
Abstract
Different species of the genus Pouteria have been used in folk medicine for the treatment of inflammation, fever, ulcers, diabetes, and diarrhea. We analyzed the phytochemical profile of the hydroethanolic extract from Pouteria ramiflora leaves by electrospray ionization ion trap tandem mass spectrometry and high-performance liquid chromatography-diode array detection, and examined whether it alone and in combination with cisplatin interfered with cell proliferation and death processes in HepG2 (human hepatocellular carcinoma) and FGH (human gingival fibroblasts) cells. Five compounds were identified in the extract: gallic acid, myricetin-3-O-α-
Introduction
Cisplatin is frequently used as chemotherapeutic agent to treat different carcinomas, and acts through mechanisms that damage DNA due to the formation of adducts and induce apoptotic cell death. The high incidence of chemoresistance among patients treated with cisplatin 1 has been a major challenge for cancer treatment due to recurrence, metastasis, and death. 2 A strategy to circumvent resistance to cisplatin is to associate it with other drugs, but synthetic compounds often have strong adverse effects. 3 Natural products from plants may be a safer alternative, since the combination of medicinal plants with conventional therapies has provided beneficial effects on survival, immune modulation, and quality of life of patients with cancer. 4
Synergism is the property of enhancing the efficacy of a chemotherapeutic agent in a co-treatment protocol, with the advantage of decreasing its toxicity to the patient. Thus, compounds with such property can be used as adjuvants to improve the efficacy of therapeutic interventions. 5 This approach can reduce drug resistance and provide anticancer benefits, such as reduction of tumor growth and metastatic potential, cell cycle arrest, and apoptosis induction. 6
Several species of medicinal plants belonging to the genus Pouteria, family Sapotaceae, are used in folk medicine to treat fever, inflammation, skin eruptions, ulcers, diabetes, diarrhea, nausea, vomiting, and back pain. 7 Pouteria ramiflora, popularly known as curriola, is widely distributed in South America, produces edible fruits, has been used to treat dysentery, pain, inflammation, hyperlipidemia, ulcers, and kidney disorders, and displays antinociceptive and anti-inflammatory properties. 8 –11
A previous study from our research team has demonstrated that the hydroethanolic extract from P. ramiflora leaves is cytotoxic toward HepG2 cells and exerts protective activity against the carcinogen benzo[a]pyrene, which supports further studies about its biological effects. 12 This study aims to analyze the chemical profile of the same extract, examine its antiproliferative activity, and characterize the type of cell death that it induced alone and in combination with cisplatin in HepG2 and FGH cells, with the ultimate goal of proposing an alternative treatment for cancer.
Materials and Methods
Chemicals
Cisplatin (CAS: 15663-27-1), Hoechst 33342 (CAS: 23491-52-3), fluorescein diacetate (CAS: 596-09-8), propidium iodide (CAS: 25535-16-4), and the analytical standards myricetin (98%), quercetin (98%), and gallic acid (99%) were purchased from Sigma-Aldrich (St. Louis, MO, USA). Myricetin-3-O-β-
Plant material and extract preparation
P. ramiflora (Mart.) Radlk. leaves were collected by Prof. F.F. Morais, PhD, from the State University of Mato Grosso, in September 2011, in the city of Cáceres, Mato Grosso State (MT), Brazil (16°21′1770“S; 58°18′2760”W). A voucher specimen was identified and deposited at the São Paulo State University “Júlio de Mesquita Filho” (UNESP) Herbarium of Rio Claro (SP-Brazil), under the collection number HRCB 58166. The P. ramiflora leaves were dried at 40°C to constant weight and ground in a Wiley mill. The plant material was submitted to exhaustive percolation with 70% ethanol solution (ethanol:H2O 70:30, v/v), 14 and the resulting extract was evaporated to dryness under low pressure at 40°C, providing 15.04% of the extract. A 50 mg/mL stock solution of the hydroethanolic extract was used in biological experiments.
ESI-IT-MS/MS analysis
The hydroethanolic extract of P. ramiflora leaves was analyzed in the LTQ-XL mass spectrometer (Thermo Fisher Scientific, Waltham, MA, USA), in electrospray ionization mode (ESI), and by fragmentation in multiple stages (MS/MS) using an ion trap type interface (IT). The negative mode was chosen. For the generation and analysis of the mass spectra, the following conditions were set: capillary voltage of −4 V, spray voltage of −5 kV, capillary temperature of 280°C, and carrier gas (N2) flow of 60 (arbitrary units). The acquisition range was m/z 150–1000 with two or more scan events performed simultaneously in the LTQ-XL mass spectrometer. The first event was a complete scan (full scan) of the mass spectrum to acquire the data of ions in the established m/z range. Subsequently, MS/MS experiments from the first scan data to preselected precursor ions were performed with collision energy between 25% and 30% of the total energy of the instrument. The Xcalibur 1.3 software (Gibco/Thermo Fisher Scientific, Grand Island, NY, USA) was used for the acquisition and processing of spectral data.
