
Abstract
Research on the beneficial effects of Maillard reaction products (MRPs) and phenolic compounds derived from roasted peanut flour on the nervous system remains insufficient. This study aimed to evaluate the effect of a 28-day oral administration of defatted peanut extract rich in MPRs and polyphenolic compounds on the cognitive impairments and oxidative injury induced by scopolamine in a mouse model. Light and dark extracts from peanut flour were prepared by heating peanuts at 187°C for two different times (8.6 and 12.7 min) and defatted using soxhlet apparatus. The mice were orally pretreated with either roasted defatted peanuts extracts (100 mg/kg) or donepezil (3 mg/kg) for 21 days. On day 19 and until day 28, mice were injected subcutaneously with water or scopolamine (1 mg/kg body weight) 15 min after roasted defatted peanuts extracts/water feeding. Mice were subsequently subjected to a battery of behavioral tests including open field locomotor activity assay, and Morris water maze test. Brain tissues were collected to measure acetylcholine, acetylcholinesterase, and oxidative parameters (glutathione and malondialdehyde). Roasted defatted peanuts (light and dark) (100 mg/kg) treatment significantly ameliorated cognitive performance and reversed the oxidative damage when compared with the scopolamine group. These data demonstrate the defatted peanuts extracts exert potent anti-amnesic effects via the modulation of cholinergic and antioxidant activities.
Introduction
Cumulative oxidative stress can lead to cell and tissue damage, which results in age-related neurodegenerative diseases, such as Parkinson's disease and Alzheimer's disease (AD). It is estimated that there are ∼35 million people worldwide living with AD. 1 This number is expected to grow by an average of 2.3 million people per year until 2030 because of the increasing life expectancy and population demographics. AD with high prevalence in older population manifests two notable symptoms, loss of memory and behavior changes. 2 A large body of evidence suggests that oxidative stress resulting from reactive oxygen species (ROS) and reactive nitrogen species affects glial cells and neurons from patients with AD and therefore, leading to neuronal damage and decrease of central acetylcholine (ACh) levels. 3,4 Moreover, antioxidants defense systems, such as superoxide dismutase, glutathione peroxidase, and catalase, scavenge surplus ROS and protect brain cells from oxidative injury. 4 Therefore, the use of combinations of antioxidants with acetylcholinesterase (AChE) inhibitors can protect neurons against oxidative conditions and prevent the destruction of ACh.
Recent years have seen increased consumption of tree nuts and peanuts in general. Research suggests that this increased intake of tree nuts and peanuts may be associated with a reduced incidence of disorders induced by ROS, including cardiovascular disorders, cancer, and neurodegenerative diseases. 5,6 Nuts (tree nuts and peanuts) are nutrient-dense foods that are rich in unsaturated fatty acids, fiber, vitamins, minerals, and many other bioactive substances, such as phenolic antioxidants and phytosterols. 7 These natural antioxidants play a vital role in the prevention or delaying the course of these diseases. 8 However, the composition and the bioaccessibility of these components are largely affected by nuts roasting. Indeed, during roasting, some of the compounds are lost, and the Maillard reaction products (MRPs) appear, thus affecting the product's sensory and antioxidant properties. 9 The Maillard reaction (MR) is a complex reaction between reducing sugars and amino acids and is responsible for the brown color. 10 The class of MRPs that appears during food processing is defined by heating duration and temperatures. These neoformed compounds might be one of the important factors for either disease progression or combating disease. Research on the beneficial effects of MRPs on the nervous system remains insufficient. The objective of this study is to determine the effect of a 28-day oral administration of defatted peanut rich in MPRs and phenolic compounds on the cognitive performance of adult mice as assessed by Morris water maze (MWM) task, brain oxidative status, and AChE determination.
Materials and Methods
Chemicals
Chemicals used were of analytical grade. Scopolamine hydrobromide, ACh chloride, ACh assay kit, donepezil hydrochloride monohydrate, potassium ferricyanide, ferrozine, trichloroacetic acid, 5,5-dithiobis (2-nitrobenzoic acid), and 2, 4-dinitrophenyl hydrazine were obtained from Sigma Chemical Company (St. Louis, MO, USA).
