
Abstract
Flavonoids—compounds abundant in balanced daily diets—have been extensively investigated for biological activity. The pronounced antiproliferative effects of flavonoids have prompted studies to elucidate their mode of action against tumor cells. The anticancer properties of myricetin, a 3′,4′,5′-tri-hydroxylated flavonol, have been confirmed for a number of neoplasms, but myricitrin, its 3-O-rhamnoside derivative found in fruits and other parts of edible plants, has been scarcely investigated as a chemopreventive agent. This study evaluated the antiproliferative potential of myricitrin obtained from Combretum lanceolatum (Combretaceae) against MCF7 (breast), PC-3 (prostate), HT-29 (colon), 786-0 (kidney), and HL-60 (acute promyelocytic leukemia) cancer cell lines, using the sulforhodamine B and tetrazolium salt assays. Myricitrin proved most effective in inhibiting growth of HL-60 cells (GI50 = 53.4 μmol·L−1), yet showed weak antiproliferative activity against other cell lines. Possible cytotoxic mechanisms involving inhibition of topoisomerases I and IIα by myricitrin were also evaluated, revealing inhibitory activity only against topoisomerase IIα. The results suggested that topoisomerase IIα inhibition is the probable mechanism responsible for the antiproliferative activity of myricitrin. In vivo mutagenicity by myricitrin and its possible antimutagenic effect on doxorubicin-induced DNA damage were also investigated by performing the somatic mutation and recombination test (SMART) on Drosophila melanogaster. Myricitrin proved nonmutagenic to the offspring of standard (ST) and high-bioactivation (HB) crosses, while cotreatments with doxorubicin revealed the antimutagenic properties of myricitrin, even under conditions of high metabolic activation.
Introduction
Flavonoids comprise secondary polyphenolic metabolites found naturally in many foodstuffs, such as grains, fruits, vegetables, and teas 1 —hence the importance of investigating both the potential toxicity and medicinal properties of these compounds. In addition to their industrial utility as preservatives and as coloring and flavoring agents, flavonoids are potent antioxidants, as well as cardioprotective and anticarcinogenic agents. 2,3
The mechanisms identified for the antineoplastic properties of flavonoids have revealed antioxidant, anti-inflammatory, and antiproliferative effects. 4 This class of compounds is also known to inhibit topoisomerases, an effect possibly related to antiproliferative activity. 5,6 These nuclear enzymes control torsional stress in DNA double strands during cell division and catalyze DNA topological isomerization by transient breakage and resealing of strands. 7 Inhibition of topoisomerase activity—related to blockade of the cell proliferation cycle, in which maladjustments play a major role in cancer development—is viewed as a desirable property of chemopreventive agents. 8
Myricetin (

Structures of the flavonoids myricetin (
The present study—as part of our group's ongoing investigation of the Pantanal biome flora of Mato Grosso do Sul State, Midwest Brazil, for compounds not only potentially antitumoral but also exhibiting chemopreventive properties against mutagenic agents—reports the isolation of myricitrin from branches and leaves of a specimen of Combretum lanceolatum Pohl ex Eichler (Combretaceae) collected from this biome. To further contribute toward elucidating the cytotoxicity mechanism of myricitrin, its ability to inhibit topoisomerase I and topoisomerase IIα was also investigated. In addition, to investigate its chemopreventive potential, the somatic mutation and recombination test (SMART) on wing cells of Drosophila melanogaster was performed to assess mutagenic and antimutagenic properties, using standard (ST) and high-bioactivation (HB) crosses.
Materials and Methods
Materials
Silica gel 60 RP-18 (230–400 mesh; Merck) and Sephadex LH-20 (Amersham Biosciences) were used for column chromatography. Thin layer chromatography (TLC) was performed on precoated silica gel 60 F254 plates (Merck). The plates were developed using a solution of cerium sulfate (2.0 g) in 2% H2SO4(v/v), natural products polyethylene glycol reagent (NP-PEG)—that is, a 1% methanol (MeOH) solution of 2-aminoethyldiphenylborinate (w/v) + a 5% ethanol solution of polyethylene glycol (w/v)—and ultraviolet (UV) radiation. 1 H and 13 C NMR spectra were obtained at room temperature in CD3OD (Cambridge Isotope Laboratories) on a Bruker DPX-300 spectrometer operating at 300.13 MHz ( 1 H)/75.47 MHz ( 13 C).