High-performance liquid chromatography-diode array detection analysis
The extract preparation consisted of a cleaning up step by solid-phase extraction, as described by Costa et al.
15
The chromatographic conditions and materials used were also described by the same authors. Based on the compounds quantified by Tuttis et al.
12
in the hydroethanolic extract of P. ramiflora leaves and those partially elucidated by ESI-IT-MS/MS in this study, the extract was coinjected with 10% standard (standard:extract, m/m) of gallic acid, myricetin-3-O-β-
Cell culture conditions
The cell line HepG2, isolated from human hepatocellular carcinoma, and the human primary cell line FGH, established from the gingival biopsy of a normal patient, were kindly provided by the Nutrigenomics Laboratory of the School of Pharmaceutical Sciences of Ribeirão Preto (FCFRP/USP—Brazil). The cells were cultured in Dulbecco's modified Eagle's medium low glucose culture medium supplemented with 10% fetal bovine serum (Gibco/Thermo Fisher Scientific, Waltham, MA, USA), 0.06 g/L penicillin, and 0.10 g/L streptomycin (Sigma-Aldrich). Cultures were maintained at 37°C, under a 5% CO2 atmosphere with 95% relative humidity.
Cell treatment
Preliminary cytotoxicity assays based on 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) reduction, neutral red uptake, and lactate dehydrogenase release were performed to determine the extract concentrations to be used in this study. The results supported the choice of noncytotoxic extract concentrations (0.5 and 1.0 μg/mL), as well as the concentration that reduced cell viability by ∼13% in the MTT assay (2.0 μg/mL). 12
As the extract concentration of 1.0 μg/mL was the lowest concentration tested, which exerted cytotoxic action in the apoptosis assay, we selected it to continue investigating the mechanisms of action of the extract alone and in combination with cisplatin. To examine how P. ramiflora extract (1.0 μg/mL) associated with cisplatin (2.0 μg/mL) acted, three treatment protocols were used: (1) pretreatment (PRE), where cells were treated with the extract for 24 h and with cisplatin for further 24 h; (2) simultaneous treatment (SIM), where the extract and cisplatin were simultaneously added to the reaction medium and the cells were treated for 24 h; and (3) post-treatment (POST), where cells were treated with cisplatin for 24 h and with the extract for further 24 h.
Triple staining apoptosis/necrosis assay
To analyze the cell death rate, 1 × 105 HepG2 and FGH cells were seeded in six-well plates, treated with the extract and/or cisplatin, and labeled with fluorochromes (final culture concentration of 4.0 μg/mL Hoechst 33342, 7.5 μg/mL fluorescein diacetate, and 1.0 μg/mL propidium iodide). A total of 600 cells/treatment were analyzed using the Axioplan 2 Mot epifluorescence microscope (Carl Zeiss, Jena, Germany) at 400 × magnification. The criteria for considering cells as viable, apoptotic, or necrotic were described by De Santis et al. 16 (Fig. 1).

Cell staining patterns of viable, necrotic and apoptotic cells differentially stained with Ho, FD, and PI. FD, fluorescein diacetate; Ho, Hoechst 33342; PI, propidium iodide.
Cell cycle kinetics
To analyze the cell cycle kinetics by flow cytometry, 4 × 105 HepG2 cells were seeded in six-well plates, treated with the extract and/or cisplatin, and fixed in 70% ice-cold ethanol. Ethanol was removed and the pellet was suspended in 0.5 mL phosphate-buffered saline (PBS), 25 μL RNase (1 mg/mL), and 25 μL propidium iodide (1 mg/mL). After incubation for 10 min, the material was analyzed on the FACSVantage™ cell sorter flow cytometer (Becton Dickinson, Franklin Lakes, NJ, USA). The acquired data were analyzed using the Cell Quest 3.1 software (Becton Dickinson) and the cell cycle profiles were outlined using the Summit 6.2 software (Beckman Coulter, Brea, CA, USA).
Immunocytochemistry
This assay was performed following the manufacturer's recommendations, with modifications. HepG2 cells (1 × 105) were seeded on coverslips placed in six-well plates, fixed in 100% methanol, permeabilized with 1% bovine serum albumin and 0.1% Tween in PBS for 1 h, and incubated overnight, at 4°C, with antibodies to proliferating cell nuclear antigen (PCNA; 2.5 μg/mL) and cyclin D1 (1/250). Next, the cultures were incubated with anti-mouse (1/500) and anti-rabbit (4 μg/mL) antibodies for 1 h, and subsequently incubated with Hoechst 33342 (1 mg/mL) for 10 min. All the antibodies used in this assay were purchased from Abcam (Cambridge, MA, USA).