Sample preparation
Raw peanut kernels (Arachis hypogaea) were purchased from grocery stores and the kernels with skin (1 kg) were roasted in an electric oven with the roasting temperature of 187°C for 0, 8.6, and 12.7 min according to McDaniel et al. 11 After cooling, the skin was removed, and 20 g of each kernel were recovered, ground, and defatted with 200 mL of acetone for 24 h using a Soxhlet apparatus to obtain peanut flour. Finally, the defatted samples were dried at 35°C to remove residual solvent and preserved at −20°C. For the extraction, 5 g of peanut flour were extracted with ethanol-water in the ratio of (1:1) in an ultrasonic water bath at 60°C for 30 min. The extract was filtered, and the residue was re-extracted two times, with ethanol–water at the same conditions. Finally, the samples were filtered, concentrated using a rotavapor, and lyophilized. Samples are named “RPE” for raw peanuts extract without roasting; “LPE” for light peanuts extract roasting temperature at 187°C for 8.6 min, and “DPE” for dark peanuts extract roasting temperature at 187°C for 12.7 min.
Color measurement
The sample color appearance was measured using a colorimeter (Spectraflash 600 plus; Data-Color International, Lawrenceville, New Jersey, USA). The International Commission on Illumination (CIE) color values recorded were as follows: L* = lightness (0 = black, 100 = white); a* (−a* = greenness, +a* = redness); and b* (−b* = blueness, +b* = yellowness). Color differences (ΔE) were calculated by the following equation [Eq. (1)]:
Three readings were recorded for each sample and the average values were calculated.
Browning and fluorescence intensity
The extent of the MR on roasted and defatted peanuts extract was measured at 294 and 490 nm using an ultraviolet-visible spectrophotometer (Model 6305; Jenway, Staffordshire, UK) according to the method of Rufián-Henares and Morales. 12 Fluorescence intensity (FI) of extracts was measured using a fluorescence spectrophotometer (Versa Fluor Spectro-Fluorometer; Bio-Rad, Hercules, CA, USA), with the excitation wavelength at the 370 nm and the emission wavelength at the 450 nm 13
Total polyphenol content determination
The total phenolic content (TPC) of the extract was determined according to the Górnaś et al. method using the Folin–Ciocalteu's phenol reagent and gallic acid as a standard. 14 The absorbance (Abs) was measured at 725 nm. The content of total phenolic was expressed as milligram gallic acid equivalents (GAE)/gram dried extract.
Antioxidant activities determination
Measurement of reducing power
The reducing power was determined according to the method of Górnaś et al. 14 The Abs was measured at 700 nm against a blank. A higher Abs indicates a higher reducing power.
Chelating ability on ferrous ions
Chelating ability was determined according to the method of Dinis et al. 15 The percentage was calculated using the following equation:
Metal chelating effect (%) = (1 − (Abssample/Abscontrol)) × 100
Experimental animals
Male Swiss Albino mice weighing between 18 and 22 g (8-weeks old) were used for the study. Mice were obtained from the Pasteur Institute of Tunisia and were housed under laboratory conditions (12 h light–dark cycle and temperature of 25 ± 2°C) for 1 week before the start of the experiment. The experimental protocol was duly approved by the local ethics committee of Tunisia University for the use and care of animals in conformity with the National Institutes of Health recommendations.
The mice were divided randomly into five experimental groups including a control group, model group, Donepezil (DON) group, DPE, and LPE group. Then they were orally pretreated with saline (0.9%), roasted defatted peanuts extracts (100 mg/kg), or donepezil (3 mg/kg) for 21 days. DON is a reversible, selective AChE inhibitor that is currently approved for the symptomatic treatment of AD; it is believed to inhibit the breakdown of the neurotransmitter ACh and compensate for the deficiency of ACh in the brain. 16 On day 19, 20, and until day 28, mice were subcutaneously injected with water or scopolamine (1 mg/kg body weight) 15 min after roasted defatted peanuts extracts/water feeding. During the test session, drug administration continued 45 min before the trial, and scopolamine was given to all groups except the control group 30 min before the trial. The dose of roasted defatted peanuts extracts was determined based on acute toxicity results.