Collection of plant material and preparation of C. lanceolatum extracts and phases
Leaves (1.65 kg) and branches (1.70 kg) of C. lanceolatum were collected from Corumbá county, Mato Grosso do Sul State, in March 2013 and identified by Arnildo Pott, of the Universidade Federal de Mato Grosso do Sul (UFMS). A voucher specimen (CGMS 49230) has been deposited at the CGMS Herbarium at the UFMS. Research on Brazilian biodiversity was conducted under permit number A5DBC20.
Powdered air-dried leaves (1.65 kg) and branches (1.70 kg) were separately extracted for 5 days in 95% ethanol at room temperature. Concentration under reduced pressure yielded syrupy extracts. To achieve partial separation of major constituents, a roughly 80.0 g amount of each extract was resolubilized in MeOH:H2O 1:1 (v/v) and subsequently partitioned with hexane, dichloromethane, ethyl acetate (EtOAc), and n-butanol to yield the corresponding hexane, dichloromethane, EtOAc, and n-butanol phases, respectively.
Myricitrin isolation from C. lanceolatum
TLC revealed the presence of flavonoids in both EtOAc phases (EtOAc-L, 8.65 g, obtained from leaves, and EtOAc-B, 9.93 g, from branches) after plate elution with CHCl3:MeOH 9:1 (v/v), followed by spraying with NP-PEG and inspection under UV radiation.
21
These phases (1.10 g each) were separately chromatographed on RP-18 silica gel columns eluted with an MeOH:H2O gradient (2:8, 4:6, 8:2), MeOH, and CHCl3, each phase yielding fractions F1 to F5. Flavonoids were concentrated in both fractions F2 (eluted with MeOH:H2O 4:6), as shown by TLC monitoring (NP-PEG), revealing compound
Myricitrin (
Antiproliferative activity and cell cultures
MCF7 (ATCC-HTB-22, breast carcinoma), HT-29 (ATCC HTB-38, colon carcinoma), 786–0 (ATCC-CRL-1932, kidney carcinoma), PC-3 (ATCC-CRL-1435, prostate carcinoma), and HL-60 (ATCC-CCL-240, acute promyelocytic leukemia) cells were cultured in RPMI 1640 medium supplemented with 1% penicillin–streptomycin and 10% bovine fetal serum (Invitrogen) and kept in a humidified incubator at 37°C in a 5% CO2 atmosphere. 22
Cytotoxicity tests
Sulforhodamine B
Cytotoxicity was tested on adherent cells (MCF7, HT-29, 786–0, and PC-3) in 96-well plates (test plates). The cells were seeded at a density of 7500 cells/well and exposed 24 h later to four myricitrin concentrations (0.25, 2.5, 25, and 250 μg·mL−1) in triplicate (Fig. 2). The test compound was dissolved in dimethylsulfoxide while ensuring the final concentrations of the latter (0.25% at the highest sample concentration) did not affect cell viability. Doxorubicin (0.025–25 μg·mL−1; DXR) was used as the positive control. At 48 h of exposure, the cells were fixed by adding 20% trichloroacetic acid and subsequently stained with sulforhodamine B (SRB; 0.1%) diluted in acetic acid. 23

Effect of myricitrin (0.25, 2.5, 25, and 250 μg·mL−1) on cell viability in five human cancer cell lines. Results obtained from triplicate SRB and MTT assays. MTT, [3-(4,5-dimethylthiazol-2-yl)-2,5]-diphenyltetrazolium bromide; SRB, sulforhodamine B.