The coverslips were dried at room temperature and fixed onto microscope slides using fluorescence mounting medium (Dako, Carpinteria, CA, USA). The images were acquired using the Axioplan 2 Mot epifluorescence microscope equipped with a charge-coupled device (Carl Zeiss) and the Isis software version 5.2 (MetaSystems, Heidelberg, Germany), and analyzed using the open-source GIMP software 2.8. 17 The PCNA parameters were analyzed according to Schönenberger et al. 18
Gene expression
To analyze gene expression, HepG2 cells were seeded in T25 culture flasks (5 × 105 cells/flask) and treated with the extract and/or cisplatin. Total RNA was extracted using the PureLink® RNA Mini Kit (Life Technologies, Carlsbad, CA, USA) and quantified using the NanoDrop 2000C spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA). Next, RNA was treated with DNAse I (Amplification Grade; Invitrogen, Carlsbad, CA, USA) and reverse transcribed using the SuperScript III First-Strand Synthesis System, Oligo-dT, and random primers (Invitrogen), according to the manufacturer's recommendations. Sequences for the reference and target gene primers were generated using Gene Runner Software version 3.05, 19 except for the cyclin-dependent kinase 2 (CDK2) gene primers, which were purchased from Invitrogen (Table 1).
Genes Used in the Quantitative Reverse Transcription–Polymerase Chain Reaction Assay
Real-time PCR was performed on a Techne Quantica® Real-Time PCR Cycler System (Techne/Cole-Parmer, Vernon Hills, IL, USA) using Platinum® SYBR® Green qPCR SuperMix-UDG (Invitrogen). The gene expression data were analyzed using the Quansoft 1.35 Software (Techne/Cole-Parmer) and normalized using the Ct (cycle threshold). The relative expression of the genes was calculated according to Pfaffl. 20
Statistical analyses
Statistical analyses were performed using the GraphPad Prism software version 5.01 (La Jolla, CA, USA). Experimental data were compared using one-way analysis of variance followed by the Tukey's test, with P < .05. The relative expression of the genes was compared with the negative and positive controls using the Student's t-test, with P < .05.
