
Abstract
Carrageenan oligosaccharide (CAO), the hydrolysate of carrageenan from marine red algae, is used as a prebiotic additive or medical material. In this study, male Drosophila melanogaster was used as an animal model to explore the possibility that CAO can extend the life span through its relationship with antioxidation, immunity, and gut microbiota in vivo. The results show that a certain amount of CAO effectively prolonged the average life span and improved the climbing vitality and fecundity of male Drosophila. In addition, 0.125% CAO in the diet significantly increased the activity of Cu,Zn-superoxide dismutase (Cu,Zn-SOD) and catalase (CAT), reduced the content of malondialdehyde (MDA), and significantly repressed the expression of nuclear factor kappa B (NF-κB) gene in old male Drosophila tissues. In the intestinal microbiota analysis, 0.125% CAO in the diet increased the diversity of gut microbiota and improved the abundance of Commensalibacter at the genus level in Drosophila on the 40th day. The above results indicated that CAO supplementation could extend the life span of male Drosophila by improving antioxidant activity, immunity, and by regulating intestinal microflora.
Introduction
Red seaweed is the oldest division of marine macrophytes and has attracted extensive attention as an excellent source of nutrients. 1 In recent years, numerous studies have indicated that marine algae polysaccharides extracted from red seaweed have the potential for immune-modulatory, antitumor, and antioxidant biological activity, reflecting the fact that marine algae are superior to terrestrial-derived polysaccharides. 2 –4 However, the disadvantages of polysaccharides are that they are poorly soluble in water and not easily absorbed by the intestine, hindering its further application in functional foods and medicines. Carrageenan, a linear sulfated seaweed polysaccharide, has different structures based on the degree of sulfation 5 and can be hydrolyzed into carrageenan oligosaccharide (CAO) by chemical or enzymatic methods. 6,7 Many in vitro studies have shown that CAOs have anti-inflammatory, antioxidant, and antitumor properties as prebiotics. 8,9 However, there are few studies on the activity in vivo, and the relationship between the antiaging properties of CAO and intestinal microbes has not been reported.
As a novel model animal, Drosophila melanogaster is suitable for use in nutritional intervention studies, as it exhibits many similarities to mammalian species. 10 Although the insect body structure is simpler compared with mammals, the anatomy of Drosophila includes organ systems with equivalent functions of the mammalian heart, lung, kidney, liver, and gonads. Moreover, the Drosophila has a complex and dynamic gut similar in structure and organization to the mammalian gut. 10 Studies have shown that the senescence of Drosophila is gender specific, and female Drosophila is susceptible to estrogen interference, which affects the experimental results. 11,12 Therefore, male Drosophila is more suitable as research objects to investigate the effects of bioactive ingredients on body aging.
During age-related physiological diseases, the intestinal microflora changes significantly, which could affect the health and aging status of the body. 13,14 Furthermore, the bacterial composition and barrier function of the intestine can significantly change before and after senescence in Drosophila. 15 In addition, the intestinal flora is closely related to the immune system, and an imbalance in the microflora leads to inflammatory and metabolic disorders in the body, including obesity, type 2 diabetes, and other metabolic diseases. 16 –18 Aging is a complex physiological process accompanied by oxidative damage of tissues and organs. The accumulation of reactive oxygen species (ROS) is one of the major factors responsible for aging. 19 The most common ROS are superoxide radical (O2−) and hydrogen peroxide (H2O2), which stimulate and cause oxidative damage to cells. 20 However, there are several antioxidants in the body, such as superoxide dismutase (SOD) and catalase (CAT), which can mitigate the damage caused by oxidative stress by neutralizing excessive ROS. 21
In this study, male Drosophila, hereafter just called Drosophila, was used as an animal model to explore the possibility that a certain amount of CAO could extend the life span by affecting antioxidation, immunity, and gut microbiota in vivo (the Drosophila mentioned in the following text was referred to male Drosophila).
Materials and Methods
Reagents
Malondialdehyde (MDA) Assay Kit (TBA method) (A003-1-2), Cu,Zn-Superoxide Dismutase (Cu,Zn-SOD) Activity Assay Kit (A001-2-2), Catalase (CAT) Assay Kit (Visible light) (A007-1-1), and Total Protein Quantitative Assay Kit (A045-2-2) were purchased from Nanjing Jiancheng Bioengineering Co., Ltd. (Nanjing, China).
Drosophila strain and sample preparation
The Canton-S line of D. melanogaster was obtained from the Drosophila Stock Center at Shanghai Academy of Life Sciences, Chinese Academy of Sciences. Pharmaceutical-grade CAO of low molecular weight (≥95%) was purchased from Qingdao Bozhi Huili Biotechnology Co., Ltd., China, and contained polymerization degree 2–6 oligosaccharides obtained by mild acid hydrolysis of carrageenan.