Tetrazolium salt—[3-(4,5-dimethylthiazol-2-yl)-2,5]-diphenyltetrazolium bromide
For nonadherent cells (HL-60), inoculation density was 25,000 cells/well. At 48-h exposure of the test samples (Fig. 2), and at the same concentrations used in the SRB assay, [3-(4,5-dimethylthiazol-2-yl)-2,5]-diphenyltetrazolium bromide (MTT) dye (Sigma) was added (final concentration: 0.5 mg·mL−1). This soluble dye is metabolized only by living cells and mitochondrial enzymes, yielding formazan, a colored insoluble product. 24
Data analysis of cytotoxicity assays
For both methods described above, absorbance values were read at 540 nm on SpectraMax 190 microplate reader (Molecular Devices) and growth percentages were calculated according to Monks et al. 25 Antiproliferative activity was expressed as the drug concentration that inhibited cell growth by 50% (GI50). Growth was determined by nonlinear regression using Origin 6.0 software (OriginLab). Data are shown as mean ± standard deviation, and statistical significance was analyzed using an unpaired Student's t-test or a one-way analysis of variance. P < .05 was considered statistically significant.
DNA-topoisomerase inhibiting activity
Inhibition of topoisomerases I and IIα was evaluated by converting supercoiled plasmid DNA (pBR322, 0.5 μg mL−1; Thermo Scientific) into relaxed DNA. 26 –28 The reactions were prepared by incubating the positive controls (100 μM etoposide, which inhibits topoisomerase IIα, and 100 μM irinotecan, an inhibitor of topoisomerase I) and/or the test samples with plasmid DNA (0.25 μg) at 37°C for 1 h in the presence of topoisomerase IIα reaction buffer (0.5 M Tris-HCl at pH 8.0, 1.50 M NaCl, 100 mM MgCl2, 5 mM dithiothreitol, bovine serum albumin [BSA] at 300 μg·mL−1, and 20 mM ATP) or, for topoisomerase I, 10 × buffer (10 mM Tris- HCl at pH 7.9, 1 mM EDTA, 1.5 M NaCl, 1% BSA, 1 mM spermidine, and 50% glycerol), followed by addition of two units of human TOP enzymes (topoisomerase I or IIα from Sigma).
All reagents, except positive controls and myricitrin, were added to the negative controls. The reaction was stopped by immersion in an ice bath for 20 min and addition of stop buffer (50% glycerol, 10% sodium dodecyl sulfate, and 25% bromophenol blue), followed by addition of proteinase K (1 mg·mL−1), keeping the mixture at 55°C for 40 min. The reaction products (negative control, positive control, and test) were separately (10 μL) applied to wells containing 1% agarose in Tris-borate-EDTA buffer, along with 3 μL of Blue Juice buffer 10 × , subjected to 65 V for 2 h 40 min, and subsequently visualized under UV radiation and photographed using a PowerShot SX10 IS camera (Canon). Values of n reported in Figure 3 indicate independent biological repeats.