The combination index (CI) was calculated using the Bliss Independence model, as described by Foucquier and Guedj, 21 and used as parameter to determine whether the plant extract and cisplatin acted in synergy in the apoptosis and flow cytometry assays. CI values lower than, equal to, and higher than 1 indicate synergy, simple additivity, and antagonism, respectively.
Results
ESI-IT-MS/MS analysis of the extract composition
ESI-IT-MS/MS fragmentation of some ions enabled partial elucidation of the chemical structure of thirteen secondary metabolites present in the hydroethanolic extract of P. ramiflora leaves (Table 2).
Compounds Identified in the Hydroethanololic Extract of Pouteria ramiflora Leaves and Their Negative m/z Values ([M − H]−) of the Molecular Ions and Main Fragments (MS 2 ) Detected by ESI-IT-MS/MS
High-performance liquid chromatography-diode array detection analysis of the extract composition
The presence of gallic acid (compound
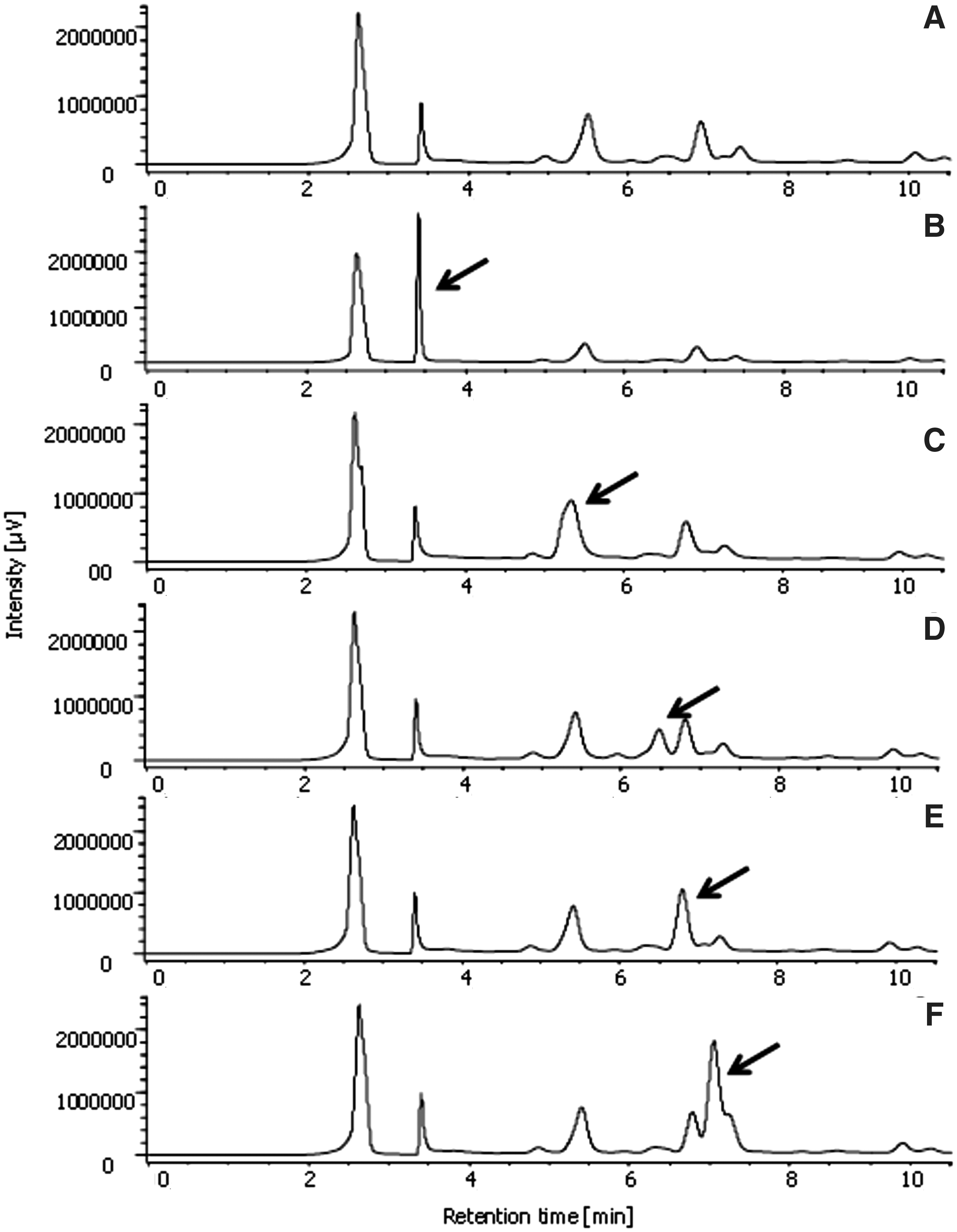
Chromatogram of the hydroethanolic extract of Pouteria ramiflora leaves alone
These results, associated with the UV spectra of the major peaks of the chromatograms of P. ramiflora extract, support that the peaks present corresponded to the phenolic compounds reported in Table 3.
Wavelength of Maximum UV Absorption (λ ma x) and Retention Times of the Peaks Identified in the Chromatogram of Hydroethanolic Extract of Pouteria ramiflora Leaves
RT, retention times.
Induction of apoptosis/necrosis
In HepG2 cells, P. ramiflora extract at 1.0 and 2.0 μg/mL and cisplatin induced apoptotic cell death, with most of the cell population in the early phase of apoptosis (Fig. 3A). The extract at 1.0 μg/mL acted in synergy with cisplatin at 2.0 μg/mL when tested using the PRE (late apoptosis, CI = 0.37) and SIM (early apoptosis, CI = 0.64 and late apoptosis, CI = 0.54) treatment protocols (Fig. 3B).
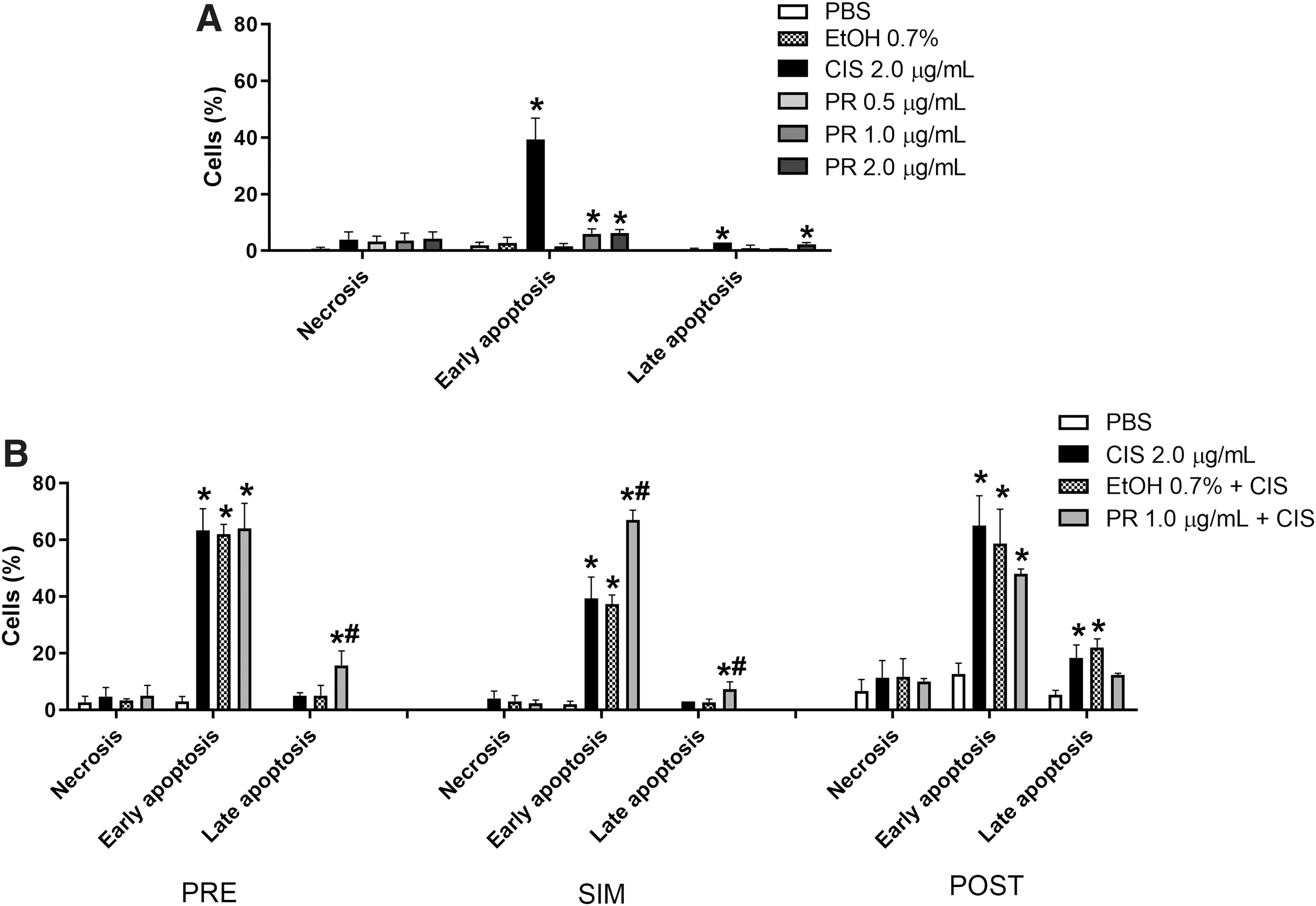
Percentage of HepG2 cells in early apoptosis, late apoptosis, and necrosis after treatment with P. ramiflora extract alone
