
Abstract
Emerging evidence suggests that aging is associated with the deterioration of immunity, a term known as immunosenescence, which may lead to a higher incidence of infections in the elderly population. Our previous studies reported that supplementation of royal jelly (RJ) extended the lifespan of Caenorhabditis elegans (C. elegans), a nematode model. The aim of this study was to investigate the potential benefits of RJ supplementation on modulation of the innate immunity in C. elegans. Using Staphylococcus aureus (S. aureus; ATCC 25923) as the infection model, we showed that RJ supplementation from the egg hatching stage could protect C. elegans against the infection. Further mechanistic studies demonstrated that RJ coordinated pathways of IIS/DAF-16, p38 MAPK, and Wnt to modulate the innate immunity. In addition, when RJ was administrated to the aged C. elegans, the worms displayed prolonged survival time to a variety of bacterial infections compared with the nontreatment group. This result indicates the RJ may help delay the innate immunosenescence.
Introduction
Aging is an inevitable process for most organisms and is associated with the progressive decline of physiological functions. During this time, the proper function of the immune system is weakened (immunosenescence) in the elderly people, which can lead to increased vulnerability to pathogenic infection and poorer quality of life. Given the increasing number of people worldwide who are entering the later stages of life, there is a greater demand to alleviate immunosenescence and fight against pathogenic infections.
Dietary intervention with nutraceuticals has been used as an effective way to prevent disease and promote health historically. Combined with medical treatment, some nutraceuticals can also provide very positive benefits that can help improve the function of the immune system. As such, the strategy that explores different nutraceuticals to determine their function on immunomodulation is an attractive and promising one. Another attractive characteristic is the edibility of nutraceuticals, which makes the safety of their clinical translation reassured once the molecular mechanisms underlying their beneficial effects have been determined.
Royal jelly (RJ) is usually produced by young worker bees, aged 4–12 days and has been consumed by human beings for many years to promote a healthy lifestyle because of its beneficial properties such as antioxidant, anti-inflammatory, antitumor, antibacterial, hepatoprotective, and so on. 1 Our recent studies demonstrated that RJ supplementation promotes the lifespan and proteostasis in Caenorhabditis elegans. 2,3 The anti-aging function prompted an investigation to determine if RJ may help to enhance immunity and delay the progress of immunosenescence. To address this concern, the innate immunity was initially studied through the genetically tractable model organism C. elegans.
Innate immunity plays a pivotal role in protecting against pathogens by blocking infections at an early stage and facilitating the elicitation of adaptive immunity. 4 As a bacterivore that lacks adaptive immunity, the nematode C. elegans has evolved a multifaceted innate immune system that includes physical barriers, avoidance behaviors, and secretion of antimicrobial molecules. Through these mechanisms, they are able to combat a variety of bacteria that are pathogenic to both human and worms. 5 –7 Of note, these responses share many elements with human innate immunity, making C. elegans a valuable model to understand the complex interactions between bacterial pathogens and host immunity.
C. elegans innate immunity is also regulated by a number of conserved signaling pathways, including the p38 mitogen-activated protein kinase (MAPK) pathway, the insulin/insulin-like growth factor-1 signaling (IIS) pathway, and the transforming growth factor β signaling pathway. 8 –11 Some of these pathways have been shown to be involved in nutraceutical-mediated prolongevity effects, 12 –14 but research on nutraceuticals' effects on host's immunity is scarce. Dissection of these signaling cascades in C. elegans will not only help understand how natural compounds modulate the innate immunity in C. elegans, but also facilitate the studies of how innate immunity is regulated in higher order organisms. 7,15
Utilizing Staphylococcus aureus (ATCC 25923) as a bacterial infection model, we demonstrated that RJ supplementation helps C. elegans defend against the pathogen and this protection was largely dependent on enhancing innate immunity. We further showed that RJ coordinated the IIS/Daf-16, p38 MAPK signaling, and Wnt signaling to regulate the defense against S. aureus.
S. aureus was chosen because to the following reasons: (1) S. aureus is a common opportunistic pathogen in humans following injuries or surgeries and it can infect a wide range of animals including C. elegans, (2) the response to S. aureus infection in mammals is mainly dependent on innate immunity, 16 (3) both living and heat-inactivated S. aureus can kill C. elegans and hence provide persistent infection stress to the innate immune system during assays, 17 and (4) despite the significant progress, the host response to S. aureus remains incomplete. Furthermore, we showed that aged C. elegans, when administrated RJ treatment, displayed prolonged survival time on different bacterial pathogens than those without RJ supplementation. This result indicated that RJ might help delay innate immunosenescence.
Materials and Methods
Strains and growth conditions
The bacterial strains used in this experiment were Escherichia coli OP50, S. aureus (ATCC 25923), Enterococcus faecalis (ATCC 47077), Salmonella typhimurium (ATCC 14028), and Vibrio cholerae C6706 (O1 El Tor isolated from Peru). 18 E. coli OP50, S. typhimurium, and V. cholerae were grown in Lysogeny broth medium, whereas S. aureus and E. faecalis were grown in brain–heart infusion (BHI) medium.
