
Abstract
Dietary patterns high in fat influence local and systemic oxidative stress through adipose tissue (AT) accrual and increased reactive oxygen species generation. Lycopene, a carotenoid with antioxidant functionality, may mitigate excess oxidative stress, yet the lipophilic nature of this compound may limit its functionality if sequestered by AT. Thus, it is critical to elucidate whether lycopene's efficacy is limited based on adiposity. The purpose of this study was to investigate the influence of lycopene-supplemented normal- and high-fat diets on systemic and AT redox status. Male Sprague-Dawley rats (n = 18) were fed a 30% normal-fat (NFD) or 60% high-fat (HFD) purified diet supplemented with 100 mg of lycopene/day. Body weight and visceral AT mass, as well as serum and AT lycopene, lipid peroxides, and antioxidant capacity (AC), were assessed after 3, 7, and 10 weeks of supplementation. At week 10, AT mass was significantly higher (P = .028) in the HFD group, yet there were no significant differences in serum or AT lycopene concentrations or lipid peroxides between groups. Additionally, AT in the HFD group exhibited significantly greater lipophilic AC (27.6% higher, P = .031). Results suggest that excess adiposity did not negatively influence circulating lycopene, nor did it limit its antioxidant functionality.
Introduction
Obesity is a multifactorial condition characterized by the accumulation of excess adipose tissue (AT) and concomitant rise in oxidative stress and inflammation. 1,2 The resulting disruption in homeostasis fosters an environment conducive to the onset of cardiometabolic disease. 3 Currently, obesity affects 12% of the world's adult population, but its prevalence is estimated to surpass 20% by 2030. 4,5
It is well established that dietary patterns high in fat contribute to local and systemic oxidative stress through accrual of AT with subsequent increased production of reactive oxygen species (ROS). 2,6 The locally generated ROS are transmitted into circulation to influence systemic oxidative stress levels. 7
Lycopene, found most concentrated in tomatoes, tomato products, and watermelons, is a potent lipophilic antioxidant with singlet oxygen quenching abilities. 8,9 As such, it may mitigate excess levels of oxidative stress and disrupt the ensuing dysfunction of a high-fat diet. Research has demonstrated an improvement in systemic redox status resulting from the antioxidant action of lycopene; however, the influence of AT on lycopene's protective mechanisms of action remains underexplored. 10 –12
The relationship between carotenoid supplementation and AT biology warrants investigation as a recent systematic review reported that the body mass index (BMI) may serve as a mediating variable such that serum lycopene was not found to be protective in individuals classified as obese. 13,14 These findings are not limited to lycopene, similar relationships have been established between BMI and other carotenoids, such as β-carotene and lutein, as well as a reported reduction in serum lipophilic antioxidant capacity (AC) for obese individuals. 15 –17 This inverse relationship has commonly been ascribed to AT sequestration of lipophilic compounds from circulation, thus rendering them less bioavailable. 18 Although this hypothesis has been evaluated with some lipid-soluble compounds, such as vitamin D, it has yet to be assessed in the context of lycopene. 19 –21
Current data suggest that lycopene uptake by AT readily occurs through a facilitated process involving the membrane transporter cluster of differentiation 36 (CD36) in adipocytes. 22 Such findings from the molecular investigation of lycopene support the need for additional research evaluating lycopene's potential sequestration and redox at the tissue and systemic levels.
Given lycopene's heightened efficiency for quenching singlet oxygen among other carotenoids, as well as its potential to influence metabolic perturbations of obesity, it is critical to elucidate whether lycopene's efficacy is limited based on adiposity. 23 –25 A relationship of this nature would challenge its proposed functionality in health and disease as a high-risk population, such as obese individuals, may not receive the purported benefits of a dietary lycopene intervention. Thus, the purpose of this pilot study was to compare the influence of lycopene-supplemented diets meeting and exceeding recommendations for fat intake in humans on systemic and AT redox status. It was hypothesized that excess adiposity would reduce circulating lycopene and limit its protective mechanisms of action as an antioxidant.