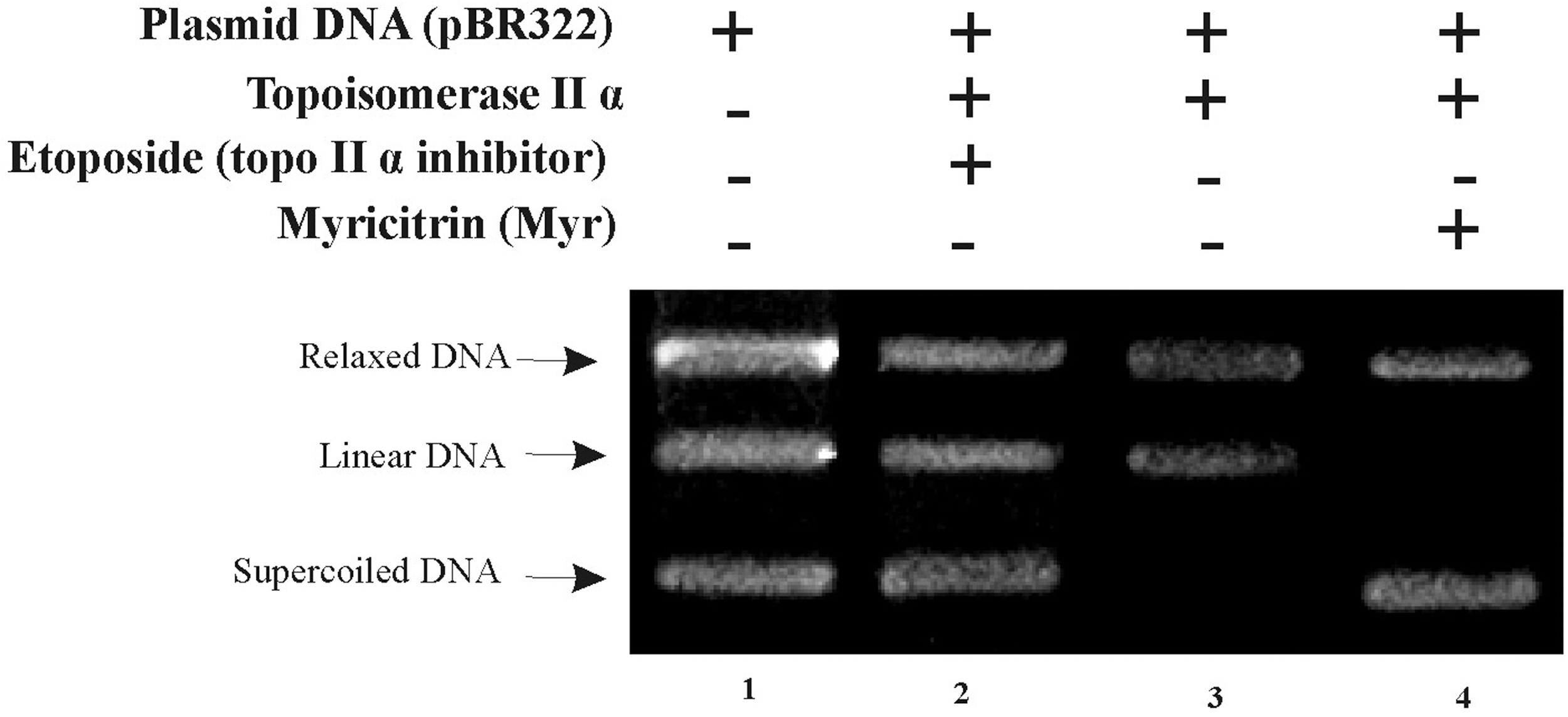
Inhibitory effect of myricitrin on human topoisomerase IIα. Line 1: 0.25 μg of pBR322. Line 2: 0.25 μg of pBR322 + 2 U of topoisomerase IIα + 100 μM etoposide (positive control). Line 3: 0.25 μg of pBR322 + 2 U of topoisomerase IIα (negative control). Line 4: 0.25 μg of pBR322 + 2 U topoisomerase IIα + 100 μM myricitrin. Reproducible results after two independent experiments (n = 2).
SMART on somatic cells of D. melanogaster
To evaluate the mutagenic effect of myricitrin and its protective potential against the mutagenicity of DXR (0.2 mM), the SMART assay was performed on D. melanogaster. The experiments were carried out according to previously established procedures. 29,30 DXR was used as the positive control for its strongly mutagenic activity in the SMART assay with D. melanogaster, inducing all types of wing spots. 31
Treatment and analysis of mutant strains
The SMART assay was performed on three mutant D. melanogaster strains: (1) multiple wing hairs (mwh), with genetic constitution y;mwhjv; (2) flare-3 (flr 3 ), with genetic constitution flr3/In(3LR)TM3, rippsep l(3)89Aa bx34eeBdS; and (3) ORR; flare-3, carrier of the marker gene ORR; flr 3 /In(3LR)TM3, rippsep l(3)89Aa bx34e eBdS. Two types of crossing were performed: (1) the ST cross, between mwh males and virgin flr 3 females, 30 and (2) the high-bioactivation cross, between mwh males and virgin ORR;flare-3 females. 29 The HB cross inherits chromosome 1 and 2 from a DDT-resistant Oregon R line, carrying genes responsible for a higher level of metabolizing enzymes of the cytochrome P450 type, which confer high sensivity to promutagens and procarcinogens. 32
Eggs of ST and HB crosses were collected over an 8-h breeding period in culture flasks containing a solid agar base (4% w/v agar in water) covered by a layer of yeast (Saccharomyces cerevisiae) supplemented with sugar. Groups of third-instar trans-heterozygous larvae (72 ± 4 h) were collected from both crosses, and transferred to glass vials containing 1.5 g of alternative culture medium (Yoki instant mashed potato flakes, from Yoki Alimentos) and subjected to mutagenicity and antimutagenicity evaluation protocols. The experiments were carried out at 23°C ± 1°C in a 12-h photoperiod, at 60–70% relative humidity.