The acute toxicity study of the roasted defatted peanuts extracts (Dark and Lite) was carried out using the “Up-and-Down” method of testing in mice at single doses of 50, 150, 375, 750, 1500, and 2000 mg/kg boy weight following the Organization for Economic Development (OECD) guideline no. 425. 17 Six male mice were used for each dose level in the study. An animal was picked at a time, weighted, and dosed with the equivalent volume of extract dissolved in distilled water. The extract was administered orally. Each animal was observed after dosing for the first 5 min for signs of regurgitation and kept in a metallic cage. Each was then observed every 15 min in the first 4 h after dosing, every 30 min for 6 h and 24 h for behavioral signs of toxicity (changes in the skin, hair, eyes, mucous membranes, and respiratory, circulatory, autonomic, and central nervous systems, motor activity, convulsion, tremors, salivation, diarrhea, lethargy, or sleep) according to the specifications of the OECD. 17
Behavioral assessments
Open-field test
Mice were placed in a Plexiglas test apparatus (76 × 76 × 42 cm) and recorded for 5 min. 18,19 For analysis, the open-field arena was divided into 25 equal squares by a 5 × 5 grid. The center square was defined as the central zone, in which the animal's activity is usually regarded as a measure of anxiety. The time mice spent in the central zone and the numbers of squares crossed in the open arena were quantified. 20 Activities of control mice and drug-treated mice were monitored alternatively to avoid order effects.
MWM task
MWM was effectuated in a circular pool (65 cm diameter × 35 cm height) with a black escape platform (6 cm diameter × 15 cm height). The location of the platform can be discriminated by a visual cue. The water within the pool was made opaque by the addition of 15-mL India ink to track the rat's swim paths proficiently and more importantly to make the platform invisible. The pool was divided into equal quadrants. A platform was placed in one of the quadrants, 1 cm below the surface as described previously. 20,21 Data were recorded using a video camera connected to the corresponding software AnimalTracker. 22
Mice were given an acquisition trial for 4 days. Each day, the mouse was released at one of four locations on the edge of the pool and allowed to swim freely. The trial ended when the mouse reached and climbed onto the platform. If it had not climbed onto the platform after 60 sec, the mouse was placed on the platform. The mouse remained on the platform for 30 sec after which it was removed and the next trial began. The escape latency was recorded during each acquisition trial. On fifth day, all mice were subjected to probe trials without the platform and were recorded for 60 sec. The time spent in the target quadrant and the crossing numbers over the position at which the platform had been located were measured for spatial learning and memory.
Biochemical parameters
At the end of experimentation on the 28th day, 60 min after the last dose of saline, scopolamine (1 mg/kg body weight), or roasted defatted peanuts extracts (100 mg/kg), all the animals were sacrificed by cervical dislocation. The hippocampus was removed and homogenized in phosphate buffer (w/v: 1/40, 50 mM, pH 7.4) using a homogenizer (15 bursts), centrifuged at 5500 rpm at 4°C for 15 min, and the collected supernatant was used to estimate AChE activity, reduced glutathione (GSH), malondialdehyde (MDA) levels, and total protein by the methods described by Ellman and Callaway, 23 Ohkawa et al., 24 and Lowry et al., 25 respectively. Hippocampal ACh levels were determined using a commercially available ACh assay kit (Sigma) according to the manufacturer's protocol.
Data analysis
Analysis of variance (ANOVA, one-way ANOVA) followed by Newman–Keuls test was used to analyze the data. The criterion for statistical significance was P < .05.
Results and Discussion
Color parameters
The color of peanuts after roasting is an important measurement of the extent of MR and correlates with the generation of brown products. The total difference (L*, a*, b*, and
CIE Lab Coordinates of Defatted Unroasted and Roasted Peanuts Extracts at Different Time
Values are given as mean ± SD (n = 3).
Different superscript letters in each column indicate significant difference at P < .05.
ΔE: color differences; L * : lightness (0 = black, 100 = white); a * : (−a * = greenness, +a * = redness); and b * : (−b * = blueness, +b * = yellowness). * RPE: refers to “raw peanuts extract” without roasting; LPE: refers to “light peanuts extract” roasting temperature at 187°C for 8.6 min; DPE: refers to “dark peanuts extract” roasting temperature at 187°C for 12.7 min.
SD, standard deviation; CIE, the International Commission on Illumination.