Materials and Methods
Experimental animals
Male Sprague-Dawley rats (n = 18; 4 weeks of age) (Charles River, Wilmington, MA, USA) with comparable body weights were individually housed under controlled conditions (23 ± 1°C, 12-h light–12-h dark cycle) with free access to standard chow and water during a 3-day acclimation period. 26,27 All procedures were approved by the Institutional Animal Care and Use Committee of the University of Alabama (Tuscaloosa, AL, USA) in accordance with guidelines from the U.S. National Institutes of Health.
Experimental diets
Following acclimation, rats were randomly divided into one of two purified diet groups with differing percentages of fat: 30% normal-fat diet with lycopene (NFD) or 60% high-fat diet with lycopene (HFD) (Research Diets, New Brunswick, NJ, USA) (Table 1). Percentages were selected to model dietary patterns meeting and exceeding recommendations for fat intake in humans. 28 Diets were replaced every evening and food consumption was determined at time of change.
Diet Composition
Diet composition for Sprague-Dawley rats fed normal- or high-fat purified diets. Formulation details are provided in grams (g) and kilocalories (kcal).
Research diets (rodent diet with 30% kcal fat, D08091811).
Research diets (rodent diet with 60% kcal fat, D12492).
Each diet group was supplemented with 100 mg/day of lycopene (LycoVit® dispersion 10%; BASF North America, Florham Park, NJ, USA). This dose was selected to assess the maximum potential for lycopene sequestration without surpassing the no-observed-effect level established in a previous 90-day toxicity study with 1% dietary lycopene supplementation. 29 The amount of lycopene ingested was calculated from daily food consumption.
Study design
At weeks 3, 7, and 10, three rats in each diet group were euthanized through carbon dioxide inhalation and body weight was recorded. Blood was drawn from the vena cava, and following centrifugation, serum was obtained and stored at −80°C until analysis upon study completion. Visceral white AT was harvested with weight recorded and snap-frozen in liquid nitrogen for storage at −80°C until analysis.
Analysis of lycopene
Tissue and serum lycopene extraction was conducted according to a previously validated procedure by Boileau et al. 30 Chromatographic separation was carried out using an ACQUITY ultrahigh-performance liquid chromatography system with a photodiode array detector and ACQUITY BEH Shield RP18 2.1 × 100 mm, 1.7 μm (Waters, Milford, MA), according to a previously described method for fat-soluble micronutrients. 31 Lycopene and β-apo-8′-carotenal were sourced from Sigma-Aldrich (St. Louis, MO), and β-apo-8′-carotenal served as the internal standard. The limit of quantitation for lycopene was 0.039 μM.
Measures of redox status
Oxidative stress
Malondialdehyde (MDA), a product of lipid peroxidation, is a biomarker of oxidative stress. Serum and AT lipid peroxides were quantified according to the thiobarbituric acid-reactive substances assay, as previously described. 32 Results are expressed as mM MDA.
Antioxidant capacity
Serum and AT were deproteinated according to a published method using methanol/acetonitrile/acetone (1:1:1, v/v/v) added to samples in a ratio of 1:4 (v/v). 33 This method allows for detection of small-molecular-weight antioxidants (<6 kDa). AC was measured using the oxygen radical absorbance capacity assay on a FLUOstar Optima plate reader (BMG Labtech). 34 The compound, 2,2-azobis(2-amidinopropane) dihydrochloride, was used as the peroxyl radical generator and Trolox, a water-soluble analog of vitamin E, served as the reference antioxidant standard. Results are expressed as μM Trolox equivalents.
Statistical analyses
As distinction in adiposity between the two groups was integral to the study design, sample size calculations were based upon expected differences in AT accrual from previous literature. 35 Thus, power analysis generated a sample of n = 18 (n = 3 per group at each time point) for 80% power at an alpha level of 0.05. Differences in outcome measures were evaluated by multivariate analysis of variance with dietary intake as the continuous covariate. Partial correlations for each outcome measure between serum and AT were also conducted. Statistical significance was defined as P < .05. All analyses were performed using SPSS, version 25 (SPSS, Inc., Chicago, IL, USA). Results are expressed as mean ± standard deviation.
