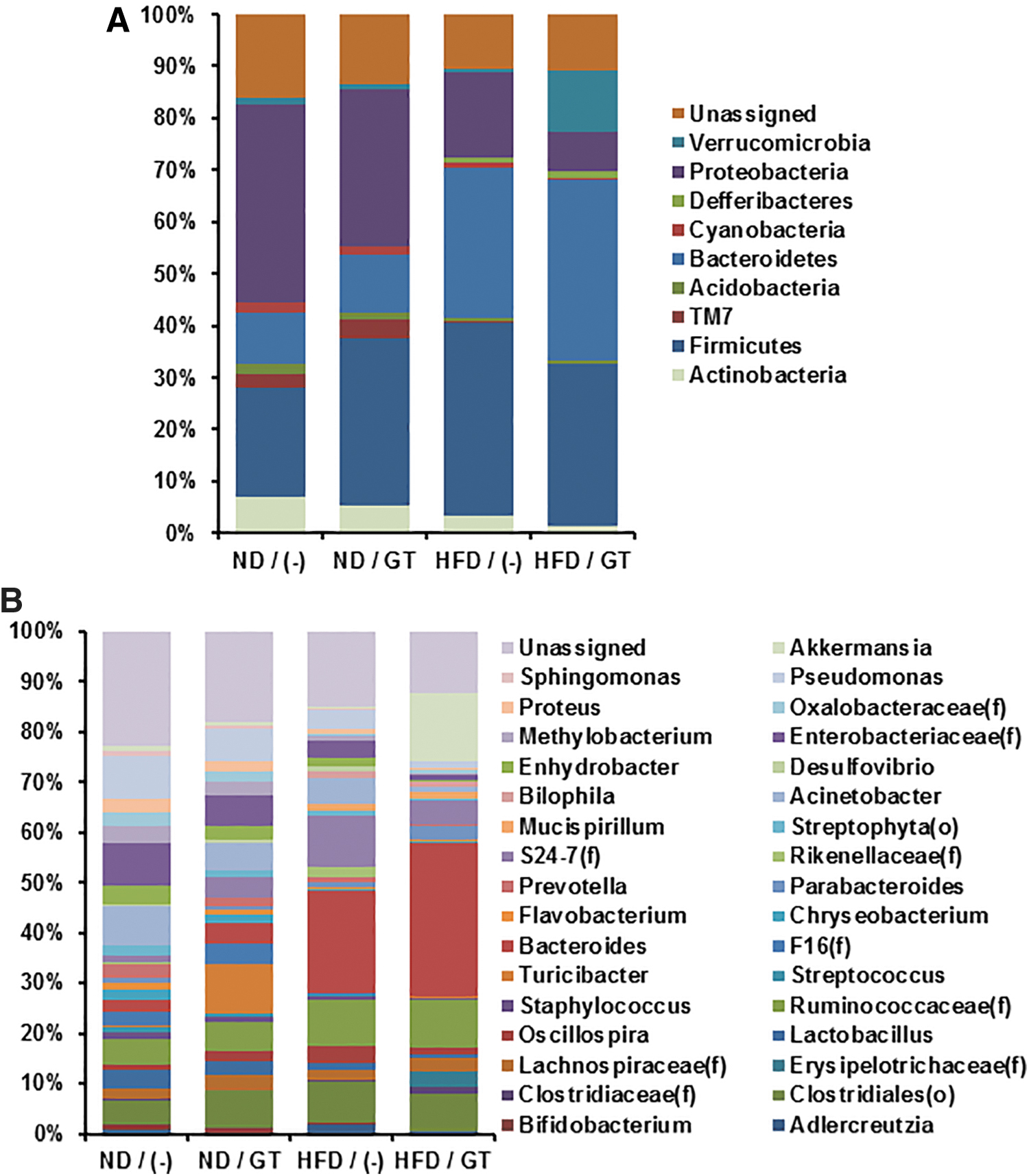
Proportional change of EVs from gut microbiota by green tea consumption. Proportion of microorganism-derived EVs was analyzed in phylum
EGCG governs the effect of GT on Akkermansia growth
GT promoted the growth of Akkermansia despite the high-fat–fed condition. Next, we wondered which compound is the mediator of Akkermansia growth. Since the growth of Akkermansia is encouraged by dietary polyphenols 37,38 and GT contains a large amount of catechins (especially EGCG), we presumed that EGCG would be a major promoter of Akkermansia growth. To determine whether EGCG is the effector molecule of Akkermansia growth, we administered the EGCG (56 mg/kg; the equivalent amount in the GT) and determined the composition of gut microbiota and microbiota-derived EVs. As expected, EGCG was the sufficient factor of Akkermansia growth (Fig. 8A) as well as Akkemansia-derived EV secretion (Fig. 8B). Based on this result, we suppose that EGCG could be the regulatory molecule of GT to alleviate metabolic disorders by encouraging the growth of Akkermansia and secretion of Akkermansia-derived EVs.