All C. elegans strains were obtained from the Caenorhabditis Genetics Center (CGC), University of Minnesota, USA. The strains were maintained at their permissive temperature on nematode growth medium (NGM) seeded with E. coli OP50 feeding strain. One hundred microliters of OP50 was dropped on the center of 60 mm NGM plates, which were allowed to dry overnight before the assays were carried out. Strains used in this study were as follows: N2 Bristol (wild type), daf-2(e1370) (CGC#CB1370), age-1(hx546) (CGC#TJ1052), daf-16(mgDf50) (CGC#GR1370), hsf-1(sy441) (CGC#PS3551), nsy-1(ag03) (CGC#AU3), sek-1(ag1) (CGC#AU1), pmk-1(km25) (CGC#KU25), axl-1 (tm1095) (CGC#KN611), dsh-1(ok1445) (CGC#RB1328), lin-17(e1456) (CGC#CB1456), egl-20(mu39) (CGC#CF263), and bar-1 (ga80) (CGC#EW15).
Preparation of RJ
The lyophilized RJ powder standardized to contain 3.85% (E)-10-hydroxy-2-decenoic acid (10-HDA) and 0.67% 10-hydroxydecanoic acid was provided by Yamada Apiculture Center, Inc. (Okayama, Japan). Each 100 g of RJ powder contains 48.6 g carbohydrates, 40.9 g proteins, 5.5 g fats, 2.8 g minerals, and 2.2 g H2O. To prepare RJ supplemented food, appropriate amount of RJ powder was suspended in sterile distilled water and incorporated into the NGM (2 mg/mL RJ) 1 day before the assay. 2
C. elegans killing assay
Synchronized worm populations were acquired by allowing 10–15 hermaphrodites to lay eggs on NGM plates (control or with 2 mg/mL RJ) seeded with E. coli OP50 for 4 h at permissive temperature, and then the parents were removed. Eggs left on the plate would reach L4 after ∼40 h. Pathogenic bacteria were individually grown in freshly prepared medium overnight with shaking at 37°C. The overnight culture was then 10-fold concentrated and dropped (50 μL) on the center of 35 mm NGM plates (containing 50 μg/mL FUDR to prevent the growth of worm progeny), 19 –22 which were allowed to dry for 2 h at room temperature before killing assays were carried out. The control plates were prepared the same way by dropping 50 μL of 10-fold concentrated overnight OP50 culture.
For each killing assay, 25–30 synchronized worms at appropriate adult stage were transferred onto each plate and their survival was monitored thereafter. All killing assays were carried out at 25°C in triplicate and a minimum of three independent trials were performed for all strains or conditions. Day 0 was defined as the day when appropriate stage worms were transferred onto the killing assay plates. SPSS software (IBM SPSS Statistics) was used to carry out all the statistical analysis. Kaplan–Meier lifespan analysis was carried out and P-values were calculated using the log-rank test. Value of P < .05 was accepted as statistically significant.
Gene expression analysis by quantitative real-time PCR
S. aureus strain was grown overnight at 37°C in BHI medium supplemented with or without 2 mg/mL RJ. The bacteria were collected and washed with phosphate-buffered saline. Total RNA was prepared using RNAzol® RT reagent (Molecular Research Center, Inc.) and stored at −80°C. Complementary DNA was synthesized using the Bioline sensifast cDNA synthesis kit (Bioline). qPCR was performed using SensiFAST SYBR No-Rox Kit (Bioline) and the CFX96 real-time PCR detection system according to the manufacturer's suggested protocol (Bio-Rad). The qPCR conditions were as follows: 95°C for 3 min, followed by 40 cycles of 10 s at 95°C and 30 s at 50°C.
Relative fold changes for transcripts were calculated using the comparative CT (2−ΔΔCT) method. 23 Cycle thresholds of amplification were determined by Light Cycler software (Bio-Rad). The transcription levels of 16S rRNA were used as internal controls to normalize the expression level of target transcripts. Each qPCR experiment was repeated three times using independent RNA preparations. The data were pooled and analyzed using unpaired Student's t-test, and P < .05 was accepted as statistically significant. The qPCR primers for sarA are 5′- GATTTTCAATTAGCTTTGA-3′ (forward primer) and 5′-CTTGTGGTTGTTTGTAGTTT-3′ (reverse primer). Primers for 16S rRNA are 5′- TGTCGTGCGATGTTGGG-3′ (forward primer) and 5′- CGATTCCAGCTTCATGT-3′ (reverse primer).