Results
Dietary intake, lycopene intake, and AT mass
Although energy intake (kcal/day) was not significantly different between groups, rats consuming the NFD had a significantly greater (P < .0001) dietary intake (g/day) and subsequent lycopene intake (mg/day) compared with the HFD group through week 7 (Table 2). Body weight of the HFD group was significantly lower at weeks 3 and 7 (5.3% and 7.0% lower, respectively) (Table 2), yet AT mass did not differ significantly between groups at these time points. The HFD was effective in inducing AT accrual, as evidenced by a significantly greater AT mass at week 10, compared with the NFD group (33.3% higher, P = .028) (Fig. 1).

Accrual of AT. Accrual of visceral AT in Sprague-Dawley rats fed a lycopene-supplemented normal- or high-fat diet. Points represent the mean ± standard deviation. *P < .05 between-group difference. AT, adipose tissue; NFD, normal-fat diet (30%); HFD, high-fat diet (60%).
Body Weight and Dietary, Energy, and Lycopene Daily Intake
Body weight (g), dietary intake (g/day), energy intake (kcal/day), and lycopene (mg/day) daily intake in Sprague-Dawley rats fed a lycopene-supplemented normal- or high-fat diet. Each value represents the mean ± standard deviation.
P < .05 between-group difference.
HFD, high-fat diet (60%); NFD, normal-fat diet (30%).
Serum and AT lycopene concentrations
After 3 weeks of lycopene supplementation, the NFD group exhibited significantly greater serum lycopene concentrations compared with the HFD group (1.2% higher, P = .038) (Fig. 2). These between-group differences were not observed at week 7 or 10. Furthermore, lycopene concentrations in AT were similar between the two groups at each time point. A significant positive correlation between serum and AT lycopene was noted at week 10 (P = .021, r = 0.879).
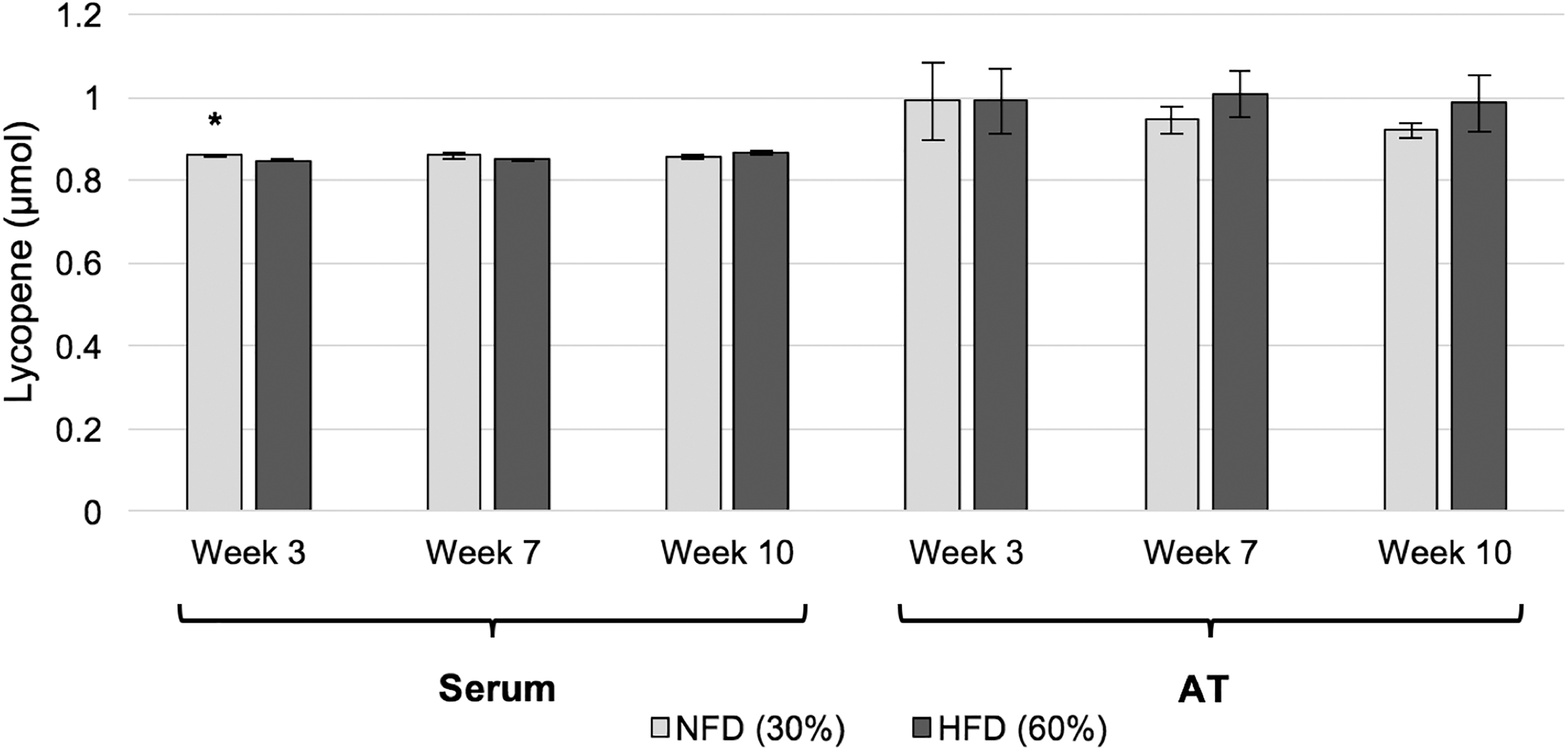
