
Abstract
Idiopathic pulmonary fibrosis (IPF) is a chronic fibrosing interstitial lung disease with a poor prognosis similar to that of malignancy. The causes of IPF are not clearly known, and there is no effective therapy to date. In this study, the natural compound plumbagin, which was isolated from Plumbago rosea root extract, was screened for p300 inhibitory activity. Plumbagin specifically inhibited the activity of p300 toward histone acetyltransferases. Plumbagin treatment significantly suppressed transforming growth factor-β-induced profibrotic target-gene expression and proliferation of fibroblast cell lines. Moreover, plumbagin significantly inhibited bleomycin-induced pulmonary fibrosis in mice. Taken together, these data demonstrate the inhibitory effects of plumbagin on lung fibrosis and its promise as a therapeutic agent for IPF.
Introduction
Idiopathic pulmonary fibrosis (IPF) is a serious interstitial lung disease with a poor prognosis. 1 To date, the causes of IPF have not been clearly established, but excessive activation of fibroblasts, increased expression of extracellular matrix (ECM) proteins, damage to epithelial cells, and genetic causes have been reported. 2 There is currently no drug that can effectively treat IPF. The antifibrosis drug pirfenidone and the immunosuppressive agent azathioprine are typically used for treatment of IPF patients, but the effect is not significant. 3 Thus, it is urgent to develop effective and specific therapeutic agents.
Regulation of gene expression by epigenetic influences has recently been found to be an important mechanism of phenotypic changes in cells, tissues, and organs through environmental or other stimuli such as tobacco smoke, air pollution, and aging. 4 Histone modifications in epigenetics include histone acetylation, methylation, and phosphorylation, and histone acetyltransferases (HATs) are highly associated with transcriptional regulation of target genes. 5 Among them, p300 is the most widely studied HAT protein and is involved in transcriptional activation of various genes in response to cellular signaling such as inflammation, growth, and nuclear hormones. 6 Expression of p300 is increased in systemic sclerosis, and hyperacetylated H4 on the collagen promoter induces transcription with Smad activation. 7 Notably, p300 mediates transforming growth factor (TGF)-β-induced transcriptional activation of fibrotic genes during pulmonary fibrosis. 8 Recently, it has been shown that expression of active p300 is increased in IPF patient-derived fibroblasts, which causes inactivation of the HDAC1 protein and eventually leads to pulmonary fibrosis. 9 In addition, inhibition of p300 and discoidin domain receptor 1 (DDR1) synergistically reduces pulmonary fibrosis in an in vivo mouse model. 10 Collectively, these studies suggest that p300 is a promising target for treatment of IPF. Therefore, further research is needed to develop IPF therapeutic agents using p300 inhibition.
Plumbagin (5-hydroxy-2-methyl-1,4 naphthoquinone) is a quinoid component extracted from the roots of herbs of the Plumbago genus and has various pharmacological effects. 11 Plumbagin has shown potential therapeutic benefits for diverse diseases such as melanoma, breast cancer, colon, nonsmall cell lung cancer, and liver fibrosis, 12,13 and a previous report showed an inhibitory effect of plumbagin on p300 HAT activity. 14 However, the role of plumbagin in pulmonary fibrosis by inhibiting p300 has not yet been demonstrated.
In this study, single substances derived from natural compounds were assessed for p300-inhibitory activity. Among these compounds, plumbagin had effective and selective p300-inhibitory effects on HAT activity. Plumbagin inhibited fibroblast proliferation and fibrotic target-gene expression. In addition, the therapeutic effects of plumbagin were verified in a bleomycin-induced lung fibrosis mouse model. Taken together, these data demonstrate that the selective p300 inhibitor plumbagin suppresses lung fibrosis and is a potential therapeutic agent for IPF treatment.
Materials and Methods
Cell culture and reagents
The human lung fibroblast cell line MRC5 and the mouse lung fibroblast cell line Mlg were purchased from Korean Cell Line Bank (Seoul, Korea). Cells were cultured in Dulbecco's modified Eagle's medium supplemented with 10% (v/v) fetal bovine serum and 1% antibiotic/antimycotic solution (Corning, Manassas, VA, USA) at 37°C under 5% CO2. Plumbagin, lawsone, juglone, geraniol, sennoside A, allyl isothiocyanate, and phenethyl isothiocyanate were purchased from Sigma-Aldrich (St. Louis, MO, USA). Taraxerol, prunetin, and sakuranetin were purchased from ChemFaces (Hubei, China), and TGF-β1 was purchased from ProSpec (East Brunswick, NJ, USA).