Cisplatin and all the P. ramiflora extract concentrations tested (0.5, 1.0, and 2.0 μg/mL) increased the number of FGH cells in early apoptosis (Fig. 4A), indicating that they induced cell death by apoptosis. Synergism occurred only when cisplatin and the extract were added simultaneously to the FGH cell culture (early apoptosis, CI = 0.95) (Fig. 4B).

Percentage of FGH cells in early apoptosis, late apoptosis, and necrosis after treatment with P. ramiflora extract alone
Alteration of cell cycle kinetics
P. ramiflora extract and cisplatin altered the cell cycle kinetics of HepG2 cells, by, respectively, decreasing and increasing the G1- and S-phase cell populations. The lowest extract concentration (0.5 μg/mL) affected the cell cycle kinetics the most strongly, and augmented the G2-/M-phase cell population as effectively as cisplatin (Fig. 5A). The extract acted in synergy with cisplatin to enhance cell cycle arrest in the S phase when tested using the SIM (CI = 0.90) and POST (CI = 0.98) treatment protocols (Fig. 5B).
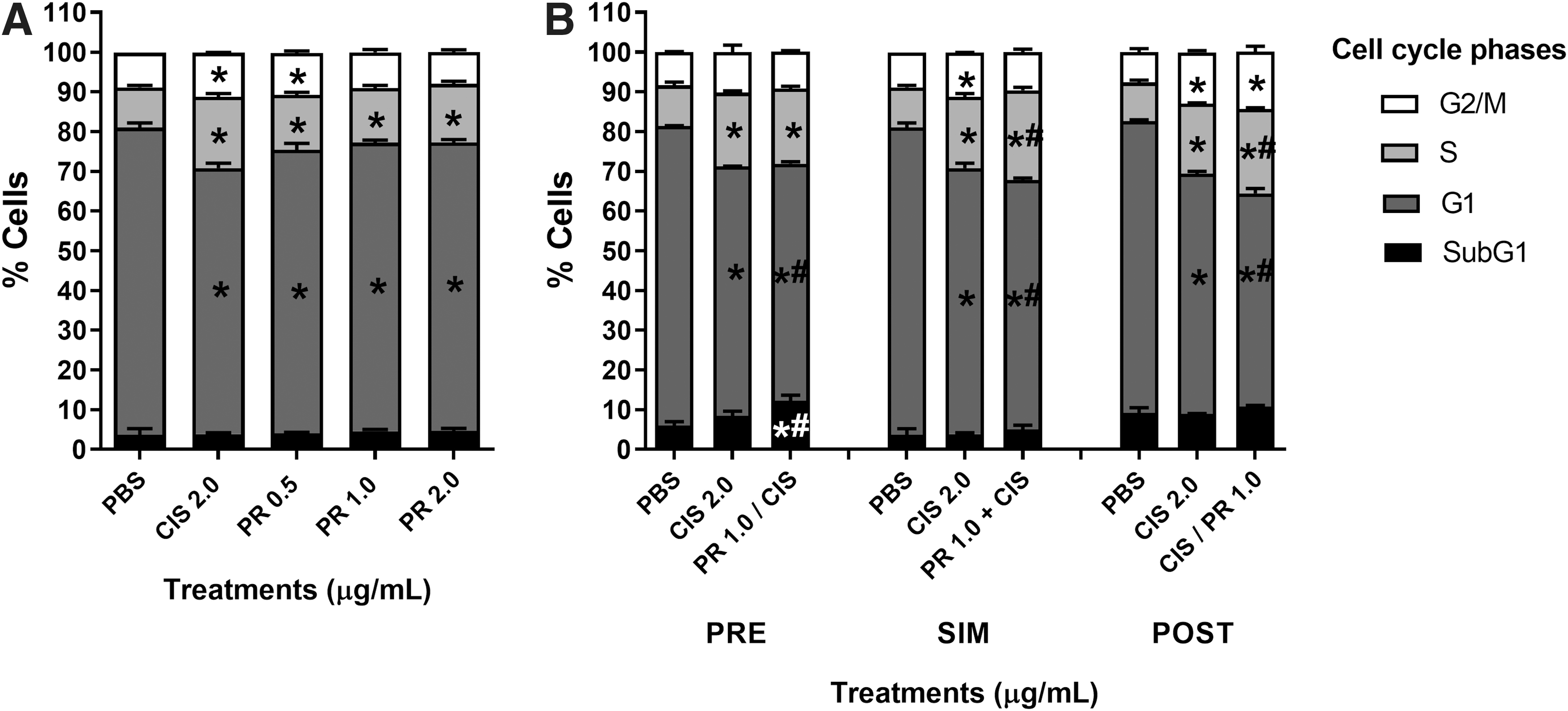
Frequency of HepG2 cells at different cell cycle phases after treatment with P. ramiflora extract alone
Immunocytochemistry
Compared with the control group, the P. ramiflora extract did not change the staining pattern of HepG2 cells with anti-PCNA and anti-cyclin D1 antibodies. In contrast, cisplatin reduced the number of cells labeled with anti-cyclin D1 antibody (G1 phase) and augmented the number of cells labeled with anti-PCNA antibody heterogeneously distributed through the nucleus (S phase) (Fig. 6A). Cell treatment using the PRE protocol raised the cell population with homogeneously distributed PCNA (G2 phase), whereas cell treatment using the SIM protocol increased the cell population with heterogeneously distributed PCNA (S phase) and decreased cyclin-D1-labeled (G1 phase) cell population (Fig. 6B).

Frequency of cyclin D1- and PCNA-labeled HepG2 cells after treatment with P. ramiflora extract alone
Gene expression
The SIM with P. ramiflora extract and cisplatin upregulated expression of the BAX (proapoptotic) and CDK2 (S phase) genes (Fig. 7).
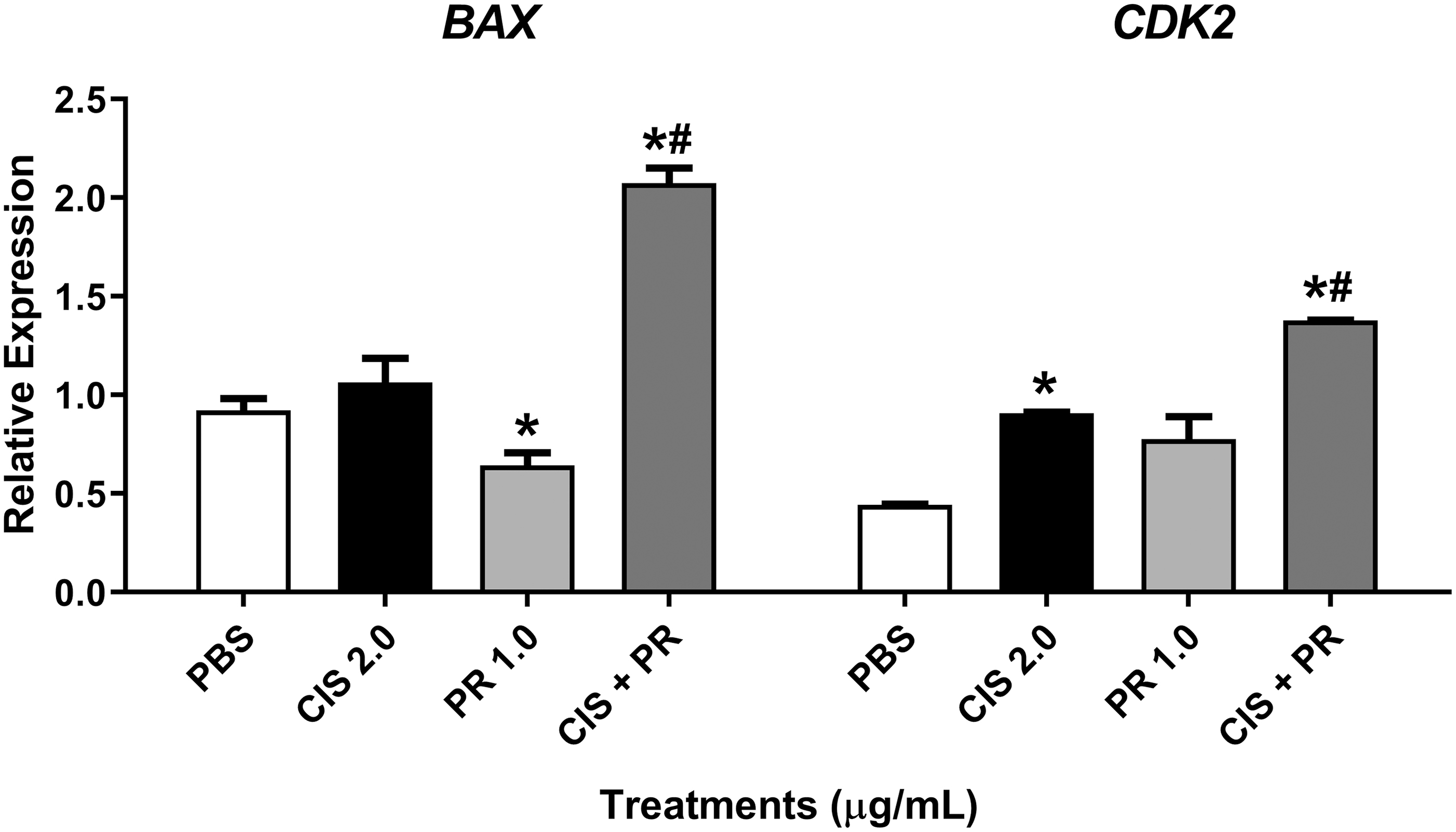