For evaluation of mutagenic activity, 5 mL aliquots of myricitrin (0.5 × 10−2, 1 × 10−2, and 2 × 10−2 mmol·L−1) were added to the culture medium.
For antimutagenicity evaluation, myrictrin samples (0.25, 0.5, and 1.0 mmol·L−1) were associated with the mutagenic agent DXR at 0.2 mmol·L−1. For both protocols, DXR (0.2 mmol·L−1) and solvent (Milli-Q water with 1% Tween-40 and 3% ethanol) served as the positive and negative controls, respectively. All samples were tested in independent experiments. The results of duplicated experiments were pooled after verifying that no significant differences had occurred between repetitions.
Havin fed on the culture medium until completing the larval stage (roughly 48 h), emerging adults of both crosses (ST and HB), carrying the marker-trans-heterozygous genotype (MH; mwh +/+ flr 3 ) or the balancer-heterozygous genotype (BH; mwh +/+ TM3, Bds ), were collected and stored in 70% ethanol. Wing blades were mounted on slides in Faure's solution (30 g of gum arabic, 50 g of chloral hydrate, 20 mL of glycerol, 100 mL of water) and examined for the occurrence of different types of mutant spots using an optical microscope at 400 × magnification. Induced mutations were detected as single mosaic spots on the wing blades of surviving adults that showed either mwh or flr 3 phenotype. Mutant wing spots were classified as (1) small single spots (one or two mwh or flr 3 spots), (2) large single spots (three or more mwh or flr 3 spots) caused by different genetic events—namely, mitotic recombination, point mutations, and chromosomal aberrations—or (3) twin spots (both mwh and flr 3 genotypes) originating exclusively from mitotic recombination between the flr 3 proximal marker and the centromere of chromosome 3. 29,30
Statistical analysis
Data analysis was performed as per Frei and Würgler, 31 to yield the following possible statistical diagnoses: positive, weakly positive, inconclusive, or negative. For assessment of mutagenic and antimutagenic effects, the frequencies of each mutant spot type per wing in a treated series were compared with the concurrent negative and positive (DXR) controls using the Kastenbaum—Bowman conditional binomial test 33 with significance levels set at α = β = 0.05.
Inhibition percentages were calculated from the control-corrected frequencies of clones per 105 cells (total number of mutant spots/number of individuals/approximate number of analyzed trichomes for each individual) obtained from individuals treated with myricitrin associated with DXR, 34 as follows: (DXR alone − myricitrin plus DXR) × 100/DXR.