Feeding
According to the procedures of Tang et al., 22 the Drosophila was cultivated at 24 ± 1°C and 55% relative humidity on a 12-h light/12-h dark cycle and reared on various media in this experiment. The basal Drosophila diet consisted of 102 g corn, 78 g sucrose, 20 g yeast, 20 g agar, and 6 mL propionic acid mixed with hot water to prepare 1000 mL of medium. The experimental group diet supplemented with 0.0625%, 0.125%, and 0.25% COS in the basal diet. The experiment was divided into four groups: the basal diet control group and three experimental groups with different doses of CAO. In vivo experiments were performed in the early stage to determine the optimum concentration for administration according to the following formula: experimental in vitro drug concentration μg/mL = 50 × D/5000 ÷ 50% × 103 (D, clinical dose in mg·kg/day). The optimal dose of CAO for the experimental Drosophila was calculated to be 0.125%. Therefore, we selected CAO in high, medium, and low doses (0.25%, 0.125%, and 0.0625%) for Drosophila experiments.
Life span assay
According to Shen et al., 23 in this experiment, newly emerged Drosophila was divided into male and female under mild anesthesia with carbon dioxide gas. Then 200 males were randomly transferred to the control group and the experimental groups, with 10 tubes in each group and 20 Drosophila per tube. The food was replaced every 3 to 4 days, and the number of Drosophila deaths was recorded until all Drosophila were dead. Excel was used to calculate the maximum and average life span of each group of Drosophila, and the differences between groups were analyzed using GraphPad6 (version 6.01; GraphPad Software, La Jolla, CA, USA).
Climbing ability assay
According to method of Coulom and Birman, 24 in this experiment, three biological replicates were set up, and newly emerged Drosophila was randomly divided into four groups according to the above grouping method, with 20 flies per tube. The tubes were tapped until all flies reached the bottom. The number of Drosophila at the top and bottom was recorded after 10 s. The number on the top was subtracted from the number on the bottom, and the percentage was calculated. The tests were carried out at the 10th, 25th, and 40th day of culture.
Reproduction assay
According to Zou et al., 25 the reproductive capacity of Drosophila is also used as an indicator of aging. According to the above grouping method, newly emerged Drosophila was divided into male and female, and into four groups, and each group was set with five biological replicates. On day 10, 25, and 40 of culture, three pairs of Drosophila from the same group were combined into one vial containing the corresponding medium. The parent was removed after 5 days of incubation. Considering the emergence of the first Drosophila as the first day, the number of Drosophila that emerged within 7 days was counted.
Antioxidation assay
To investigate the mechanism of CAO in delaying the senescence of Drosophila, we analyzed its effects on the activity of antioxidant enzymes Cu,Zn-SOD and CAT and the content of MDA. Drosophila cultured on days 10, 25, and 40 was collected, starved for 2 h, and weighed. Drosophila homogenate was prepared in an ice bath at a ratio of 1:9 (weight of Drosophila [mg]: physiological saline [mL]). The supernatant was collected by centrifugation at 560 g/min for 15 min at 4°C. Detection of MDA, SOD typing, and CAT tests were performed according to the manufacturer's instructions.
Quantitative reverse transcription-polymerase chain reaction analysis
The optimal oligosaccharide dose was 0.125% based on the above experimental results, and flies in the basal diet control groups and 0.125% oligosaccharide test groups were cultured for the subsequent experiment. Drosophila was collected at the 10th and 40th days, ground in liquid nitrogen, and lysed by 1 mL TRIzol. Total RNA (8–10 per sample) was extracted from whole Drosophila, and cDNA was reverse transcribed. Ribosomal protein (rp49) was used as a reference gene and analyzed by high-throughput fluorescence quantitative PCR of tumor necrosis factor-α (TNF-α), nuclear factor kappa B (NF-κB), immune deficiency (IMD), and PIMS genes. Fold change was calculated using the 2–ΔΔCt method. The primer sequences and annealing temperatures are shown in Table 1 (primers were designed and synthesized by Wcgene Biotech, Shanghai, China).
Real-Time PCR Primers and Annealing Temperatures
16S rDNA sequencing
The same dose as the quantitative reverse transcription-polymerase chain reaction (RT-PCR) analysis, 0.125% dose diet, and basal diet groups were also selected for this test. The midgut of Drosophila was collected at the 10th and 40th days, corresponding to “adult” and “elderly.” The collected Drosophila was sterilized with 75% ethanol for 1 min and rinsed with 10% Tween phosphate-buffered saline (TPBS) for 1 min and was dissected in the midgut on ice. Then 16S rDNA assay (LC-Bio Technology Co., Ltd, Hang Zhou, China) was used to analyze the intestinal microbes. Briefly, DNA from different samples was extracted using the E.Z.N.A. ®Stool DNA Kit (D4015; Omega Bio-Tek, Inc., Norcross, GA, USA) according to the manufacturer's instructions. PCR amplification was performed by targeting the 16S rRNA gene sequence (region V6–V8), and libraries were prepared according to the guidelines provided by Illumina, provided by LC-Bio. The amplified 16S rDNA fragments were then sequenced using the Illumina MiSeq platform (version 1.8.0) with the Microbiome Helper workflow. Chimeric sequences were filtered using VSEARCH software (v. 2.3.4). Sequences with ≥97% similarity were assigned to the same operational taxonomic units (OTUs) by VSEARCH (v. 2.3.4). Representative sequences were chosen for each OTU, and taxonomic data were then assigned to each representative sequence using the Ribosomal Database Project (RDP) classifier. The differences in the dominant species in different groups and multiple sequence alignment were conducted using MAFFT software (v. 7.310) to study the phylogenetic relationships of different OTUs.