In vitro HAT assay
HAT assay was carried out using the HAT Activity Fluorometric Assay Kit (BioVision, Milpitas, CA, USA) according to the manufacturer's instructions. Two hundred nanograms of GCN5, p300 (Enzo Life Science, Farmingdale, NJ, USA), and PCAF (Sigma-Aldrich) recombinant proteins were used as enzyme sources and Histone-H3 peptide was used as a substrate. Enzyme reaction monitored by measuring fluorescence (Ex/Em = 535/587 nm) using a Varioskan Flash 3001 reader (Thermo Fisher Scientific, Vantaa, Finland) in kinetic mode for 60 min at 25°C.
RNA isolation and quantitative reverse transcription-polymerase chain reaction (qRT-PCR)
Total RNA from cells was prepared using Ribospin II (GeneAll Biotech, Seoul, Korea), and cDNA was synthesized using CellScript (CellSafe, Yongin, Korea), both following to the manufacturer's protocol. The following PCR primer sequences for humans were used for qRT-PCR: collagen 1a1 (Col1a1) 5′-CCTCAAGGGCTCCAAC-3′ and 5′-GGTTTTGTATTCAATCACTGTCTTGC-3′, collagen 3a1 (Col3a1) 5′-TGGTCTGCAAGGAATGCCTGGA-′ and 5′-TCTTTCCCTGGGACACCATCAG-3′, alpha-smooth muscle actin (α-SMA) 5′-CTGGCATCGTGCTGGACTCT-3′ and 5′-GATCTCGGCCAGCCAGATC-3′, fibronectin (FN) 5′-GAGCTATTCCCTGCACCTGATG-3′ and 5′-CGTGCAAGGCAACCACACT-3′, Snail 5′-CCCCAATCGGAAGCCTAACT-3′ and 5′-GGACAGAGTCCCAGATGAGC-3′, plasminogen activator inhibitor-1 (PAI-1) 5′-CCCCACTTCTTCAGGCTGTT-3′ and 5′-GCCGTTGAAGTAGAGGGCAT-3′, β-actin 5′-TGCGTGACATTAAGGAGAAG-3′ and 5′-GTCAGGCAGCTCGTAGCTCT-3′. The cDNA concentration was normalized using human β-actin. Quantitative RT-PCR analysis was performed using SYBR Green PCR Master Mix reagents and an ABI Prism 7700 Sequence Detection System (Applied Biosystems, Carlsbad, CA, USA).
MTT assay
Cell viability was measured using the MTT assay. Fibroblast cells were seeded in a plate and incubated overnight. The cells were then starved of serum with the indicated concentration of plumbagin for 2 h, followed by TGF-β1 treatment for 24 h. Fifteen micromolar concentration of MTT solution (Sigma-Aldrich) was added to cells and incubated for 90 min at 37°C, and then incubated with dimethyl sulfoxide for 30 min with shaking at room temperature to dissolve the formation of formazan. The absorbance was measured at 570 nm using a microplate reader with a reference at 630 nm.
Animal studies
Animal experiment was approved the study protocol by the Institutional Animal Care and Use Committee of Yonsei University College of Medicine (Certification No. IACUC-2018-0087). Eight-week-old male mice were intratracheally injected with saline (vehicle control) or 4 mg/kg bleomycin (Santa Cruz Biotechnology, Santa Cruz, CA, USA). Five mice were used for each group. The next day, the mice were injected with plumbagin (1 or 2 mg/kg). The dose of the plumbagin injection was determined by referencing previous studies. 15,16 The mice were then sacrificed 14 days after receiving bleomycin, and bronchoalveolar lavage (BAL) fluid and lung tissues were harvested. Mouse lung tissues from each group were fixed in formaldehyde (10% v/v) for 48 h at room temperature and embedded in paraffin.
Masson's trichrome staining
Paraffin-embedded lung tissue sections were fixed in Bouin's solution (Sigma-Aldrich), stained with Weigert's Iron Hematoxylin Set (Sigma-Aldrich) and Masson's Trichrome Staining Kit (Sigma-Aldrich) according to the manufacturer's manual. Blue stained collagen fiber quantification was analyzed using ImageJ software in × 200 field image area (Olympus, Tokyo, Japan).