Results
RJ protects C. elegans against S. aureus infection
Our previous report showed that supplementation of 2 mg/mL of RJ could significantly promote the lifespan and proteostasis in C. elegans. 2,3 We hence adopted the same concentration to examine whether RJ supplementation may protect hosts from pathogenic infection by carrying out S. aureus killing assays in C. elegans. Specifically, wild-type C. elegans N2 worms were supplemented with RJ on NGM plates from the egg hatching stage until L4/young adult. The L4/young adult wild-type N2 worms were subsequently transferred and exposed to S. aureus in the presence of RJ. The survival of these worms was monitored every 24 h until the last worm died. Worms cultured without RJ served as the control. The mean lifespans of the control and RJ-treated are 10.92 ± 0.53 and 12.78 ± 0.53, respectively, suggesting the protective effect of RJ on S. aureus infection (Fig. 1A and Table 1).
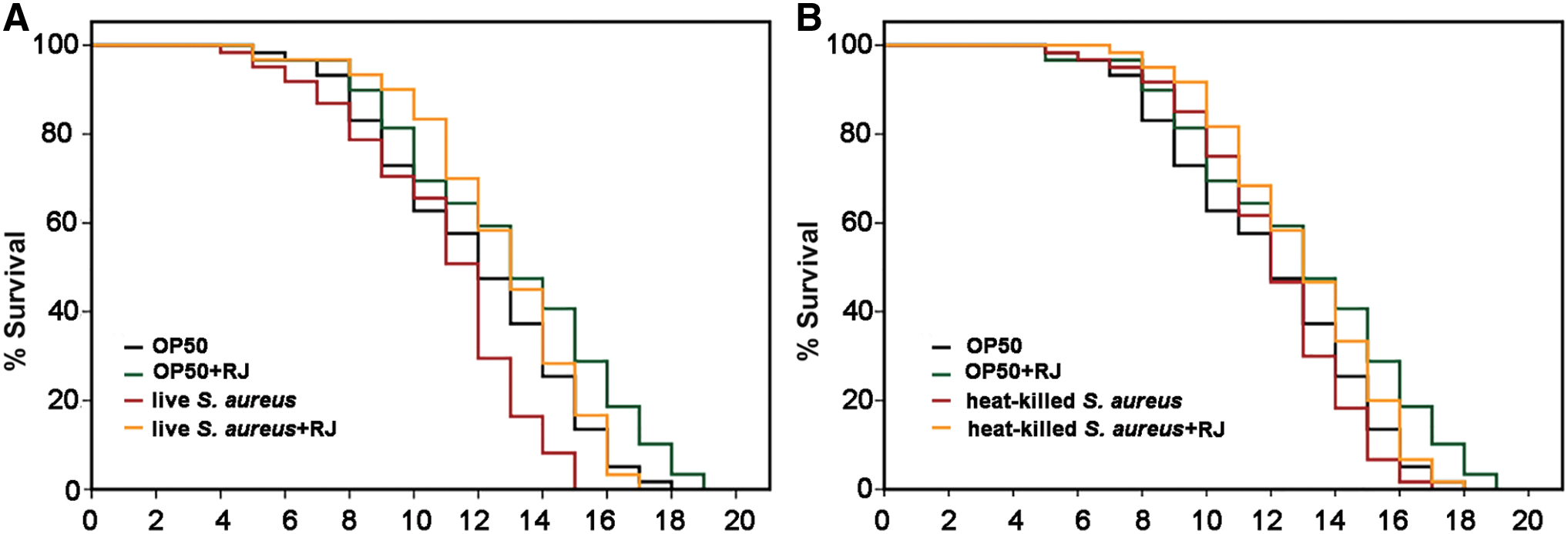
Supplementation of RJ protects Caenorhabditis elegans from live Staphylococcus aureus infection. N2 worms were exposed to live
The Survival Time of N2 Worms at 25°C When They Were Exposed to Live and Heat-Killed S. aureus
The killing assays were repeated at least three times with similar results, and the data for representative experiments are shown. The lifespan data were analyzed using the log-rank test and P-values for each individual experiment are shown.
Results presented in Figure 1A.
Results presented in Figure 1B.
The P-value was calculated by comparing N2/pathogen with the N2/OP50 group.
The P-value was calculated by comparing RJ-treated worms with the untreated worms (same bacterial food).
SE, standard error.
Considering the antimicrobial activities of RJ, we cultured S. aureus in BHI medium in the presence of 2 mg/mL RJ and monitored the growth of the bacteria. No growth inhibition of S. aureus at any of the growth phases (Supplementary Fig. S1A) was observed, indicating the prolonged survival time of C. elegans by RJ was not owing to the direct killing of the bacteria.
Because both the bacterial virulence and the capacity of the host immunity determine the severity of a bacterial infection, we therefore proposed that RJ may either attenuate S. aureus virulence, enhance the host immunity, or both. Irazoqui et al. reported that both live and heat-inactivated S. aureus could kill C. elegans. 17 We reasoned that RJ would not provide protection against dead S. aureus if it acts through attenuating the bacterial virulence. To test this, the same procedure as described by Irazoqui was used to kill S. aureus by heat. However, C. elegans fed on the heat-killed S. aureus lived as well as those fed on E. coli OP50 (Fig. 1B and Table 1), which is contradictory to the published result. Hence, we could not determine whether RJ attenuates the virulence from this experiment.