Larger changes were observed for color attributes in LPE and DPE when compared with RPE due to the longer processing times. L* of the sample decreased from 86.9 ± 1.7 to 54.3 ± 1.1 and 43.3 ± 1.2 in RPE, LPE, and DPE, respectively. On the other hand, a* value of the sample increased from 0.5 ± 0.1 to 1.3 ± 0.2 and 6.3 ± 0.4 in RPE, LPE, and DPE, respectively, while b* value increased from 11.6 ± 0.6 to 19.5 ± 0.9 and 33.1 ± 1.1 for the same processes. The total color difference of 33.5 ± 2.3 and 49 ± 0.8 was observed in the LPE and DPE processed samples, respectively, with a significant (P < .05) difference among them. Changes in the color parameters (L*, a*, and b*) are attributed to Maillard and browning reactions that produce brown pigments with a low and high molecular weight in the advanced stage of the browning reaction. 26
Change in browning and FI of defatted peanuts extracts
The effect of roasting time on the extent of the MR of defatted extracts is given in Table 2. Results showed an increase in the Abs at 294 and 420 nm of LPE and DPE with increasing heating time (P < .05). In fact, during the development of the MR, colorless intermediate that absorbs high UV are formed. 27 The progress of the reaction involves the formation of brown compounds with a high molecular weight such as melanoidins that absorb visible light at 420 nm. 28,29 Our results showed that the Abs of LPE and DPE were 0.3 and 0.56 at 240 nm and 0.5 and 1.19 at the 420 nm, respectively (Table 2). Accordingly to these results, previous data reported by Żyżelewicz et al. 29 who found that the heating time increase the production of intermediate compounds and melanoidin during the MR. Results also showed that the FI of products formed during MR from LPE and DPE was 1970 and 10,000 relative fluorescence units (RFU), respectively (Table 2). Indeed, the significant rise in the Abs and FI of defatted extracts during roasting at 187°C were correlated with color parameters level (L*, a*, b*, and ΔE*), confirming the development of MR by the formation of intermediate compounds and their polymerization reactions during thermal treatment that contribute to the generation of fluorescent and colored compounds. 13,30
Effect of Roasting Time in Absorbance and Fluorescence Intensity of Defatted Peanuts Extracts
Values are given as mean ± SD (n = 3).
Different superscript letters in each column indicate significant difference at P < .05.
LPE: refers to “light peanuts extract” roasting temperature at 187°C for 8.6 min; DPE: refers to “dark peanuts extract” roasting temperature at 187°C for 12.7 min.
RFU, relative fluorescence units.
TPCs and antioxidative activity