Results
Myricitrin (compound
Antiproliferative activity
Evaluated at 0.25, 2.5, 25, and 250 μg·mL−1, myricitrin proved mildly cytotoxic to adherent MCF7, HT-29, 786-0, and PC-3 human neoplastic cells after 48-h incubation (GI50 = 95.0–182.8 μM; Table 1). Activity was higher, however, against nonadherent HL-60 cells (GI50 = 53.4 μM), and proliferation was inhibited in a dose-dependent manner. As shown by the curves depicted in Figure 3, myricitrin exhibited a stronger activity against HL-60 cells when compared with HT-29, PC-3, and MCF-7 cells, and negative control at the concentrations of 25 and 250 μg·mL−1, and against 786-0 cells at the concentration of 25 μg·mL−1 (P < .05). It had a cytostatic effect on all cell lines (growth inhibition, shown by points above zero on the curves), as well as a cytocidal effect (cell death, shown by points below zero on the growth curves). As expected, myricitrin proved less active than the positive control doxorubicin, since this anthracycline-type antibiotic is ranked among the most effective chemotherapy drugs against cancer.
Antiproliferative Activity (GI50, μM) of Myricitrin on Human Cancer Cell Lines
GI50: dose that inhibits cell growth by 50%. Human tumoral cell lines: MCF7 (breast), HT-29 (colon), 786–0 (kidney), PC-3 (prostate), and HL-60 (acute promyelocytic leukemia). Values represent means ± standard deviations of three independent experiments.
Topoisomerase-inhibiting effects
The inhibitory effect of myricitrin at 100 μmol·L−1 on topoisomerase IIα is demonstrated in Figure 2, which depicts the products of plasmid DNA (pBR322) relaxation at 0.25 μg of topoisomerase IIα. This DNA concentration failed to inhibit topoisomerase I (data not shown). Topoisomerase IIα, however, was inhibited, as shown by the supercoiled DNA band (Fig. 2, line 2).
Mutagenicity and antimutagenicity
Tables 2 and 3 summarize the mutagenic and antimutagenic effects of myricitrin (SMART assay) on the offspring of ST and HB crosses of D. melanogaster chronically treated with different concentrations of myricitrin alone or combined with a fixed DXR concentration. DXR was selected for the antimutagenicity assay for its well-known potent carcinogenic properties, including mutagenic, aneugenic, and clastogenic effects that manifest irrespective of enzymatic activation. 36
Frequencies of Mutant Spots in the Wings of Drosophila melanogaster mwh/fl 3 Descendants Derived from the Standard and High Bioactivation Crosses, After Chronic Treatment of Larvae with Different Concentrations of Myricitrin (MYR)
Statistical diagnosis according to Frei and Würgler 31 : +, positive; w+, weakly positive; −, negative; i, inconclusive. m, multiplication factor for the assessment of significantly negative results. Significance levels: α = β = 0.05 when compared with respective control.
Including rare flr 3 single spots.
Considering mwh clones from mwh single and TWS.
flr 3 , flare-3; HB, high-bioactivation; LSS, large single spots; mwh, multiple wing hairs; SSS, small single spots; ST, standard; TS, total spots; TWS, twin spots.
Frequencies of Mutant Spots in the Wings of Drosophila melanogaster mwh/fl 3 Descendants Derived from the Standard and High Bioactivation Crosses, After Chronic Treatment of Larvae with Different Concentrations of Myricitrin Associated with Doxorubicin
Statistical diagnosis according to Frei and Würgler 31 : +, positive; w+, weakly positive; −, negative; i, inconclusive. m, multiplication factor for the assessment of significantly negative results. Significance levels:α = β = 0.05 when compared with respective control.
Including rare flr 3 single spots.
Considering mwh clones from mwh single and TWS.
Induction frequencies corrected for spontaneous incidence estimated from negative controls.
Calculated according to Abraham. 34
DXR, doxorubicin; NC, negative control.
On the mutagenicity evaluation (Table 2), no statistically significant differences in the frequencies of any mutant spot category were observed after treatment of larvae with myricitrin, at any concentration tested, when compared with the spot frequencies observed in the respective negative controls, for both ST and HB crosses. The latter carries genes responsible for high levels of constitutively expressed metabolizing enzymes of the cytochrome P450 complex, conferring high sensivity for promutagens and procarcinogens. 32,37 As shown in Table 2, for both ST and HB progeny, the total frequency of mutant spots per fly ranged from 0.60 to 0.70 and from 0.55 to 0.70, respectively, whereas in concurrent negative control groups, these values were 0.45 and 0.60, respectively. Evaluated for its potential to prevent DXR-induced DNA damage (antimutagenic activity) (Table 3), myricitrin significantly inhibited mutagenic events caused by DXR in both ST (61.85–85.44%) and HB (49.10–74.89%) progenies when administered at three concentrations (0.25, 0.50, and 1.0 mmol·L−1) in association with DXR (0.2 mmol·L−1), relative to larvae treated with uncombined DXR.