Soluble collagen assay
Collagen content in the mouse lung tissues was determined biochemically using the Sircol Collagen Assay Kit (Biocolor, Carrickfergus, United Kingdom) according to the manufacturer's protocol.
Statistical analysis
All results were analyzed with Prism software, version 5 (GraphPad Software) and are shown as the mean ± standard error of the mean. Student's t-test used for determining statistical significance of two groups. One-way analysis of variance was used when comparing samples from two groups for repeating parameters, or for multiple comparisons. The significance levels are indicated as follows: not significant, P > .5; *P ≤ .05; **P ≤ .01; and ***P ≤ .001.
Results
Plumbagin specifically inhibits p300 HAT activity
We previously reported that various plant extracts possess anti-HAT activities; however, it was not known which specific compound or compounds were responsible. 17 –19 To identify the effective p300 HAT inhibitors among those natural substances, we selected the 10 primary single compounds and measured the p300 HAT activity. Plumbagin is known to have anticancer effects and to regulate histone acetylation in previous studies. 14,20 Lawsone and juglone, which are isoform compounds of plumbagin, have anticancer and antibacterial and regulation of autophagy signal. 21,22 Also, Allyl- and Phenethyl isothiocyanate are sulfur-containing compounds and known to undergo epigenetic regulation. 23,24 Sennoside A, 25 Taraxerol, 26 Geraniol, 27 Sakuranetin, 28 and Prunetin 29 are known to be highly effective in regulating NF-κB pathway, anticancer, and anti-inflammation. Among these, plumbagin displayed the most significant inhibitory effects against the p300 enzyme (80% inhibition of p300 activity at 1 μM concentration) (Fig. 1A). Ravindra et al., established that the hydroxyl group of plumbagin is required for inhibition of p300 HAT activity. 