Phenolic compounds from peanuts are a mixture of phenolic acids, flavonoids, and tannins that contribute to TPC and antioxidant potential of peanuts extracts with a synergistic effect. 14,31 Results showed that the TPC increased from 48.2 mg GAE/g of extract in RPE to 62.6 and 66.8 mg GAE/g in LPE and DPE, respectively. Our results are in agreement with Win et al. 31 who reported that the TPC of peanut kernel flour (Arachis hypogaea L.) increased significantly as a function of roasting times. Additionally, Boateng et al. showed that heat treatment breaks down insoluble phenolic compounds, leading to a better extractability of phenolic compounds. Indeed, several studies indicated that heat treatment is an effective method to increase the TPC in the food system. 31,32
The reducing power and ferrous chelating ability assays were used as the first tools to investigate the effect of roasting in the antioxidant activity of peanut extracts. As shown in Figure 1, the antioxidant activity on reducing power and ferrous chelating ability agrees with previous studies that demonstrated that roasting improves the antioxidant activity of food. 32,33 Likewise, the defatted light peanuts heated for 8.6 min showed better reducing power than dark and raw peanuts at all concentrations (Fig. 1a), indicating that lightly roasted peanuts showed good hydrogen donating activity (HDA). In Figure 1b, the iron-chelating ability of the various defatted peanuts extracts was 78%, 84%, and 92% for raw, light, and dark peanuts extract at a concentration of 1 mg/mL, respectively (P < .05). From these results, the reducing power and the chelating ability of roasted peanuts samples with different heating times correlated well with the extent of MR. The products formed (intermediate stages or/and melanoidins) and liberated (phenolic compounds) during roasting could play an important role in the antioxidant activity of peanuts extracts. 12,14,31,34 Additionally, intermediate stages products (reductones) formed during the first stage of roasting exhibit better reducing power due to their high HDA. 28,34
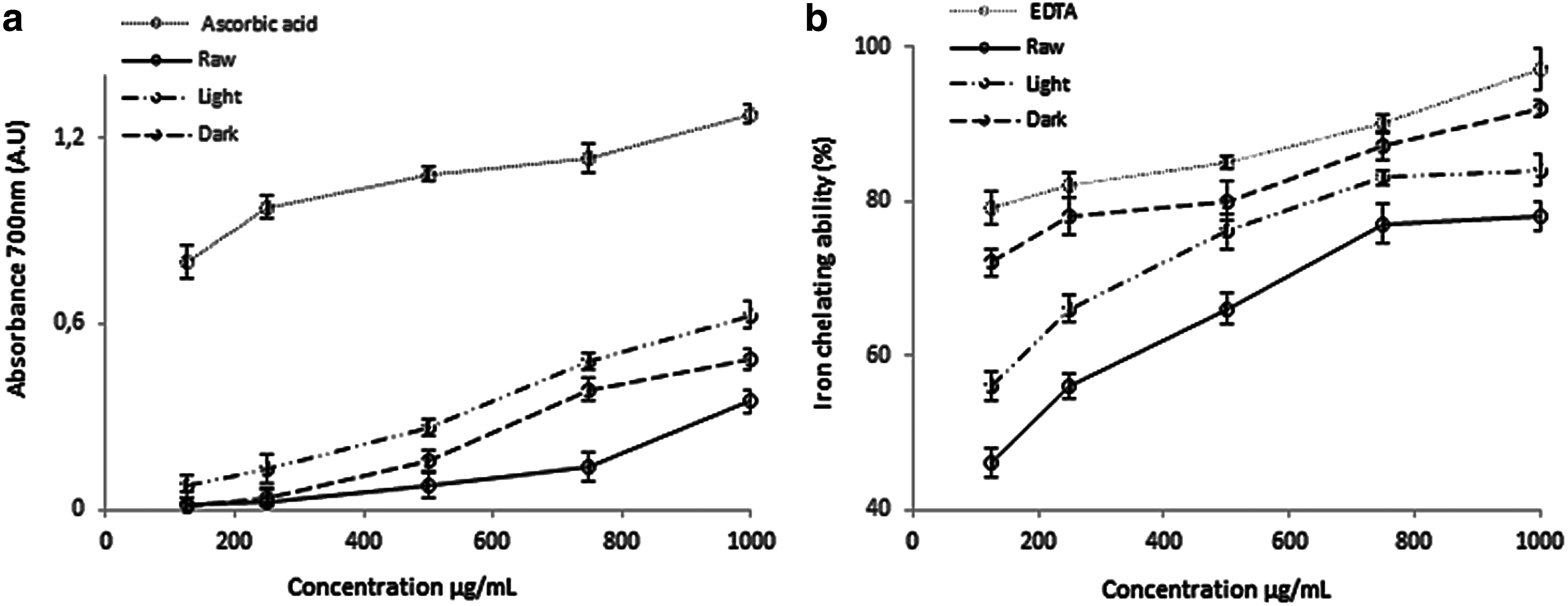
Antioxidant activities of raw, light, and dark defatted peanuts extract.
Behavioral assessments
Defatted peanut extracts did not affect the locomotor activities of mice
Before assessing the behavioral changes in memory performance an open-field test was additionally performed to preclude any perplexing motor impairments, which may have an impact on results in memory-specific behavioral tasks. 35 Here, two parameters of open-field behavior are measured: (i) latency to move from the central square and (ii) the number of squares crossed during 5 min period (Fig. 2). Data analysis of latency to move from the central square and the number of square crossing by one-way ANOVA revealed no significant differences between groups [F (5,17) = 0.271, P = .893], [F (5,17) = 1.609, P = .207 respectively]. These results showed that the locomotor function did not change between all experimental groups in the open-field test, which proposes that the amnesic effects of scopolamine had no significant impact on motor performance.