We therefore measured the expression of S. aureus sarA gene using quantitative real-time PCR (qRT-PCR). Previous studies reported that SarA (the global transcriptional regulator of virulence genes) is essential for the lethal infection in C. elegans. 24 As given in Figure 2, instead of reduction, the expression of sarA was slightly increased in response to RJ supplementation. In short, our data indicated that the RJ-mediated protection against S. aureus infection is not through reducing the SarA-dependent virulence, but we could not rule out the possibility of other virulence factors being affected by RJ.

qRT-PCR analysis of S. aureus sarA gene expression in response to 2 mg/mL RJ (normalized to 16S rDNA). Results are the average of three independent experiments, and error bars are standard error of the mean. P < .05 was considered as statistically significant. qRT-PCR, quantitative real-time PCR.
RJ acts through IIS/Daf-16 pathway to promote C. elegans innate immunity against S. aureus
DAF-16 is an important transcription factor that has been reported to modulate C. elegans immunity. 9,25 The expression of DAF-16 target genes contributes to promote worms' innate immunity and fight against bacterial infections. Depletion of DAF-16 eliminated the protective effects of RJ on S. aureus infection (Fig. 3A and Table 2). This result demonstrated the pivotal role of DAF-16 in response to RJ supplementation, suggesting RJ could improve the host innate immunity.

RJ acts through IIS/Daf-16 to promote host innate immunity in a hsf-1 independent manner.
The Lifespan of Worms at 25°C When They Were Exposed to S. aureus
The lifespan experiments were repeated at least three times with similar results, and the data for representative experiments are shown. The lifespan data were analyzed using the log-rank test and P-values for each individual experiment are shown.
Results presented in Figure 3A.
Results presented in Figure 3B.
Results presented in Figure 3C.
Results presented in Figure 3D.
Results presented in Figure 4A.
Results presented in Figure 4B.
Results presented in Figure 4C.
Results presented in Figure 5A.
Results presented in Figure 5B.
Results presented in Figure 5C.
Results presented in Figure 5D.
Results presented in Figure 5E.
The P-value was calculated by comparing RJ-treated with the untreated worms of the same genetic background.
Considering that the efficient control of intestinal bacterial accumulation is an indicator of strong gut immunity and also an important causative factor of lifespan determination, 26,27 we investigated the gut colonization of E. coli OP50 and S. aureus in N2 worms in the presence and absence of RJ. We noticed a trend of decrease of E. coli OP50 in the OP50/RJ-fed worms compared with the controls without RJ supplementation (Supplementary Table S1). In addition, when the RJ-supplemented N2 worms were exposed to S. aureus, we could hardly detect any S. aureus from the gut after 6 days of infection (Supplementary Table S1). This result supported the idea that RJ improves the innate immunity of C. elegans.
Our previous studies revealed that the anti-aging property of RJ was dependent on the activity of DAF-16 in an IIS-dependent manner. 2,3 It is thus interesting to test whether RJ may also require IIS to modulate DAF-16 and thereby promote worms' immune response against bacterial infection. 28 –30 To address this, the daf-2(e1370) and age-1(hx546) mutant worms were supplemented with RJ and their survival was monitored under S. aureus infection. As compared with the genotype-matched nonsupplemented controls, neither daf-2(e1370) nor age-1(hx546) showed prolonged survival time with RJ supplementation (Fig. 3B, C and Table 2). The epistasis analyses demonstrated that RJ did require a functional IIS pathway to protect the host against S. aureus.
Heat-shock transcription factor (HSF)-1 is another important transcription factor that contributes to promote innate immunity and fight against bacterial infections in C. elegans. 31,32 Given that IIS also regulates HSF-1, we examined whether HSF-1 was required to facilitate the RJ-mediated improvement of innate immunity. Our data showed that the absence of hsf-1 did not abolish the protective effects of RJ in C. elegans when they were exposed to S. aureus (Fig. 3D and Table 2), indicating the dispensability of HSF-1 in the RJ-mediated immunity promotion.
RJ also uses p38 MAPK pathway to protect against S. aureus infection
We next examined whether p38 MAPK signaling pathway, a conserved pathway to regulate innate immunity in C. elegans, may play a role in the RJ-mediated innate immunity. 8 –10 In brief, S. aureus was applied to the pmk-1 (km25) null mutant worms supplied with or without 2 mg/mL RJ. RJ did not extend the survival time of pmk-1 (km25) mutant worms compared with the controls without RJ supplementation (Fig. 4A and Table 2), suggesting that the protective effect of RJ against S. aureus infection need functional PMK-1, the evolutionarily conserved p38 MAPK in C. elegans.