Statistical analysis
The expressions of TNF-α, NF-κB, PIMS, and IMD at the mRNA level were determined using Rp49 as an internal reference, and the relative expression level of mRNA was calculated using the 2–ΔΔCt method. The experiments were established in at least three biological replicate groups, and all data were analyzed by GraphPad Prism6 (version 6.01; GraphPad Software). Statistical significance was set at P < .05. Statistical analysis of bacterial composition and microbiota analysis were performed using R version 3.2.5.
Results
CAO prolonged the life span of Drosophila
We conducted survival tests only on male Drosophila, considering that estrogen has a great impact on the life span of female Drosophila. The results show that medium-dose and high-dose CAO supplementation may have an effect on life span extension (Fig. 1A). Therefore, we performed the analysis shown in Figure 1B. There was no significant difference between low-dose CAO groups and control groups in average life. In the middle- and high-dose groups, the average life extension rate was increased by 13.69% and 6.66%, respectively. Compared to the control group, the maximum life span in the middle- and high-dose groups was also significantly increased.

Life span extension of male Drosophila fed control and CAO-supplemented diet.
CAO increased the climbing activity of Drosophila
Drosophila fed a CAO-supplemented diet for 10 or 25 days did not exhibit significant differences in climbing activity compared to the control group. Interestingly, on the 40th day, Drosophila in the aging period showed that the diet with CAO significantly improved climbing activity compared to the control group. The vitality of the Drosophila in medium- and high-dose groups increased by 53.33% ± 9.428% and 40.00% ± 7.454%, respectively; P < .01.
CAO strengthened the reproduction of Drosophila
On day 10, there was no significant difference in the number of Drosophila that reproduced between the experimental and control groups. On day 25, the number that reproduced in the middle-dose supplementation group was significantly higher than that in the control group. It is worth noting that with senescence of Drosophila, the fertility rate was significantly higher in the middle- and high-dose supplementation groups than in the control group.
Effect of CAO on SOD activity, CAT activity, and MDA content in Drosophila
As shown in Figure 2A, at the 10th and 25th days, Cu,Zn-SOD activity in the low-dose CAO supplementation group increased by 24.01 ± 1.425 and 16.46 ± 2.034 U/mg protein, while that of the medium-dose group was significantly lower compared with control. At the 40th day, compared to the basal diet group, Cu,Zn-SOD activity in the CAO-supplemented groups was significantly increased. CAT activity with CAO supplementation was significantly lower than with basal diet at 10 days. Consistent with SOD activity, CAT activity in the CAO-dose groups was significantly higher than that in the control group, increasing by 16.51 ± 1.617, 11.82 ± 1.067, and 5.566 ± 0.8968 U/g protein, respectively (Fig. 2B). As shown in Figure 2C, compared to the control group, the MDA content increased by 9.483 ± 1.071 nmol/mg protein (P < .001) in 10-day-old male flies fed with the low-dose CAO-supplemented diet. In 25-day-old Drosophila with medium- and high-dose CAO-supplemented diets, the MDA content was significantly lower than in the basal diet. The MDA content in CAO-supplemented groups was significantly decreased compared with the control group at 40 days. It is worth noting that MDA content was consistent with Cu,Zn-SOD and CAT activity at the 40th day.

Antioxidant enzyme activity and MDA content in tissue of male Drosophila fed basal and CAO-supplemented diets on different days.
Effect of CAO supplementation on mRNA expression levels in male Drosophila
We measured the expression of representative genes that were associated with innate immune (TNF-α, NF-κB) and intestinal immune (IMD, PIMS) genes in each group at the 10th and 40th days. On the 10th day, comparing test groups with the control groups, immune-related genes (NF-κB, IMD) were significantly upregulated (Fig. 3A). It is worth noting that the expression of NF-κB and PIMS was significantly changed at the 40th day. NF-κB expression was downregulated 0.1081 ± 0.02299, and PIMS expression was upregulated 0.2955 ± 0.03728, P < .01 (Fig. 3B).

Gene expression levels of immune-related genes in tissue of male Drosophila fed basal or 0.125% CAO-supplemented diet on
CAO significantly improved microflora composition
We collected the midguts of Drosophila in each group at the 10th and 40th days, which were used for microbiota analysis. In Figure 4A, we can see a large distance between the basal diet and CAO diet groups; the number of species in the basal diet group was less than that in the CAO diet groups at the 10th and 40th days. Simpson index analysis also shows the distance between basal diet and CAO diet groups. Compared to the control group, community diversity was more abundant in CAO-supplemented groups at the 10th and 40th days (Fig. 4B).