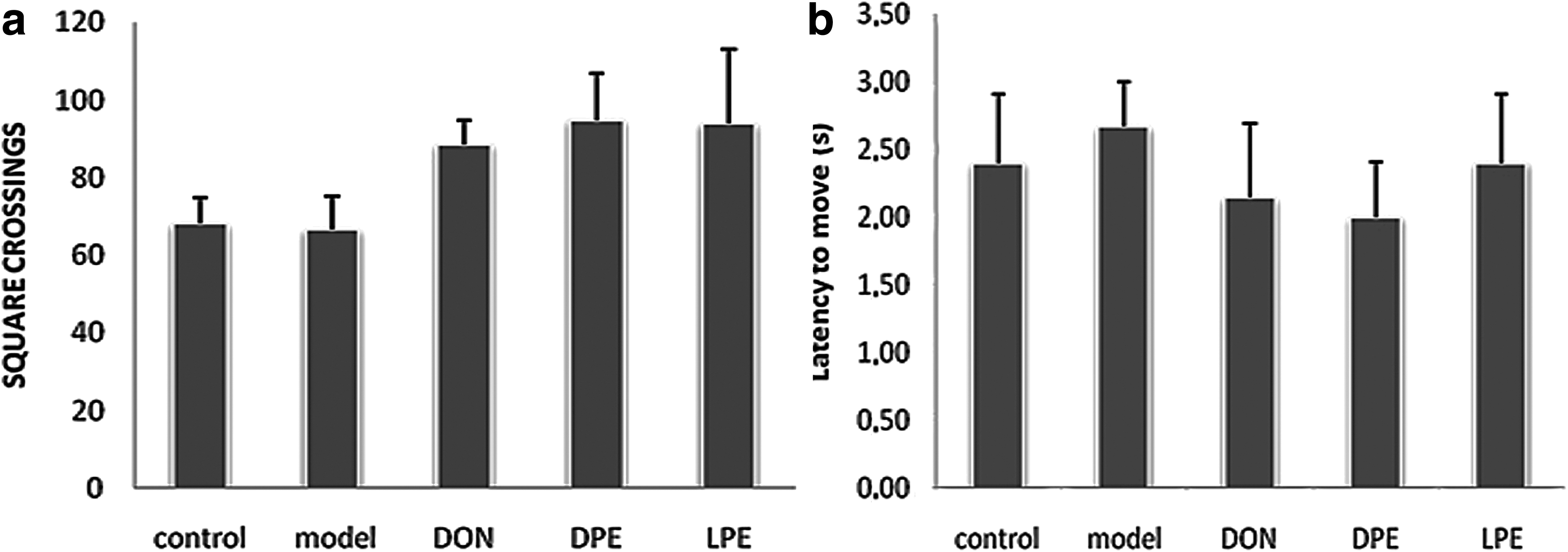
Locomotor activity was evaluated by open field test in terms of latency to move from the central square
Anti-amnesic effects of peanuts flour extract in the MWM task
Clinically, AD is characterized by an insidious deterioration of memory, associated with functional disability and neurobehavioral dysfunctions. 36 Despite the availability of different treatment strategies, the severity and prevalence of this disease are not yet under control. However, complementary and alternative medicines including herbal drugs, phytochemicals, and crude extracts are being utilized in the treatment of memory disorders in particular in AD. 37 The current hypothesis about the processes by which neurons come into apoptotic or necrotic mechanisms has led to consider that the therapeutic use of natural antioxidants may be beneficial in aging and neurodegenerative diseases. 38 In this study, the ability of roasted defatted peanuts rich in MRPs and TPC to improve cognitive impairment and oxidative stress was assessed in mice model of amnesia. In this mouse model, memory impairments were induced by scopolamine (1 mg/kg) and the neuropharmacological effects of peanuts flour extract were evaluated through the MWM Test. Since 100 μg/mL of roasted defatted extracts improves memory impairment in mice, and this concentration was used in co-treatment with scopolamine. Mice in these groups learned to find the hidden platform, and escape onto it. In contrast, scopolamine-induced memory impairment made escape latency of the scopolamine group significantly higher than that of the control group during all trial sessions (Fig. 3a). Moreover, escape latencies of DON, LPE, and DPE groups significantly decreased compared with the scopolamine group on days 2, 3, and 4. However, the treatment with DPE + scopolamine exhibited a little higher protection than LPE + scopolamine at the same concentration of 100 μg/mL. At the probe trial session, the results indicated that DON and LPE/DPE groups spent significantly more time and percentage of distance crossing over the platform site in the correct quadrant. (Fig. 3b, c). These results suggest that both extracts can ameliorate the induced memory and cognitive impairments made by scopolamine via the increase of cholinergic neurotransmission in mice. Further study is needed to investigate whether roasted peanuts extract might affect the retrieval of memory, another stage of the memory process.

The improvement of roasted peanuts extract on the impaired spatial memory induced by scopolamine in the navigation trial of the Morris water maze. Data were expressed as mean ± standard error of the mean (n = 7).