14 Our data also show an inhibitory effect of plumbagin on p300 HAT activity compared to other natural compounds. Next, we tested the dose–response of plumbagin on inhibition of p300 HAT activity. As shown in Figure 1B, p300 HAT activity was inhibited by plumbagin in a dose-dependent manner. There was a 90% inhibitory effect on p300 HAT activity with 2.5 μM plumbagin. To examine the selective effects of plumbagin on p300 HAT activity compared to other HATs, the assay was performed in the presence of the recombinant proteins p300, GCN5 (KAT2A), or PCAF (P300/CBP-associated factor, KAT2B) with or without plumbagin. Plumbagin selectively inhibited p300 HAT activity but not GCN5 or PCAF (Fig. 1C). These results demonstrate that plumbagin selectively inhibits p300 HAT activity.
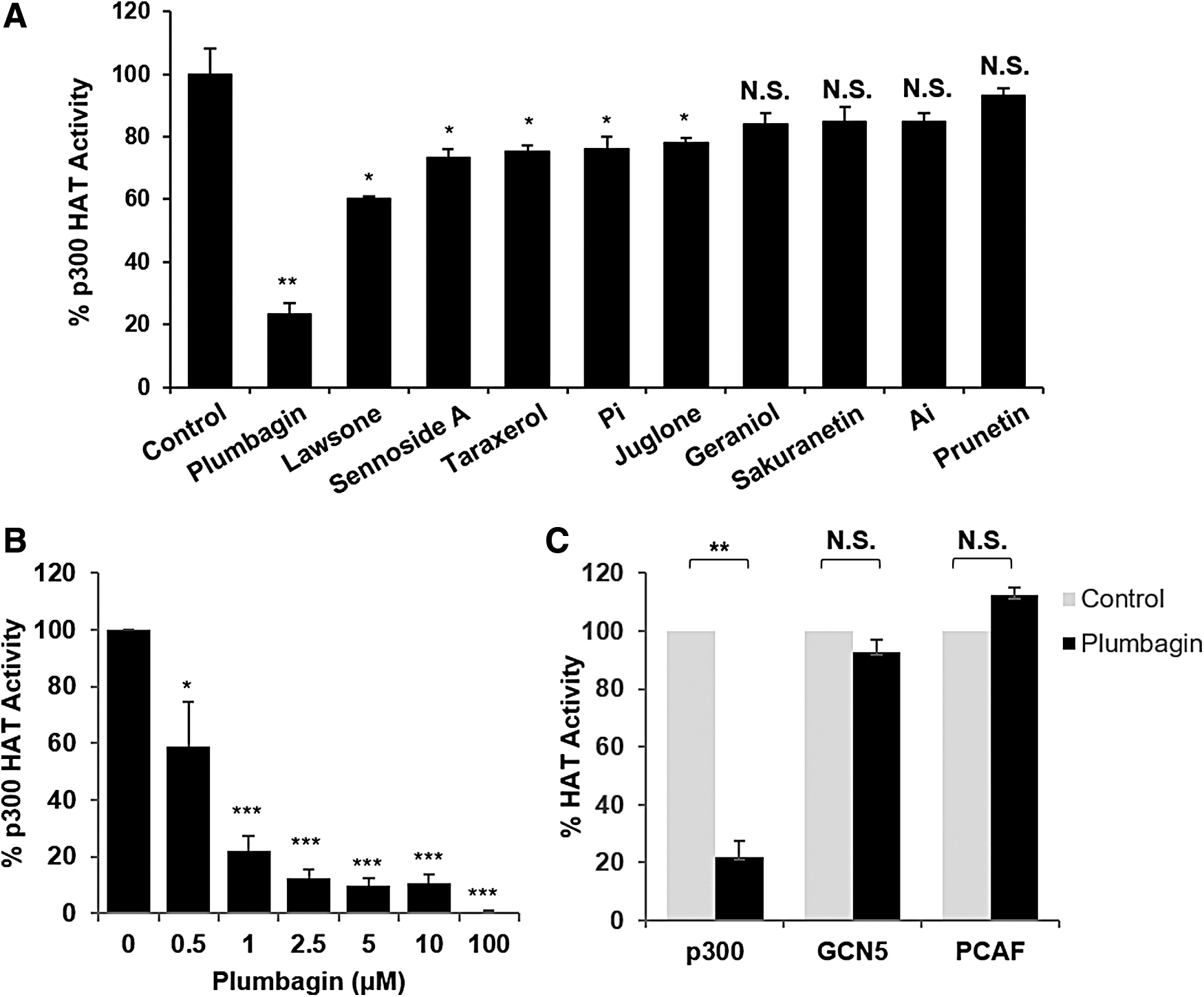
Inhibitory effects of plumbagin on p300 HAT activity.
Inhibition of p300 by plumbagin suppresses fibroblast proliferation
To evaluate the cytotoxic effects of plumbagin, the human and mouse fibroblast cell lines MRC5 and Mlg, respectively, were treated with increasing doses of plumbagin for 24 h. The MTT assay revealed that cell viability was unchanged at plumbagin concentrations lower than 10 μM in MRC5 cells and 5 μM in Mlg cells, but higher doses of plumbagin had cytotoxic effects (Fig. 2A). TGF-β is a major mediator of fibrosis in many tissues, and fibroblast cells can be activated and proliferated with TGF-β treatment. 30 To examine the inhibitory effects of plumbagin on TGF-β-induced proliferation, we assessed cell viability in TGF-β-treated fibroblast cells lines with or without plumbagin. As shown in Figure 2B, TGF-β treatment increased fibroblast proliferation; however, plumbagin dose-dependently inhibited proliferation of both MRC5 and Mlg fibroblast cells. Taken together, these data show that plumbagin inhibits TGF-β-induced fibroblast proliferation.