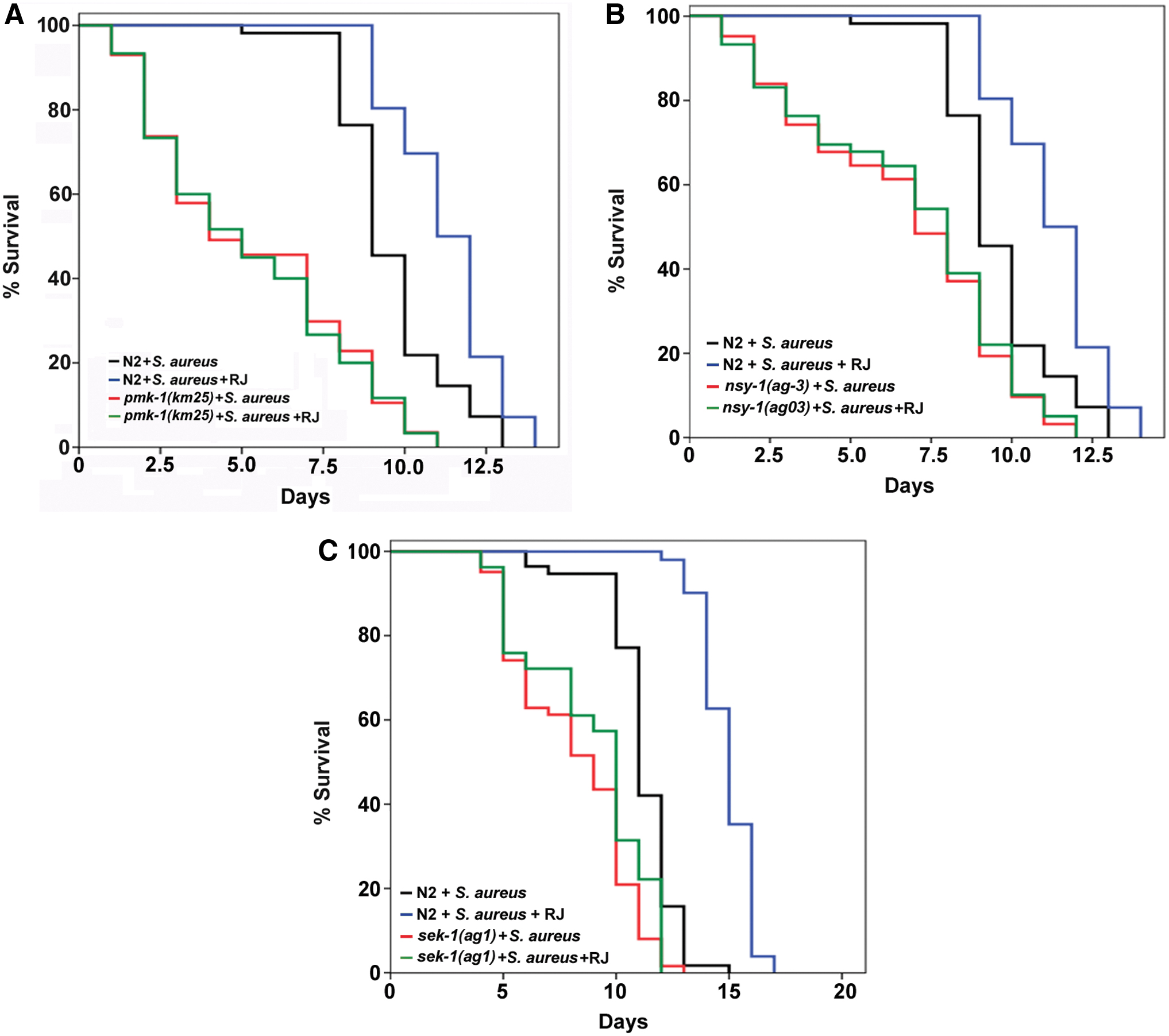
RJ-mediated protection against S. aureus is dependent on the p38 MAPK signaling pathway.
Thereafter, two upstream kinases of p38 MAPK, SEK-1, and NSY-1 were examined and similar results were obtained, that is, RJ supplementation was not able to extend the survival time of worms lacking either sek-1 (sek-1(ag1)) or nsy-1 (nsy-1(ag03)) when they were exposed to S. aureus compared with the genotype-matched nonsupplemented controls (Fig. 4B, C and Table 2). These findings together suggested that RJ acts through the p38 MAPK signaling pathway to protect C. elegans from S. aureus killing.
Canonical Wnt/β-catenin signaling is required to facilitate the RJ-mediated S. aureus resistance
bar-1 is a C. elegans β-catenin gene and plays a critical role in protecting worms from S. aureus killing. 33 The bar-1(ga80) loss-of-function mutant worm showed severe susceptibility to S. aureus infection (Fig. 5A and Table 2). The supplementation of RJ unfortunately could not protect bar-1(ga80) worms and extend their survival time when exposed to S. aureus (Fig. 5A and Table 2). This finding suggested that the functional BAR-1/β-catenin was critical for RJ to help resist against S. aureus infection in C. elegans.

RJ requires BAR-1 and the canonical Wnt signaling to protect against S. aureus.