Discussion
C. lanceolatum Pohl ex Eichler (Combretaceae) is a tropical shrub with a wide distribution in Brazil at all latitudes.
38
Ethanol extracts of leaves and branches of this species proved rich in flavonoids, as shown by TLC after detection with NP-PEG followed by UV inspection. Myricitrin (
The dose-dependent inhibition of neoplastic cells obtained for
Evaluation of mutagenic and antimutagenic potentials is an important step in the quest for natural products with biological activity. In the present study, myricitrin was evaluated for the first time for its mutagenic and antimutagenic activities using the in vivo SMART assay with D. melanogaster.
The growing use of this insect as a powerful model organism in studies of mutagenicity and other aspects related to gene function in human diseases, particularly developmental disorders, neurological disorders, and cancer, relies, among other factors, on the high degree of evolutionary conservation of D. melanogaster genome sequencing, compared with the human genome. 45 Also, roughly 75% of all human-disease genes have related sequences in D. melanogaster. 46 These and other relevant factors support the use of D. melanogaster as a biological model for medical research on a large number of genetic disorders. 46 In addition, technical advantages of using the fruit fly as a model organism include its simple, inexpensive maintenance under laboratory conditions and the generation of a large number of individuals, given the short life cycle of this species.
At the concentrations tested, myricitrin (
Evaluated for antimutagenicity on the ST cross and, under metabolic activation, on the HB cross in the present study, myricitrin exhibited a protective effect against DXR-induced mutagenicity even on HB cross offspring. This was evidenced by a significant reduction in the frequencies of all spot types (small single, large single, and twin), after cotreatment of larvae of both crosses with varying concentrations of myricitrin and DXR, relative to uncombined DXR treatment. These results reveal the chemopreventive properties of this flavonoid against DXR-induced DNA damage even after enzyme metabolization, demonstrating myricitrin to be an antimutagenic agent.
Antioxidants can reduce or prevent cancer development by blocking genetic damage—hence the interest in finding promising chemopreventive agents among these compounds. 50,51 Flavonoids act as antioxidant agents through several mechanisms, including free-radical scavenging. 50,52 One possible mechanism involved in the protective effect observed for myricitrin against DXR-induced mutagenicity in the present study might therefore be attributed to the well-known ability of myricitrin to scavenge reactive oxygen species, since one of the mutagenic mechanisms attributed to DXR is the generation of free radicals and oxidative stress. 53 Similarly, owing to its antioxidative properties, myricitrin had previously been found to attenuate DXR-induced myocardial damage by counteracting oxidative stress and enhancing the activity of antioxidant enzymes. 54
The study results here reported help to elucidate the mechanism of action of myricitrin, fostering the search for highly selective chemotherapeutic agents. Further studies are needed to establish the benefits of daily intake values (via dietary supplements or natural medicinal products) for flavonoids as chemopreventive agents—an approach also applicable to myricitrin, based on the present results.
Footnotes
Acknowledgment
Dr. Arnildo Pott (CGMS Herbarium, UFMS, Campo Grande, MS, Brazil) is acknowledged for his assistance in the identification of the plant material.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
The authors are grateful to Fundação de Apoio ao Desenvolvimento do Ensino, Ciência e Tecnologia do Estado de Mato Grosso do Sul-FUNDECT-MS (grants 025781 and 020793), and Coordenação de Aperfeiçoamento de Pessoal de Nível Superior- (grant 001) for their financial support. The authors also thank CNPq (T.V.F. and P.S.M.) and FUNDECT-MS (C.P.D.) for the grants awarded.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.