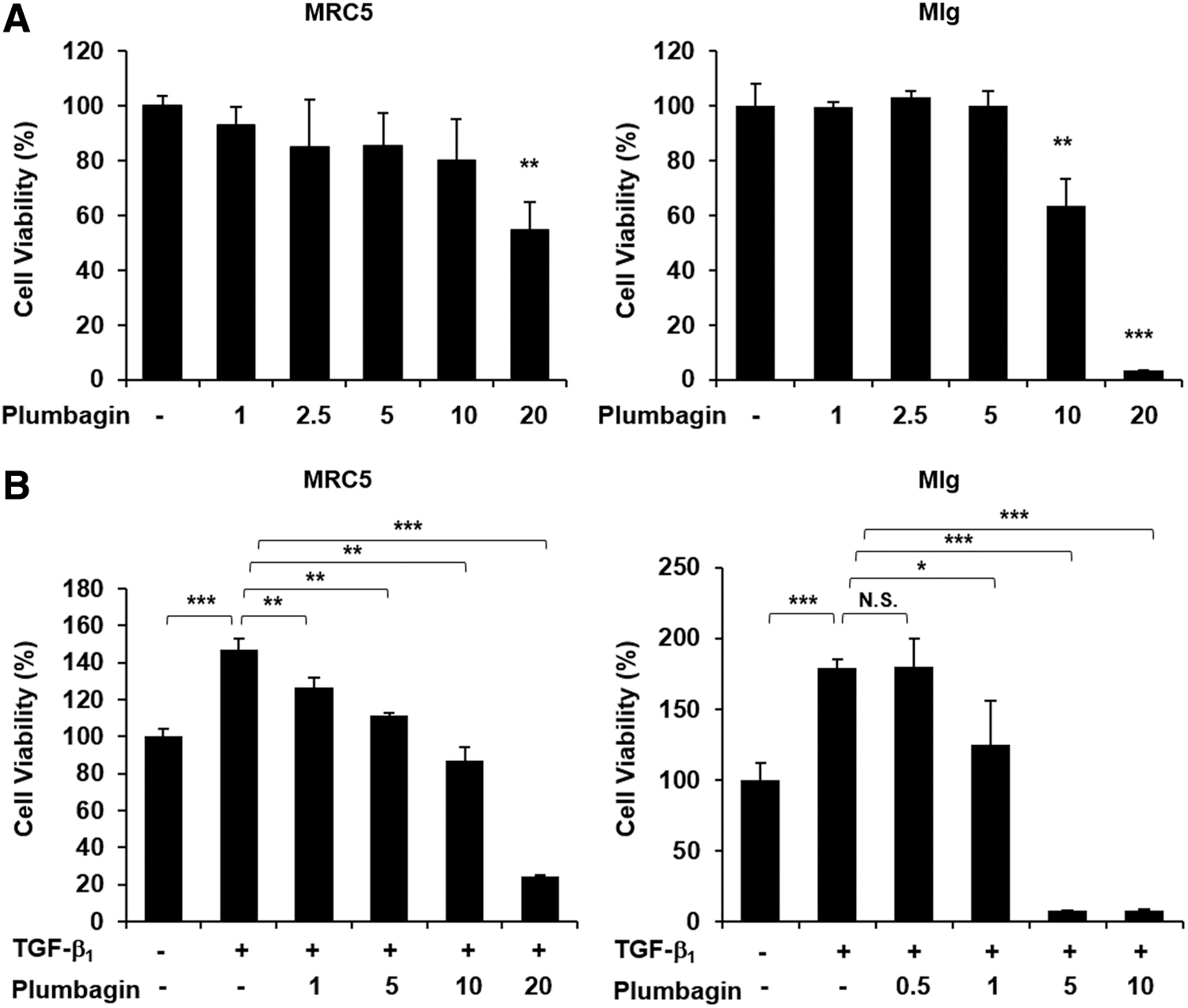
Cytotoxicity of plumbagin in fibroblast cells.
Plumbagin reduces TGF-β-induced target-gene expression in fibroblasts
Given the data suggesting that plumbagin has inhibitory effects on p300 HAT activity and TGF-β-induced fibroblast proliferation, we next tested whether plumbagin regulates fibrotic target-gene expression in fibroblasts. As expected, TGF-β treatment significantly increased the expression of fibrotic target genes, such as Col1a1, Col3a1, α-SMA, FN, snail, and PAI-1. However, plumbagin substantially suppressed TGF-β-induced upregulation of various target genes (Fig. 3). These results indicate that plumbagin inhibits TGF-β-induced fibroblast activation by regulating fibrotic target-gene expression.