Although bar-1 has been found to modulate the immunity, the upstream regulation of bar-1 regarding its antimicrobial function remains undetermined. To elucidate how RJ regulates bar-1 to improve innate immunity, several major components of the canonical Wnt pathway were studied because this signaling pathway has been reported to regulate the activation of BAR-1/β-catenin contributing to organismal development and cell proliferation. 34
Specifically, worm strains containing defective Wnt [egl-20(mu39)], Frizzled receptor [lin-17(e1456)], Dishevelled [dsh-1(ok1445)], and Axin [axl-1 (tm1095)] were supplemented with RJ and their survival was monitored under infection with S. aureus. As compared with the genotype-matched nonsupplemented controls, egl-20(mu39), lin-17(e1456), dsh-1(ok1445), and axl-1 (tm1095) failed to show prolonged survival time with RJ supplementation (Fig. 5B–E and Table 2). This finding suggested that RJ required, at least in part, the functional Wnt pathway to protect the host against S. aureus killing.
RJ supplementation protects aged C. elegans from diverse bacterial infections
In the aforementioned protection assays, RJ was administrated at the egg hatching stage. As aging is usually accompanied by a decline in immune function, we questioned whether RJ could provide comparable protection against different pathogenic bacteria in aged worms. In this regard, S. aureus, E. faecalis, S. typhimurium, and V. cholerae were chosen to challenge aged worms and their performance treated with or without RJ was monitored. Specifically, well-maintained N2 worms (day 6 adults) were used to represent the “middle-aged” C. elegans and exposed to each pathogenic bacterium in the presence of 2 mg/mL RJ. The survival of each worm was monitored every 24 h until all worms died. Age-matched N2 worms without RJ supplementation served as controls.
Our results demonstrated that RJ treatment significantly extended the survival time (P < .05) of worms exposed to pathogens compared with the controls, regardless of the bacterial species (Fig. 6A–D and Table 3). We also tested the effect of RJ on day 10 worms when they were exposed to various pathogens. As the mean lifespan of N2 worms (fed on E. coli OP50) at 25°C is normally 12–13 days, the day 10 worms would represent the true “aged” population. RJ supplementation notably extended the survival time of “aged” C. elegans on all four tested pathogens (Supplementary Fig. S2 and Supplementary Table S2). Again, we ruled out the possibility that RJ directly killed pathogens by monitoring the growth of each bacterium in the presence of RJ (Supplementary Fig. S1B–D).
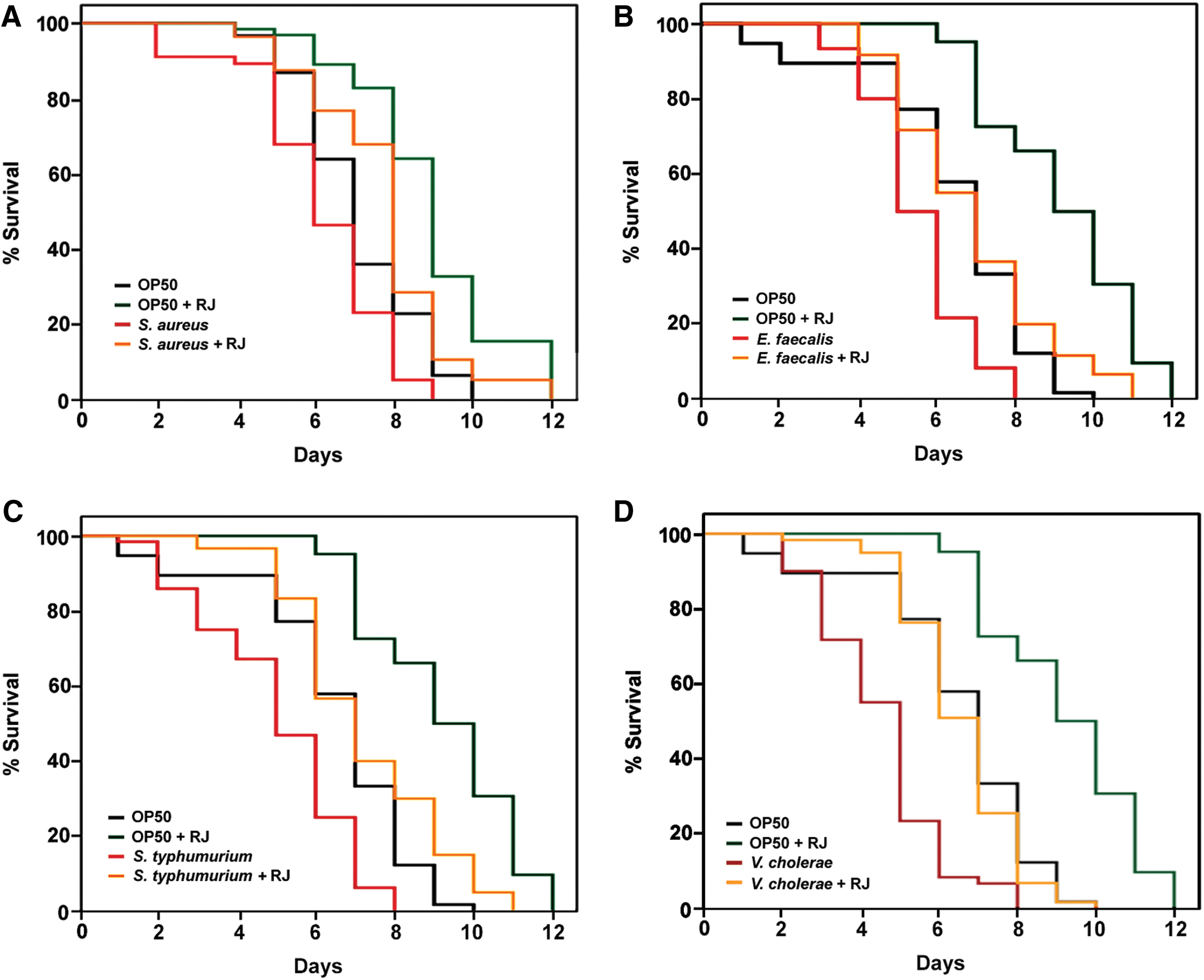
Supplementation of RJ protects day 6 C. elegans from bacterial infection in a broad-spectrum manner. RJ extends the survival time of middle-aged C. elegans under the infection of
The Survival Time of Aged N2 Worms (day 6 Adult) at 25°C When They Were Exposed to Various Pathogenic Bacteria
The killing assays were repeated at least three times with similar results, and the data for representative experiments are shown. The lifespan data were analyzed using the log-rank test and P-values for each individual experiment are shown.
Results presented in Figure 6A.
Results presented in Figure 6B.
Results presented in Figure 6C.
Results presented in Figure 6D.
The P-value was calculated by comparing N2/pathogen with the N2/OP50 group.
The P-value was calculated by comparing RJ-treated worms with the untreated worms (same bacterial food).
The resistance to a broad spectrum of pathogens indicated the promotion of host innate immunity, suggesting that RJ could retard innate immunosenescence in the aged population.