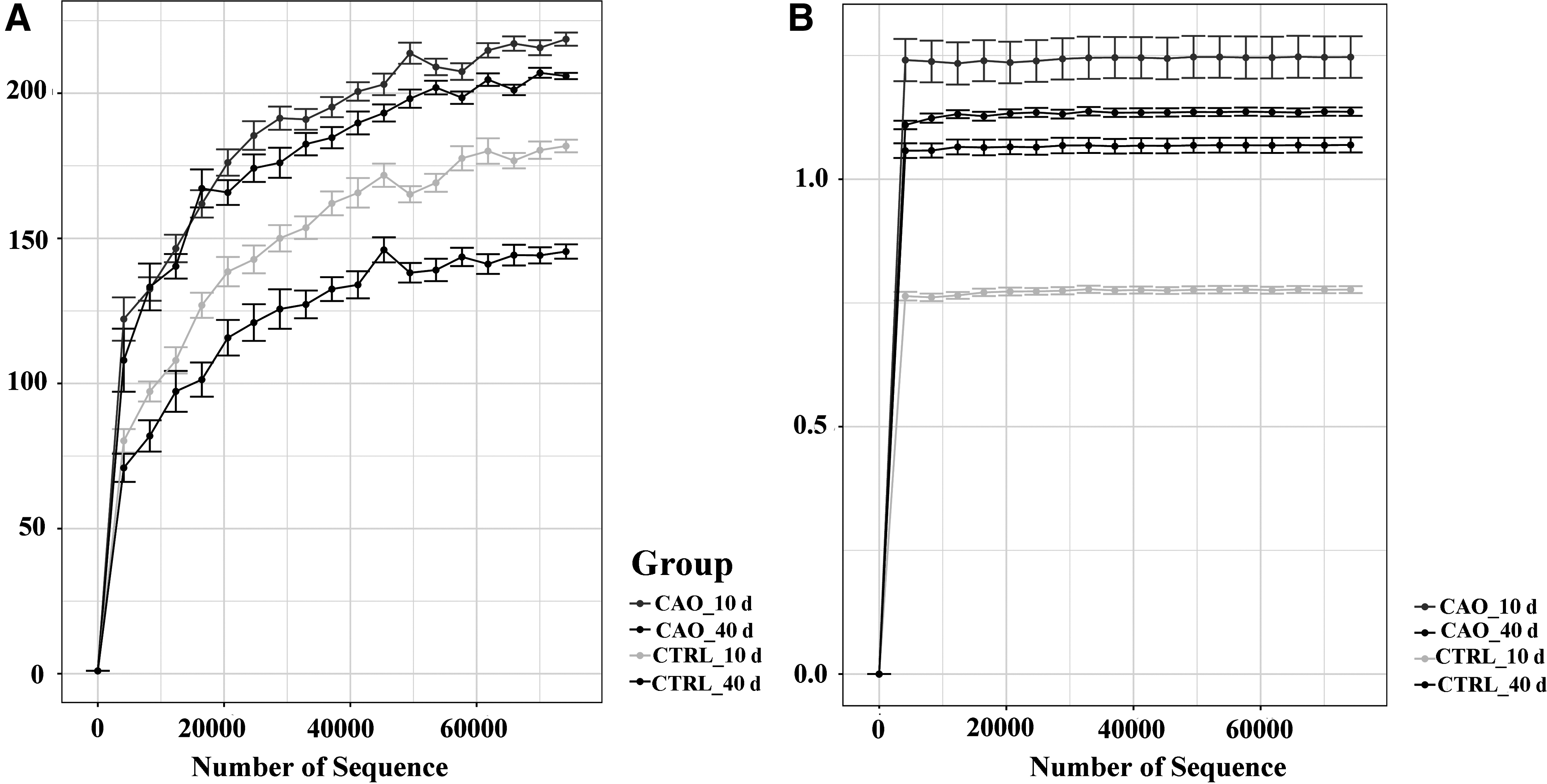
Response of alpha diversity of gut microbiota in basal and CAO-supplemented diet groups at 10th and 40th days.
CAO-supplemented diet changed the beta diversity in the midguts of Drosophila. The principal coordinates analysis (PCoA) scores separated the samples based on group (control vs. CAO supplemented) and age (10th vs. 40th day) along PCo1 and PCo2 (Fig. 5A). As shown in Figure 5B, 184 OTUs were common between CAO-supplemented and control groups at the 10th day, and 68 OTUs were unique to CAO and only 18 OTUs to basal diet. On the 40th day, 67 and 24 OTUs were unique to the CAO and basal diet, respectively.