EGCG mediates the promotional effect of GT on the growth of Akkermansia and the secretion of Akkermansia EVs. Analytical data of the composition of gut microbiota
Discussion
Previously, it was reported that consumption of GT is associated with specific changes in the gut microbiota. For example, consumption of GT is positively correlated with the abundance of Bifidobacterium species. 39 However, the relationship of GT consumption and gut microbiota composition is not yet thoroughly researched. In this study, we found that treatment with GT improves obesity and related complication and increases the proportion of Akkermansia in diet-induced obese mice, and EGCG plays a facilitating role of Akkermansia growth. Interestingly, administration of GT only augmented the growth of Akkermansia in concordance with HFD. The reason why GT changed the composition of the gut microbiota only in HFD may be due to the stability of the gut environment. Since laboratory animals are maintained in the controlled environment (e.g., temperature, humidity, and composition of meals) for a long time, the composition of gut microbiota remains stable. Therefore, it can be assumed that just an administration of GT is not enough to induce compositional change of gut microbiota. Indeed, source of food is regarded as an important causal factor for the compositional change of gut-resident microorganisms. 40 –42 For these reasons, we assume that GT-mediated changes in the gut microbiota are not observed without dietary change. In contrast, the change of food source (from normal diet to HFD) is enough to disturb gut environment, thereby inducing compositional change of gut microbiota such as decreases in Verrucomicrobia and TM7. During the unstable period of intestinal environment, treatment with GT could affect specific change of the gut environment, especially a robust induction of Akkermansia. Collectively, it is likely that GT can relieve undesirable change of gut microbiota composition that is induced by HFD.
As already mentioned, Akkermansia is reported to be negatively correlated with the progression of obesity and related complications 22 and it is useful to relieve metabolic disorders. 23,25 –27 Moreover, it is also reported that an abundance of Akkermansia and Bifidobacteria is inversely correlated with autism in children. 43 The recent discovery of Akkermansia and its role led us to regard Akkermansia as a novel therapeutic target of various diseases. 44,45 However, the difficulty of the cultivation of Akkermansia limits the development of Akkermansia-targeting therapies. Therefore, identification of growth modulators of Akkermansia would be a valuable discovery to accelerate the development of prebiotic agent of Akkermansia to treat various metabolic disorders.
As well as microorganism itself, secreted EVs can affect the host's metabolism. As already stated, EVs from harmful bacteria can induce inflammatory responses, 9,10 whereas probiotic EVs ameliorate allergic immune responses. 11,36 Moreover, Akkermansia-derived EVs can alleviate colitis, implying that EVs from beneficial microorganisms can be used to improve human health. Owing to the size and their nature, EVs can cross the intestinal barrier to penetrate the host's system, unlike parent cells. Recent studies report the importance of EVs as a therapeutic target, instead of microorganisms. Our study demonstrated that GT and EGCG induced both Akkermansia and its EVs in HFD condition. According to our result, we assume that GT could be commercially used to facilitate EV production from Akkermansia in a bioreactor or culture system.
Some microorganisms contain large amounts of lipopolysaccharides, a ligand of toll-like receptor 4 (TLR4). Activation of TLR4 initiates inflammatory response. 46 Interestingly, FFA can also bind to and activate TLR4, thus it is likely that hyperlipidemia is closely associated with chronic inflammation. 46 Since low-grade chronic inflammation interferes with the insulin signal transduction pathway to trigger diverse metabolic syndromes, 30 it is important to modulate inflammatory responses to treat metabolic disorders. In this experiment, inflammation was induced in the HFD group despite the decreased amount of proteobacteria-derived EVs. This conundrum could be explained by increased FFA concentrations due to the fat-rich diet, at least in part. Elevated circulating FFA level might superfluously counteract the effect of decreased proteobacterial EVs, thereby augmenting proinflammatory responses. Therefore, additional hypolipidemic agents would be needed to modulate proinflammatory responses in obese and hyperlipidemic conditions. In our experiment, GT lowered plasma lipid levels as well as EVs from proteobacteria. By eliminating causal factors for proinflammatory responses, GT exerts a strong anti-inflammatory property to relieve insulin resistance. Since proinflammatory responses affect a number of physiological events including cell proliferation, cell death, oxidative stress, and cancer, 47,48 treatment with GT would be helpful to improve cellular function.
Although we found that GT induces the growth of Akkermansia, it still remains to be elucidated whether the presence of Akkermansia and Akkermansia-derived EVs is needed to exhibit the beneficial effects of GT on obesity and related complications. Extra experiments using Akkermansia-free animals and Akkermansia-derived EVs would be required to prove the relationship of GT-mediated Akkermansia growth and alleviation of metabolic disorders. However, those experiments require highly advanced techniques that are not possible at this time. Instead, when comparing the effects of Akkermansia and GT on energy metabolism and inflammation, it would be reasonable to conclude that Akkermansia mediates the effects of GT on obesity and related metabolic disorders.
Collectively, we demonstrated that GT is a useful agent to relieve metabolic syndromes. We also observed that GT augmented Akkermansia in the gut, and EGCG mediates the promotional effect of GT on the growth of Akkermansia. Regarding the benefits of Akkermansia, we suggest that GT containing EGCG functions as a prebiotic agent to maintain the gut environment and health of the hosts.
Footnotes
Acknowledgments
We appreciate Mr. Jinsang Jung, Mr. Insik Lee, and Mr. Kiyeop Park (Aestura Corporation) for care and assistance in the animal experiment. We also thank members of MD Healthcare, Inc., for the analysis of gut microbiome composition.
Author Disclosure Statement
Hyun Woo Jeong, Jeong Kee Kim, A Young Kim, Donghyun Cho, Ji-Hae Lee, Miyoung Park, and Wangi Kim are employed in Amorepacific Corporation. Jin Kyu Choi is an employee of Aestura Corporation. However, all authors declared that there is no competing interest.
Funding Information
This research was supported by internal research fund (R15E700001) of Amorepacific R&D Center.
Supplementary Material
Supplementary Figure S1
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.