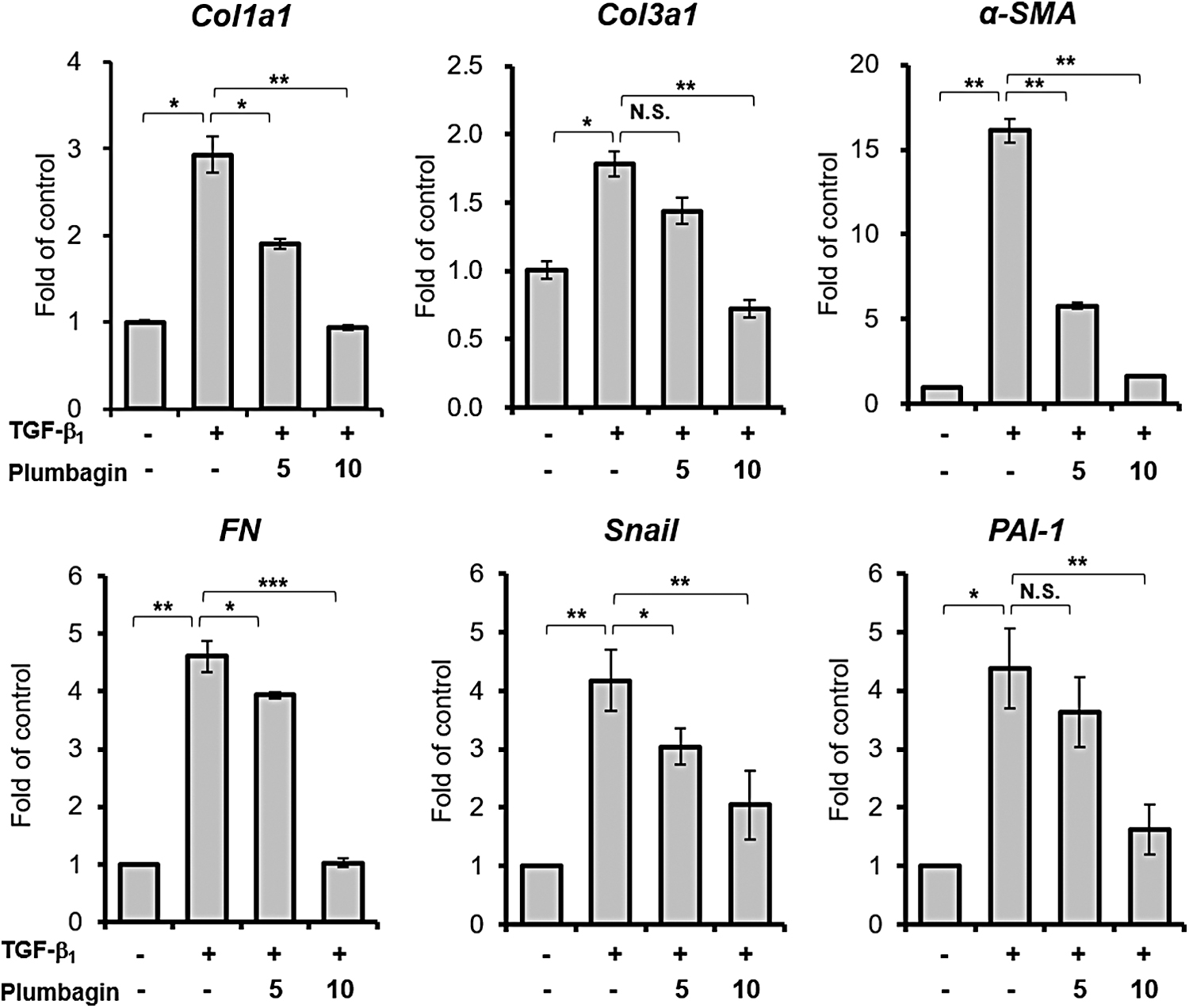
The effects of plumbagin on profibrotic gene expression in response to TGF-β. MRC5 cells were incubated without serum in the presence of the indicated dose of plumbagin (μM) along with 20 ng/mL of TGF-β. After 24 h of TGF-β treatment, the expression of indicated genes was assessed using quantitative reverse transcription-polymerase chain reaction. Results represent the mean ± SEM (n = 3); *P < .05; **P < .01; ***P < .001.
Inhibitory effects of plumbagin in a bleomycin-induced lung fibrosis mouse model
Bleomycin has been used as an experimental pulmonary fibrosis model in mice, and increases TGF-β level in the lung. 31 Therefore, we validated the antifibrotic action of plumbagin in this mouse lung fibrosis model. Numerous fibrotic changes were observed at 14 days after bleomycin intratracheal injection. Notably, bleomycin-induced lung fibrosis was dramatically inhibited following treatment with 1 or 2 mg/kg of plumbagin (Fig. 4A), and these Masson's trichrome staining (MTS) regions were quantified using ImageJ software (Fig. 4B). The Sircol Collagen Assay also showed that the bleomycin-induced fibrotic regions of the mouse lungs were significantly reduced following plumbagin treatment (Fig. 4C). Furthermore, cell counts of BAL fluid showed that bleomycin-induced inflammatory cells were significantly decreased after plumbagin injection (Fig. 4D). Collectively, these data demonstrate that plumbagin prevents pulmonary fibrosis by inhibiting ECM production.