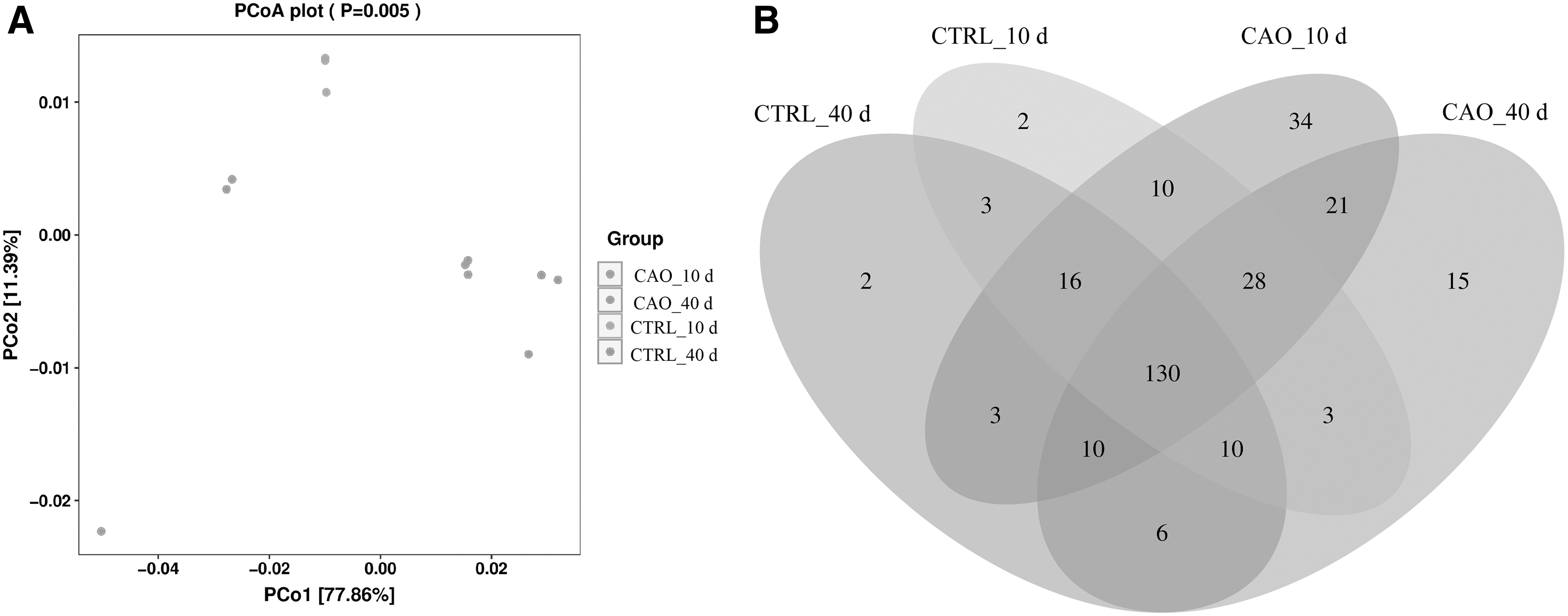
Differences in response of gut microbiota in each group at 10th and 40th day.
To further analyze the compositional changes in the midgut microbiota with CAO supplementation, the microbiota populations at the genus level were assigned by a stacked bar chart (Fig. 6A). At the 40th day, CAO changed the dominant microbiota populations. As shown in Figure 6B, C, compared to the control group, the abundance of Commensalibacter and Gluconobacter was changed significantly in the experimental groups. In the CAO-supplemented groups, the average ratio of Commensalibacter was lower than that in the control groups at 10 days. However, the abundance of Commensalibacter was increased significantly compared to the control group at the 40th study day. In contrast, a significantly lower abundance of Gluconobacter in the experimental groups than the control group was observed at the 40th day.