To investigate the effect of both extracts on the cholinergic system that controls vital aspects of memory and different cognitive functions, ACh levels in the hippocampus were measured. The hippocampus is a major region of the brain involved in learning and memory function, particularly in spatial memory. 39 It has been reported that this region of the brain is vulnerable to structural and functional damage, specifically, abnormalities in the cholinergic system after scopolamine administration. 40 As shown in Table 3, scopolamine-treated mice significantly reduced hippocampal ACh levels in the hippocampus relative to controls (P < .05). Treatment with LPE and DPE extracts (100 mg/kg) induced a significant increase in ACh levels when compared with the model group (P < .05). DON showed a significant increase in ACh levels relative to the model (P < .05). We also measured hippocampal AChE activity to elucidate the underlying mechanism of a possible roasted peanuts extracts induced increase of ACh levels. In this study, scopolamine was found to significantly increase AChE activity, which is an enzyme responsible for the degradation of ACh, the neurotransmitter essential for cognitive function. 41 This increase in AChE activity was significantly restored by DPE and LPE treatment. These observations suggest the modulation of cholinergic neurotransmission by increasing the availability of ACh levels in the central cholinergic synapses and/or prevention of cholinergic neuronal loss.
Effect of Rosted Peanuts Extract on Acetylcholine Level, Acetylcholinesterase Activity, and Antioxidant Status in Mice Brain
The mice brain biochemical parameters are expressed as mean ± standard error values.
P < .05 DON, LPE, and DPE versus scopolamine treated group (model).
P < .05 control group versus model.
ACh, acetylcholine; AchE, acetylcholinesterase, GSH, glutathione; MDA, malondialdehyde; DON, donepezil.
Oxidative stress, a major characteristic of aging, has been considered the main risk factor for amnesia. Indeed, the excessive formation of oxidative metabolites, including ROS, contributes to the disorder of calcium homeostasis, increased lipid peroxidation product (MDA), and subsequently elevated neurotoxic activity. 42 Reducing or preventing the generation of oxidative stress using antioxidants may, therefore, present a promising and effective therapeutic strategy for treating human amnesia. The antioxidative effects of roasted peanuts extracts was investigated using an in vivo scopolamine model as it encompasses notable oxidative damage. In our study, we observed an increase in MDA concentrations and a decrease in GSH levels (Table 3). The overexpression of MDA and GSH induced by scopolamine was attenuated after treatment with LPE and DPE in the hippocampal region, which is a brain region vulnerable to oxidative stress. 43 These protective effects are probably due to the presence of phenolics compounds (coumaroyl-rhamnose, feruloyl pentoside, and cis-coutaric acid); the major phenolic compounds in peanut flour 44 and antioxidant compounds generated during the MR including reductones, melanoidins, volatile compounds (nitrogen heterocyclic), and peptides-sugar complexes. 31,45 In the same context, a study conducted by Lee et al. 46 showed the inhibitory effect of a tyrosine-fructose MRP on the amyloid-β generation and inflammatory reactions in astrocytes and microglial BV-2 cells. 46 The mechanism of antioxidative effects of MRPs was poorly investigated. It was generally believed that these effects are closely related to the types, molecular weights, and structures of MRPs generated during the MR.
A recent study has reported that levels of AChE splice isoforms were increased under conditions of oxidative stress. 47 Therefore, the relationship between AChE activity and oxidative biomarkers was analyzed. According to the Pearson correlation analysis, a significant correlation is observed between the levels of MDA and AChE activity (data not shown). Our findings agree with those of a previous study 48 and suggest a possible interaction between scopolamine-induced cholinergic modulation and oxidative stress. However, further research is needed to clarify the mechanism underlying the interaction and to determine channel(s) or receptor(s) are involved in cholinergic transmission after the administration of extracts. The determination of the extract's chemical composition is also of interest to have a clearer idea about the origin of the observed effects. Further behavioral testing and a larger number of mice with longer treatment duration are necessary for a general conclusion.
Conclusions
Despite the limitations mentioned above, data from our study suggest that roasted peanuts extracts administration ameliorated cognitive deficits measured by performance in water maze tasks. Both treatments (LPE and DPE) reduced AChE activities and increased ACh levels in the scopolamine-induced model. Moreover, these treatments suppressed scopolamine-induced oxidative damage by ameliorating the increased levels of MDA and decreased levels of GSH. The results indicate that the underlying mechanism of learning and memory improvement may involve modulations of the cholinergic system and the reduction of oxidative stress. Thus, these findings provide evidence for the potential of roasted peanuts extracts as a natural, alternative treatment for amnesia.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This research was supported by the Tunisia Ministry of Higher Education, Scientific Research and Information and Communication Technologies (LR18ES47).