Expression of the BAX and CDK2 genes relative to the reference genes GAPDH and ACTB in HepG2 cells after treatment with P. ramiflora extract alone and combined with cisplatin for 24 h. PBS = phosphate-buffered saline (negative control), CIS = cisplatin at 2.0 μg/mL (positive control), PR = P. ramiflora extract at 1.0 μg/mL. *P < .05 versus negative control and # P < .05 versus positive control (Student's t test).
Discussion
The mass spectrometry analysis of glycoside flavonoids evidences that the binding of sugar to a hydroxyl oxygen bound to ring A or C can be easily observed by the loss of the sugar residue,
29
as detected in the quercetin- and myricetin-O-glycosides reported in Table 2. According to the literature, one can recognize that the sugar substituent is attached to position 6 of the aglycone because it gives rise to the loss of more intense ion fragments 0,2
X, 0,3
X, and
1,5
X.
30,31
We inferred that compound
The mass spectra of the quercetin-O-glycosides revealed the presence of a radical ion [M − H]−• at m/z 300, besides the ions [M − H]− at m/z 301, which corresponded to the deprotonated aglycone, as reported by Vukics and Guttman. 32 In addition to the ions [M − H]- at m/z 317, a radical ion [M − H]−• at m/z 316 was detected in the spectra of myricetin-O-glycosides. The presence of these radical ions is explained by the homolytic scission of the glycosidic bond, and the high radical ion/ion ratio suggests the presence of a 3-O-glycosidic bond in flavonoids. 29
Five compounds were identified as C-glycosides. In C-glycosylated flavonoids, the sugar is attached directly to the carbon at position 6 or 8 of the flavonoid nucleus. The C-glycosidic bond between the sugar and the flavonoid is stronger than the O-glycosidic bond and requires higher collision energies to be broken. Fragmentation occurs in the sugar ring where the bonds are weaker. 29 Another important information is that the fragmentation pathway of C-glycosides provides loss of water molecules, which is more pronounced in C-6 flavonoid glycosides. 33