Discussion
Age-associated decline of innate immunity is called innate immunosenescence, which dramatically compromises the health of elderly populations by making them more susceptible to harmful bacterial infection. Emerging evidence has shown that some anti-aging agents helped promote immunity and highlighted their function in delaying immunosenescence. 35 Our previous studies reported the anti-aging properties of RJ, showing that RJ supplementation not only prolonged C. elegans lifespan, but also increased the capacity of their stress resistance. 2,3,36 Hence, our present studies investigated the potential preventive effect of RJ on innate immunity and its underlying molecular mechanisms.
Our results showed that RJ not only improved the resistance of C. elegans to S. aureus infection when supplemented from an early age, but also protected aged worms from a broad spectrum of bacterial pathogens. RJ-supplemented worms had a reduced intestinal bacterial load, which suggests strong gut immunity in the hosts. Further genetic analyses suggested that RJ protection depended on daf-16, rather than hsf-1, to prevent against the decline of innate immunity.
Mechanistic studies demonstrated that RJ acted through IIS cascade to regulate DAF-16. Moreover, RJ consumption also required p38 MAPK pathway and the canonical Wnt pathway to promote C. elegans innate immunity against S. aureus killing. Owing to the imperative role of IIS and DAF-16 in regulating prolongevity and innate immunity, we reasoned that RJ played a critical role in enhancing innate immunity. In addition to IIS/DAF-16, p38 MAPK pathway and the Wnt pathway are two other essential cascades modulating C. elegans immunity. The requirement of these conserved pathways in RJ-mediated immunomodulation indicated that RJ modulated the innate immunity in a systematic manner and therefore improved the overall pathogen defense in C. elegans.
To date, it remains uncertain whether DAF-16 is essential to establish the basal innate immunity, 10 although previous studies indicated that DAF-16 was an active modulator of C. elegans innate immunity. 37 We showed that RJ relied on DAF-16 to promote the host resistance to S. aureus (Fig. 3A). This finding demonstrated the essential role of DAF-16 in establishing innate immunity, especially basal innate immunity. Taken together, our results suggested that RJ supplementation enhances C. elegans innate immunity in a DAF-16-dependent manner, at least in part.
The p38 MAPK pathway is a conserved immune response pathway and plays a key role in the innate immunity. 8,38 Decline in the PMK-1, the major component of p38 MAPK pathway, was shown to be associated with the aging and increased susceptibility to Pseudomonas aeruginosa infection. 39 In consistence with this, our results indicated that the p38 MAPK pathway was indispensable for RJ to enhance C. elegans innate immunity against S. aureus.
Meanwhile, we also noticed an early study that reported that PMK-1 was not required for the induction of selected S. aureus-responsive genes. 33 In this report, researchers ruled out the importance of p38 MAPK pathway in response to S. aureus infection by measuring the expression of some S. aureus-responsive genes. To this end, they treated worms with S. aureus for 8 h and then the expression level of selected S. aureus-responsive genes was measured. The negligible changes of the target gene expression between wild-type and pmk-1 mutant worms led to the conclusion that p38 MAPK pathway may not be important in the host response to S. aureus.