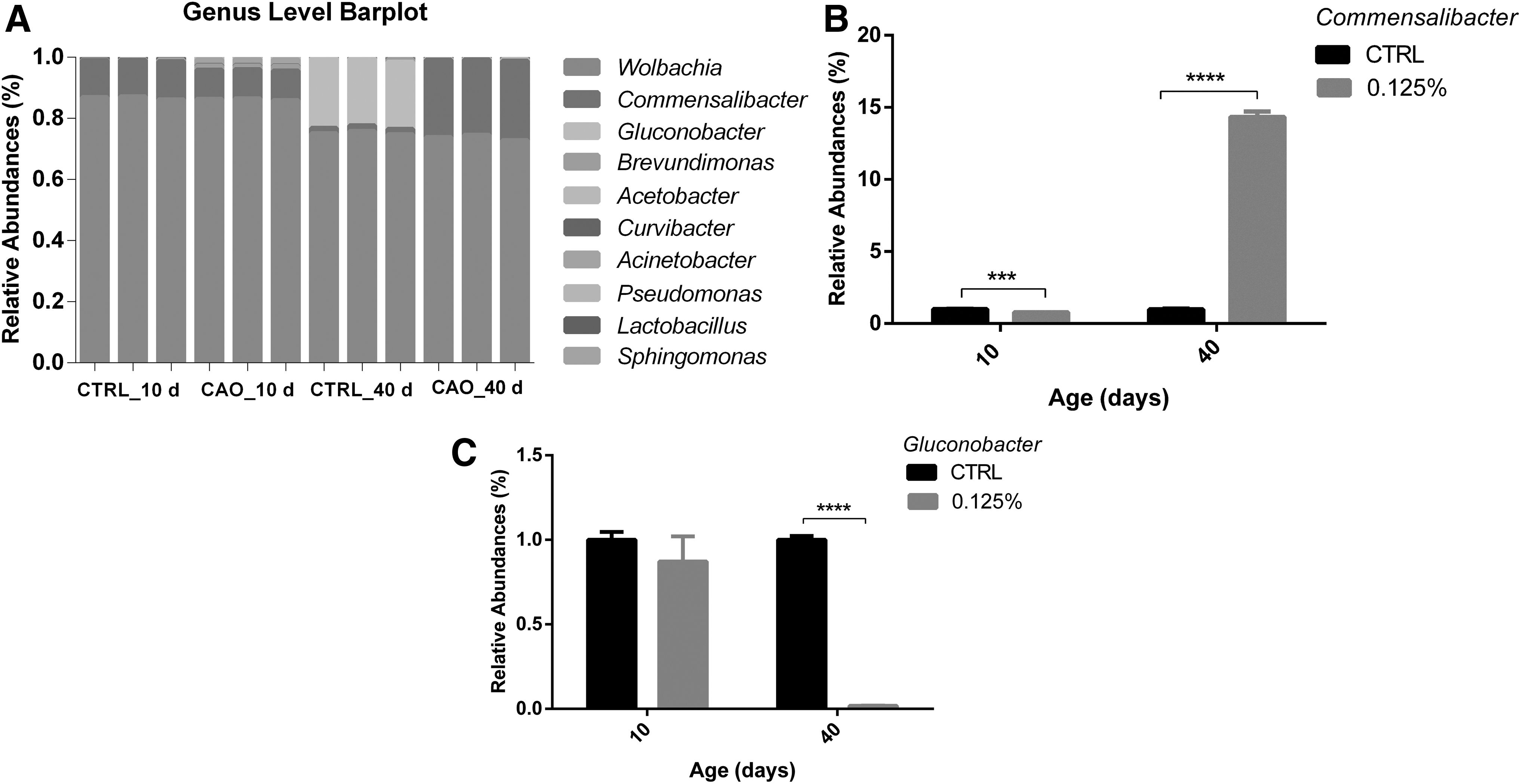
Relative bacterial abundance of different genera in fruit fly midgut microbiota in each group at 10th and 40th day.
Discussion
CAO is increasing in popularity as a supplement in food and medicine due to its various biological functions. 5,26 In this study, life span and reproduction were used to explore its antiaging effect in Drosophila, 23,27 and climbing activity as an indicator of health span was examined for the vitality of Drosophila. 28 The results show that a CAO-supplemented diet significantly prolonged the average and maximum life span of Drosophila. Furthermore, the climbing ability of old Drosophila was improved (−66.67 ± 13.33, −43.33 ± 8.82, and −56.67 ± 6.67 vs. −96.67 ± 3.33, Fig. 7), and