The P. ramiflora extract induced apoptotic cell death in HepG2 and FGH cells probably due to the presence of these myricetin derivatives and quercetin. Various studies have reported that myricetin derivatives are proapoptotic to different types of tumor cells and modulate expression of proapoptotic genes. Myricetin induces apoptotic cell death and upregulates BAX gene expression in a concentration-dependent manner in HepG2 cells, 34 as well as it elicits apoptosis through BAX/BCL2 in colon 35 and thyroid cancer cells. 36
Although the P. ramiflora extract was more strongly cytotoxic to FGH normal cells, its synergistic effect with cisplatin was stronger in HepG2 cells. Synergism is a process in which some compounds cooperate to reach a combined effect that is greater than the sum of their separate effects. As synergistic effects may occur due to the interaction between herbal products and conventional drugs, the positive synergistic plant–drug interactions should be investigated to achieve the best outcomes, such as providing benefits to patients and avoiding adverse effects. 37
Myricetin was the most abundant compound identified in the P. ramiflora extract. This compound sensitized hepatocellular carcinoma cells to cisplatin treatment, both in vitro and in vivo. The combination of myricetin and cisplatin can synergistically inhibit HepG2 and Huh7 cell growth and induce apoptosis. 38 Myricetin enhances the therapeutic effects of cisplatin in human cervical cancer cells HeLa 39 and placental choriocarcinoma cells JAR and JEG, 40 as well as protects normal cells from cisplatin-induced colon toxicity in rats by improving the antioxidant status. 41
In the SIM protocol, the P. ramiflora extract acted in synergy with cisplatin to induce apoptotic cell death with increased expression of the BAX gene. BAX protein belongs to the group of proapoptotic proteins of the BCL-2 family. Intrinsic apoptotic stimuli, such as DNA damage or stress in the endoplasmic reticulum, activate (BH3)-only proteins that lead to BAX accumulation in mitochondria and death by apoptosis. 42,43 Therefore, our findings indicate that the SIM of HepG2 cells with the extract and cisplatin activates the intrinsic apoptosis pathway.