Antifibrotic action of plumbagin in a mouse lung fibrosis model.
Discussion
Phytochemicals from plants play an important role both in traditional and modern medicinal systems. Plumbagin is a yellow crystalline phytochemical derived from naphthoquinone that has a broad spectrum of biological effects, including anti-inflammatory, anti-angiogenesis, anti-cancer, and antioxidant properties. 32 Recently, several studies have suggested an ameliorative effect of plumbagin in liver and kidney fibrosis. 33,34 Wang et al. investigated the liver-protective role of plumbagin in thioacetamine-induced liver fibrosis and found that plumbagin treatment promoted AMPK phosphorylation, which inhibited p300/SMAD complex formation and reduced α-SMA and collagen 1/3 expression in hepatic stellate cells. 35 However, the role of plumbagin in lung fibrosis has not yet been studied.
IPF is a type of lung disease that causes fibrosis of the lung for unknown reasons. 2 An important event in IPF is migration of fibroblasts into the alveolar space and abnormal proliferation after lung injury. Under normal conditions, fibroblasts play a critical role in wound healing after damage as well as in connective tissue production. However, the function of fibroblasts in the fibrotic lung is uncontrolled, and the resulting fibroblastic foci consist of active proliferative fibroblasts, immune cells, and excessive ECM protein deposition. These changes promote alterations of lung structure and function, including loss of gas exchange and decreased oxygen saturation of the blood. 36,37 The prognosis for IPF patients is typically poor, and there is an ongoing search for drugs that inhibit the pathogenic pathways in IPF to reduce or delay progression of the disease. 38 A recent study showed that p300 inhibition decreased the characteristics of fibrosis in primary fibroblasts from IPF patients, in an in vivo bleomycin mouse model, and in ex vivo precision-cut lung slices, suggesting that p300 inhibition is an efficient therapeutic target for IPF treatment. 9
HAT p300 plays an important role in fibrosis and regulates the fibrotic response by regulating homeostasis of the ECM, myofibroblast activation, and epithelial-mesenchymal transition. 39 The HAT activity of p300 and its interaction with activated Smads are essential for TGF-β-induced profibrotic signaling. 7 Moreover, p300 expression is limited in normal fibroblasts, and ectopic expression of p300 is notably enhanced in TGF-β response, demonstrating that p300 might have a critical role in the progression of tissue fibrosis. 40 –42 Small molecule modulators of HATs have received attention recently since they possess huge potential as molecules for therapy. Plumbagin has been reported to be a potent HAT inhibitor of p300 through hydrogen bonding of fifth hydroxyl group to a critical lysine residue within the HAT domain of p300. 14 In the current study, we verified that plumbagin has potent anti-p300 HAT activity compared to similarly structured compound such as juglone (5-hydroxy-2-methyl-1,4 naphthoquinone) and lawsone (2-hydroxy-1,4-naphthoquinone) (Fig. 1A). Moreover, plumbagin displayed selective anti-HAT activity against p300 but not against GCN5 and PCAF, indicating that plumbagin is a selective inhibitor against p300. Importantly, we also showed that plumbagin prevents bleomycin-induced lung fibrosis, which is in accordance with previous results showing that it inhibits liver and kidney fibrosis. 33,34 However, it is unknown whether the antifibrotic action of plumbagin is due to its anti-p300 HAT activity. Therefore, further study is needed to address whether the anti-p300 HAT activity of plumbagin is required for its antifibrotic action.
In conclusion, our study demonstrates the therapeutic effects of plumbagin on pulmonary fibrosis through inhibition of p300 activity. Plumbagin reduces fibrotic target-gene expression and proliferation in a response to TGF-β in fibroblasts and prevents bleomycin-induced lung fibrosis. Therefore, our data suggest that selective inhibition of p300 by HAT inhibitor is a possible therapeutic strategy for treatment of IPF.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This research was supported by a grant of the Korea Health Technology R&D Project through the Korea Health Industry Development Institute funded by the Ministry of Health and Welfare, Republic of Korea (HI17C0104)