the result of reproductive capacity was consistent with the vitality result, especially in the middle-dose group (6.40 ± 1.86, 31.20 ± 2.56, and 22.80 ± 3.51 vs. 3.00 ± 0.89, Fig. 8). Based on the above results, we found that CAO had a clear antiaging effect on the Drosophila.
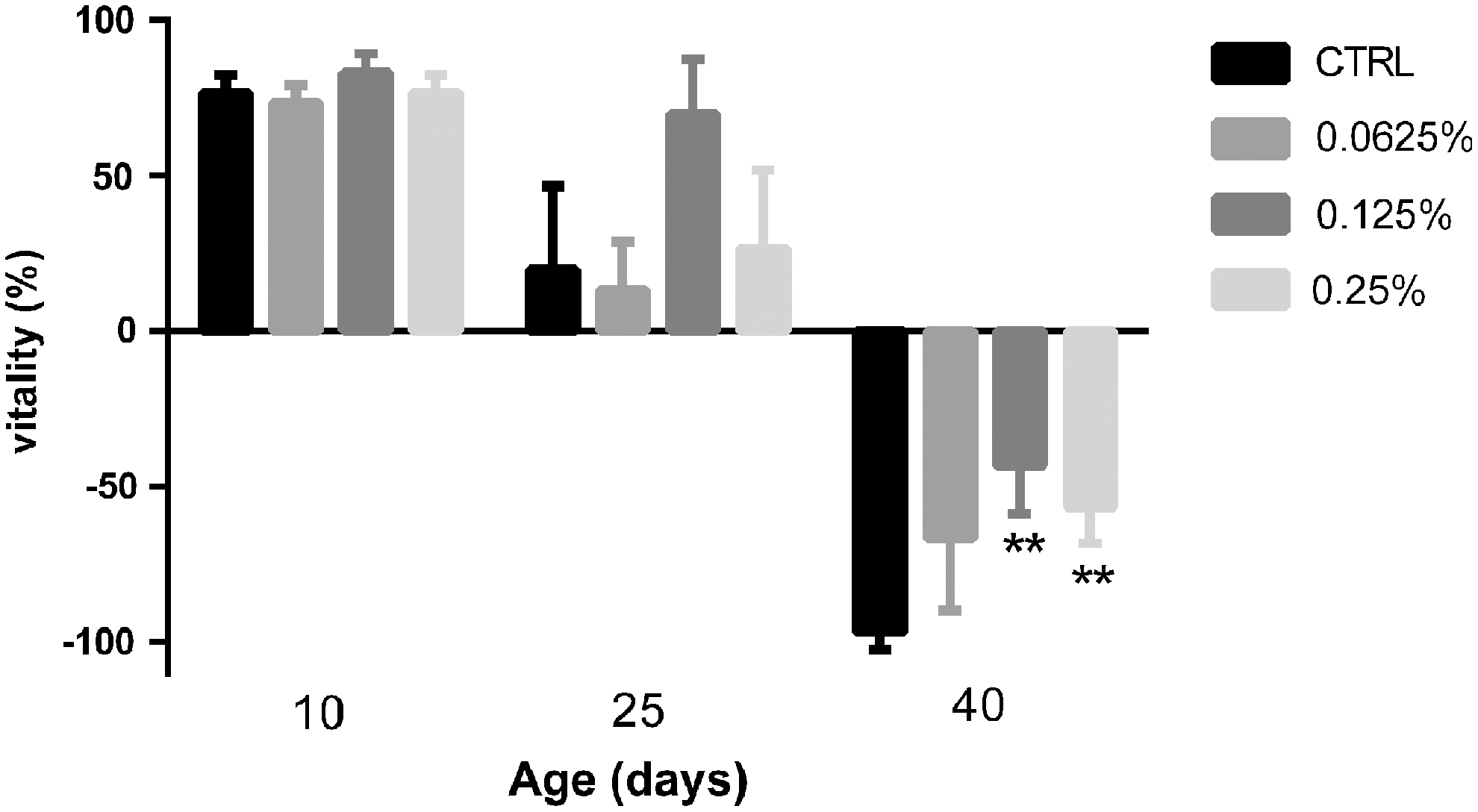
Climbing activity of male Drosophila fed control and CAO-supplemented diets on different days. Results expressed as mean ± SD (n = 3), and each group included three biological replicates; **P < .01 represents significant difference. SD, standard deviation.
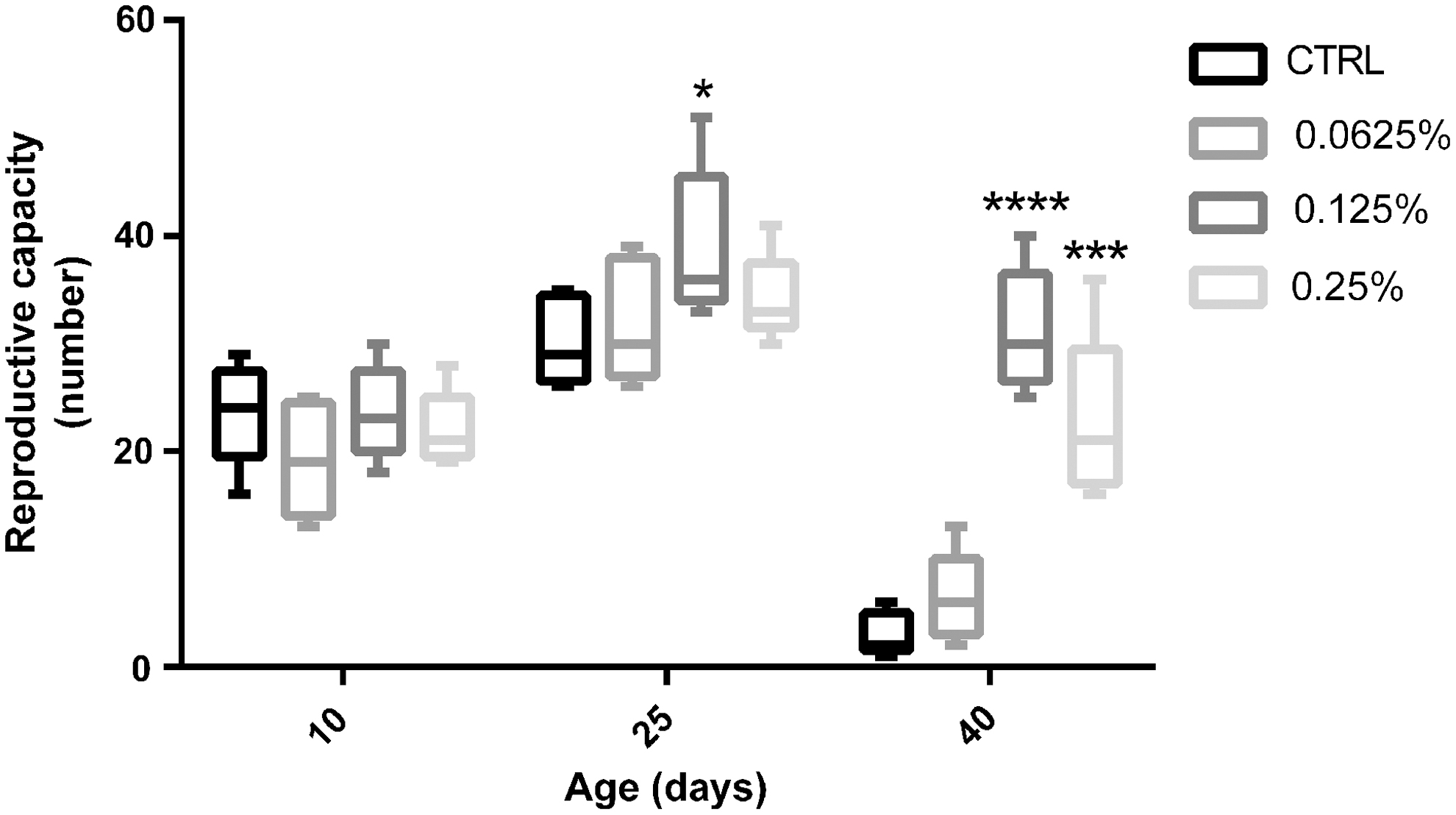
Reproduction capacity of male Drosophila fed control and CAO-supplemented diets on different days. Each group included five biological replicates; *P < .05, ***P < .001, and ****P < .0001 represent significant differences.