Serum and AT lycopene concentrations. Serum and AT lycopene concentrations in Sprague-Dawley rats fed a lycopene-supplemented normal- or high-fat diet. Bars represent the mean ± standard deviation. *P < .05 between-group difference. AT, adipose tissue; NFD, normal-fat diet (30%); HFD, high-fat diet (60%).
Measures of redox status
No significant differences in serum or AT lipid peroxide concentrations were observed between groups at any time point (Fig. 3). Similarly, the hydrophilic and total AC of serum did not differ significantly between groups throughout the study duration; however, AT in the HFD group exhibited significantly greater lipophilic AC at week 10 (27.6% higher, P = .031) (Fig. 4). A significant positive correlation between serum and AT lipid peroxides was established at week 10 (P = .036, r = 0.841).

Serum and AT lipid peroxide concentrations. Serum and AT lipid peroxide concentrations in Sprague-Dawley rats fed a lycopene-supplemented normal- or high-fat diet. Bars represent the mean ± standard deviation. AT, adipose tissue; NFD, normal-fat diet (30%); HFD, high-fat diet (60%).
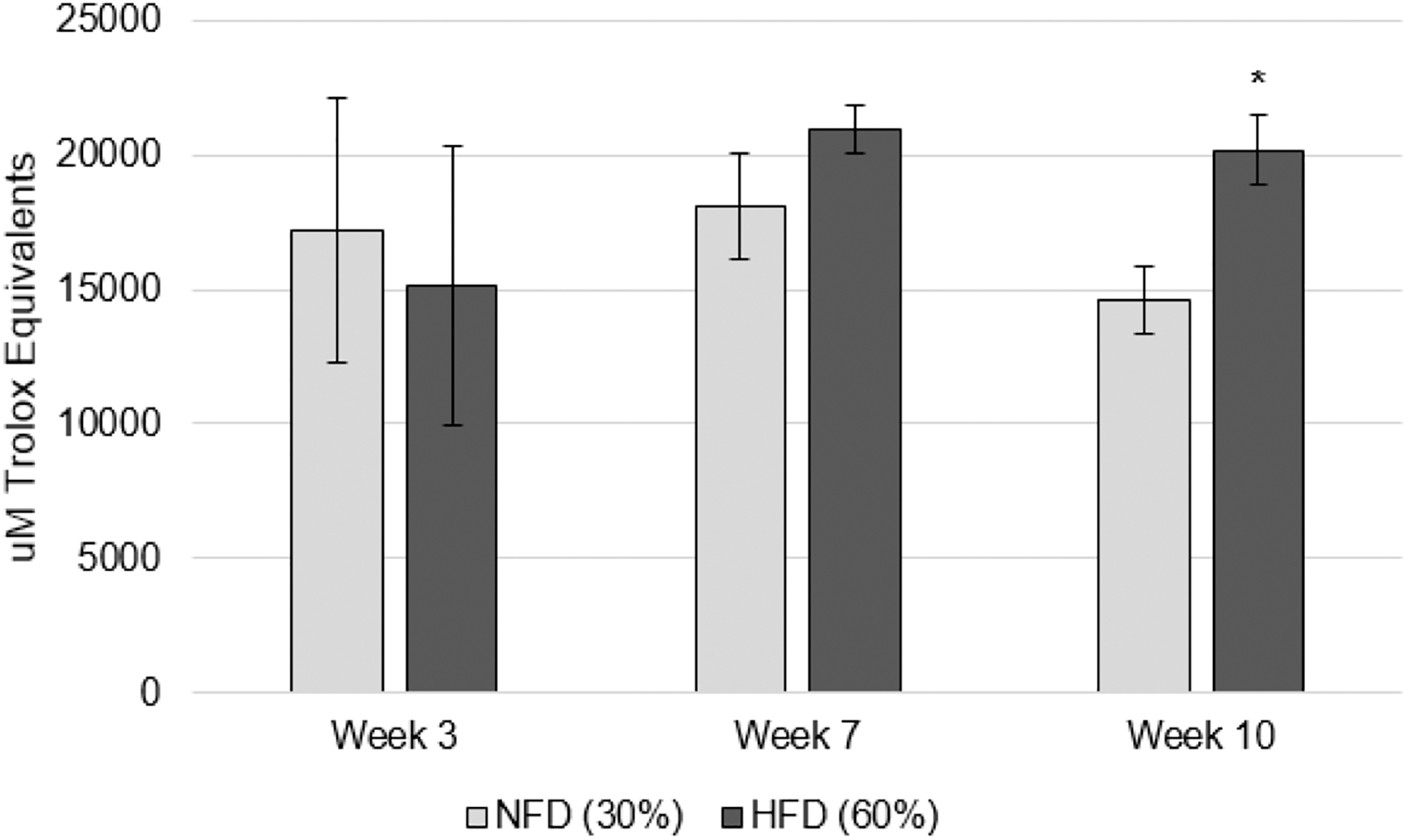
Lipophilic AC of AT. AT lipophilic AC in Sprague-Dawley rats fed a lycopene-supplemented normal- or high-fat diet. Bars represent the mean ± standard deviation. *P < .05 between-group difference. AC, antioxidant capacity; AT, adipose tissue; NFD, normal-fat diet (30%); HFD, high-fat diet (60%).
Discussion
The purpose of this pilot study was to investigate the systemic and AT redox status in Sprague-Dawley rats fed lycopene-supplemented diets meeting and exceeding recommendations for fat intake in humans. Previous studies have reported a reduced serum AC, specifically lipophilic antioxidants, among obese individuals. 