Cisplatin, as well as the three concentrations of P. ramiflora extract, induced cell cycle arrest at the S and G2/M phases. Cisplatin induces the formation of DNA adducts that delay the S and G2 cell cycle phases and thereby enables the repair mechanism to restore DNA integrity and prevent abnormal mitoses. 1 The P. ramiflora extract is rich in flavonoids such as myricetin, quercetin, and apigenin, and several studies have demonstrated that these flavonoids can cause cell cycle progression disorders both in vitro 44,45 and in vivo. 46 –48
In the SIM and POST protocols, the extract and cisplatin acted synergistically to delay the cell cycle, with S-phase arrest through increased expression of the CDK2 gene, and a decrease in the G1-phase cell population. CDK2 regulates the transition and progression during cell division, through phosphorylation of a large number of proteins that mediate cell cycle progression, DNA replication, histone synthesis, and centromere doubling. 49,50 CDK2 participates in the ATM/ATR-Chk2/Chk1-Cdc25A-CDK2 pathway that mediates the intra-S phase checkpoint, delays replication, and elicits DNA repair. If the repair is inefficient, the permanent cell cycle arrest, apoptosis, or oncogenesis may occur. 51
As quercetin is one of the major phytochemicals in P. ramiflora extract, our data corroborate the synergism between quercetin and cisplatin in HepG2 cells described by Zhao et al. 52 These authors have demonstrated that this association modulates the expression of different genes involved in cell cycle progression and apoptosis, reinforcing the hypothesis that the flavonoids present in the extract increased the antiproliferative effect of cisplatin reported in this study.
Cyclin D1 accounts for the entry of cells into the G1 phase of the cell cycle by activating CDK4 and CDK6. 50 The start of the increase in cyclin D1 levels elicits transition from the G1 phase to the S phase. In this latter phase, cyclin D1 is exported from the nucleus to the cytoplasm, where it will be degraded. Its degradation is fundamental for DNA synthesis in the S phase because cyclin D1 inhibits DNA replication by binding to PCNA and CDK2. 53 The HepG2 cell treatment with P. ramiflora extract associated with cisplatin, mainly using the SIM protocol, increased the cell population labeled with anti-PCNA distributed heterogeneously (S phase), with a consequent decrease in the cyclin D1-labeled cell population, corroborating the flow cytometry data. Considering that cyclin D1 overexpression has been linked to several types of cancer and that reduction of cyclin D1 levels decreases the number of proliferating cells, therapeutic agents that downregulate expression or induce degradation of cyclin D1 in vitro may be useful to treat cancer. 53
In summary, myricetin glycosides were the major constituents of P. ramiflora extract identified herein. The extract acted in synergy with cisplatin through mechanisms that possibly increase DNA damage, arrest cell cycle at the S phase as an attempt to repair this damage, and induce apoptotic cell death due to the failure to repair DNA. The synergistic effects between the extract and cisplatin, especially using the SIM treatment protocol, suggest that this extract increases the sensitivity of HepG2 cells to cisplatin and may be a therapeutic alternative for patients with chemoresistance to this chemotherapeutic agent. In addition, the synergistic effect may be weaker in normal cells. The findings of this study indicate that P. ramiflora extract may help to decrease the therapeutic dose of cisplatin and thereby reduce its adverse effects.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
The authors thank the Brazilian founding agencies São Paulo Research Foundation (FAPESP, grant n. 2009/52237-9), National Council for Scientific and Technological Development (CNPq) for research grants to W. Vilegas, E.A. Varanda, and I.M.S. Cólus, and Coordination for the Improvement of Higher Education Personnel (CAPES - Finance Code 001) for the scholarships to K.T. and D.L.M.G.C.