Whereas in our study, S. aureus killing assay was performed to determine whether p38 MAPK pathway was important to help RJ prevent S. aureus infection. During the killing assay, worms were exposed to S. aureus over the course of their entire lives, rather than only several hours. The different experimental conditions of the two groups brought up the possibility that the long-term infection may fully activate the innate immunity, which could be dramatically different compared with the early immune response triggered by short-term acute infection. It is very likely that p38 MAPK pathway will only upregulate innate immunity against S. aureus during the early infection stage.
In addition, it is worth noting that PMK-1 regulates the expression of antimicrobial genes at both infection and noninfection conditions, 10 indicating the important contribution of p38 MAPK pathway in the basal (noninfection) and inducible (infection) innate immunity. Taken together, our results actually suggested that p38 MAPK pathway might play a prominent role in the regulation of basal innate immunity.
BAR-1 was also found to be indispensable in protecting worms from S. aureus infection, 33 but it is still unknown whether BAR-1 actively participates in the establishment of innate immunity in C. elegans. When it was found that RJ required functional BAR-1 to resist against S. aureus (Fig. 5A), it was necessary to test whether Bar-1 plays a general role in the innate immunity. To address this concern, a lifespan assay on E. coli OP50 was carried out and we found that RJ supplementation cannot prolong the lifespan of bar-1(ga80), a mutant worm with defective bar-1 (Supplementary Fig. S3). This finding demonstrated the essential role of bar-1 with regard to beneficial effects of RJ observed in C. elegans. Hence, we speculated that bar-1 may also play a general role in innate immunity.
Recently it has been shown that Wnt signaling pathway is activated downstream of the acetylcholine receptor of the muscarinic pathway in response to S. aureus infection. 40 To this end, it was hypothesized that RJ supplementation could promote gut activity by improving neuromuscular interactions. Of course, further experiments are needed to confirm this speculation.
The complex innate immune system that evolved in C. elegans is regulated through a sophisticated signal network that consists of pathways regulating longevity, stress resistance, development, and so on. 6 No doubt, the crosstalk of these pathways modulates the overall innate immunity of C. elegans. Given the primary role of the innate immunity is to prevent the occurrence of infection, it is conceivable that some core components may play a role in maintaining basal immunity. Our findings that RJ supplementation promotes host innate immunity against diverse pathogens indicated the general elevation of the basal innate immunity. IIS/DAF-16, p38 MAPK, and Wnt facilitated RJ on immunity improvement shed light on the regulatory mechanisms of basal innate immunity in C. elegans.
Although this study focused on the immunomodulation effects of RJ, we do not completely rule out the antimicrobial function of RJ. We showed that RJ, at our testing concentration, did not kill S. aureus and did not reduce the expression of the sarA gene that encodes the major transcriptional regulator of virulence genes. A previous study reported that both live and heat-inactivated S. aureus could kill C. elegans. 17 We reasoned that RJ would not provide protection against dead S. aureus if it acts through attenuating the bacterial virulence. But we obtained contradictory result; the heat-inactivated S. aureus did not kill the RJ-untreated worms. Hence, we do not know if the SarA-independent virulence genes were affected by RJ.
The discrepancy on killing by the heat-inactivated S. aureus might be because of the bacterial strain difference; the S. aureus used in the early study may produce some heat-stable virulence factors, but further research needs to be conducted regarding this phenomenon.
Findings that RJ shared the anti-aging pathway to improve the innate immunity highlighted its potential for delaying innate immunosenescence. Concurrent requirement of IIS/DAF-16, p38 MAPK pathway, and Wnt pathway on RJ-mediated immunity elevation indicated that RJ modulated C. elegans innate immunity in a systematic way. Given that these pathways are highly conserved from C. elegans to mammals, the RJ-mediated innate immunity promotion in an aged population has practical implications in higher order organisms.
Further human clinical investigations may open up a new avenue for utilizing RJ as an immunomodulatory supplement to retard immunosenescence and promote the innate immune system. This can ultimately help to alleviate some of the burden associated with treating and caring for the elderly who have fallen victim to opportunistic pathogens, especially those present in clinical or long-term care facilities. Preventative measures and treatments are much more sustainable and economical than treating infections after the fact.
Footnotes
Acknowledgments
The RJ powder was provided by Yamada Apiculture Center, Inc., Caenorhabditis elegans strains used in this work were provided by the Caenorhabditis Genetics Center, which is funded by NIH Office of Research Infrastructure Programs (P40 OD010440). The authors thank Zane Williams for helpful discussions and careful editing of the article.
Author Disclosure Statement
No competing interests declared.
Data Availability
All data generated or analyzed during this study are included in this article and the supplementary data file.
Funding Information
This study was supported by Clemson University Creative Inquiry fund (Y.D. and M.C.).
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Table S1
Supplementary Table S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.