16,17,36 This inverse relationship has commonly been attributed to an enhanced uptake of circulating lipophilic compounds by AT. As such, the lipophilic nature of lycopene may limit its functionality under excess adiposity. It was hypothesized that excess adiposity would reduce circulating lycopene and limit its functionality as an antioxidant.
After 10 weeks of supplementation, AT mass was significantly greater in the HFD group, yet serum and AT lycopene concentrations were similar between groups after adjusting for dietary intake. This is in contrast to prior studies evaluating vitamin D and AT, as well as those examining associations between adiposity and circulating carotenoids, in which AT mass negatively influenced the bioavailability of those nutrients. 17,19,20 In the present study, a correlation between serum and AT lycopene was observed at week 10, which supports the function of AT as a dynamic storage pool for lycopene such that in times of high oxidative imbalance, AT may release this compound into circulation. 18,37 Thus, while excess AT may sequester lipophilic compounds, results from this study suggest that sequestration is not occurring at the detriment of circulating lycopene.
Although AT lycopene concentrations were not significantly different between groups, the HFD group did exhibit increased lipophilic AC at week 10. Acknowledging that the contribution of carotenoids to AC in AT is understudied, it is imperative for future research to elucidate the mechanism underlying this difference. However, it is also plausible that these antioxidant findings support AT acting as a reservoir for other dietary lipophilic antioxidants and micronutrients. 18 Nonetheless, if sequestration occurs at a rate proportional to AT, excess storage of these antioxidants may induce a pro-oxidant state, further contributing to redox imbalance within AT and systemically.