To explore the potential antiaging mechanism of CAO, we examined the antioxidant activity in vivo. The results indicate that the antioxidant effect of CAO on old Drosophila (40 days old) was significant, which is consistent with previous studies showing that CAO has antioxidant activity in vitro. 29 Drosophila's innate immunity is controlled by two major signaling pathways: the IMD and Toll pathways. Transcription of intestinal antimicrobial proteins (AMPs) is regulated by the TNF-like IMD signaling pathway. 30 TNF-α is a representative pro-inflammatory cytokine inducing inflammatory responses in the host, and NF-κB is a transcription factor regulating the expression of pro-inflammatory cytokines, including iNOS, COX-2, and prostaglandin E2 (PEG2). 31,32 PIMS is an inhibitor of the interaction of peptidoglycan recognition protein (PGRP-LC) and a negative regulator of IMD. Furthermore, PIMS is usually upregulated in the presence of commensal bacteria to suppress the immune response. 33 Our results indicate that the expression level of NF-κB and IMD was significantly upregulated by CAO at the 10th day. Interestingly, at the 40th day, the expression level of NF-κB was downregulated in male Drosophila, and the expression level of PIMS was upregulated, which was consistent with previous studies. We found that the immune-modulatory effect of CAO on old Drosophila was significant at the NF-κB gene level.
Previous research confirmed that many functional foods play a role in improving the composition of gut microbiota. Han et al. showed that alginate oligosaccharide (AlgO) can modify gut microbial composition. 34 At present, there are few reports on the effects of CAO on the intestinal microbes of Drosophila. In this study, the midgut microbiota was tested by 16S rDNA on 10- and 40-day-old Drosophila. As shown in Figure 4A, B, there were fewer species on the 40th day than the 10th day, while the alpha diversity was more abundant on the 40th day than the 10th day. The CAO-supplemented groups' alpha diversity was greatest on the 40th day. As shown in Figure 5A, there was a clear difference in intestinal microbiota between the CAO-supplemented and basal diets. Besides, the CAO-supplemented groups had more unique OTUs than the control group (Fig. 5B). The results indicate that the intestinal microbial composition was altered by CAO, and the alpha diversity was increased compared to the basal diet.
Considering the above results on antioxidant and innate immunity, we analyzed the intestinal microbial composition at the genus level to explore the potential antiaging mechanism of CAO. Obata et al. showed that the life span of Drosophila could be prolonged by microbiome remodeling. 35 In our study, the dominant microbiota at the genus level was changed using CAO supplementation. Our study shows that CAO significantly increased the abundance of Commensalibacter and decreased the abundance of Gluconobacter at the genus level at the 40th day (Fig. 6A, B). Commensalibacter was identified as a major component of the Drosophila gut microbiota. Commensalibacter and Gluconobacter genera belong to the Acetobacteraceae family. 36 Commensalibacter is a major component of the Drosophila gut microbiota, involved in innate immune regulation, and regulates commensal gut mutualism by suppressing the expression of NF-κB-dependent antimicrobial peptide genes. 36,37 Interestingly, the results were consistent with the RT-PCR results showing that the abundance of Commensalibacter was significantly increased, while the expression level of NF-κB was downregulated (Figs. 5B and 8B). This shows that CAO is involved in innate immune modulation by increasing the abundance of Commensalibacter and inhibiting the expression level of NF-κB.
In conclusion, our study demonstrated that CAO supplementation prolonged the life span of male Drosophila and improved the antioxidant activity of old male Drosophila. Furthermore, CAO maintained homeostatic immune modulation by inhibiting the expression of NF-κB and increasing the abundance of Commensalibacter of old male Drosophila. This provides a sufficient theoretical basis for the development and utilization of marine oligosaccharides as food additives or medical raw material that can be validated in future clinical trials in humans.
Footnotes
Authors' Contributions
Conceptualization, C.M. and X.J.-D.; methodology, X.J.-D. and C.M.; experimental work, Q.W.-L. and C.M.; analysis of the results, C.M.; writing—original draft, C.M.; and writing—review and editing, C.M. and X.J.-D.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This research was financially supported by a grant (2019C02064, 20191231Y150 and 2020R409032) from the Science and Technology Development Project of Zhejiang province, Hangzhou Science and Technology Development Project, China and Zhejiang Science and Technology Innovation Program for college students (ZhejiangXinmiaoTalents Program).