Despite this enhanced lipophilic AC observed at week 10, measures of oxidative stress within AT did not differ between the two groups. It may be postulated that the higher lipophilic AC in the HFD group at week 10 influenced AT oxidative stress such that levels were comparable between groups. Furthermore, this local suppression likely conferred systemic benefits in the HFD groups as the expected elevated levels of oxidative stress in serum were not observed. Although additional information is needed to ascertain lycopene's potential contribution to maintenance of AT homeostasis, it should be noted that similar redox results have been obtained by Choi and Seo. 38 The implementation of a 45% HFD in the aforementioned study induced plasma redox imbalance, yet a lycopene intervention restored oxidative stress measures to levels within parameters of the normal-fat diet groups.
In short, the provision of 100 mg/day of dietary lycopene in the context of an HFD appears to influence local redox status to a similar extent as this dose provided in an NFD. As such, its consumption has the potential to provide redox benefits that are not dependent upon levels of adiposity and, consequently, may benefit those in both a normal weight and obese classification. Although extrapolation of average lycopene intake from the current study to a human equivalent dose may not be reasonably attained through dietary sources exclusively, it could potentially be met by oral supplementation. 39 However, the aim of the current study is to investigate if lycopene evades sequestration and maintains antioxidant functionality in circumstances of excess adiposity. Nonetheless, efforts to increase lycopene intake beyond the current estimated intake levels of 5.7–10.5 mg/day are warranted. 40
Taken collectively, results suggest that excess adiposity does not appear to limit the antioxidant functionality of lycopene, nor does it result in a pro-oxidant state within AT. Instead of acting as a reservoir to sequester and compromise circulating levels of lycopene, AT may act as a dynamic storage pool to release lycopene into circulation for the attenuation of systemic oxidative stress. The implications of these tissue- and system-level findings are substantial considering that lycopene may play a role against additional obesity-related complications, including low-grade inflammation and insulin resistance, among others. 41 –44
Through the investigation of lycopene deposition, as well as redox status, within AT, this pilot study advances the current understanding of AT biology and carotenoid supplementation at the tissue and systemic levels, an area noted to be underexplored. 45 Results are strengthened by the use of purified diets in both groups to reduce the potential for confounding variables. 35 Furthermore, this is the first study to evaluate lycopene supplementation in a diet-induced obesity model utilizing a 60% fat diet. While this is not a typical macronutrient distribution, implementing a 60% fat diet represents a model for assessing maximum compound sequestration by AT.
As the primary aim of this pilot study was to investigate if lycopene's deposition and functionality were limited based on adiposity, the study design did not include a group without lycopene supplementation. However, in currently unpublished data from our laboratory with Sprague-Dawley rats consuming a similar purified high-fat diet devoid of lycopene, the lipophilic AC of AT was significantly lower than that of the HFD plus lycopene group in the current study owing to the antioxidant contribution of lycopene or its metabolites.
Despite study strengths, it is not without inherent limitations, such as the use of rats of one gender. Although this may influence generalizability, the male gender was selected to limit hormonal changes as a confounding variable since the female estrous cycle has been reported to affect food consumption. 46
Additionally, the anticipated distinction in adiposity between the diet groups was not significant until week 10. The delayed difference in the current study is likely attributed to a higher percentage of fat in the NFD group (30%) compared with the control diets of other diet-induced obesity studies (10–25%). 43,47 While the NFD composition in the present study was selected to model dietary patterns meeting the recommendation for fat intake in humans, it also resulted in more time required for significant differences in white AT accrual to be observed. Thus, although this pilot study was intentionally designed to represent typical human consumption patterns, an expanded time frame of investigation is warranted to determine whether the long-term accrual of AT with a greater distinction in adiposity between groups surpasses lycopene's ability to exert its protective antioxidant action.
In light of results reported herein, future research investigating gene expression and lycopene-derived oxidation products will provide further insight into lycopene's role in influencing AT and, consequently, systemic redox status.
Footnotes
Acknowledgment
The authors would like to thank BASF North America for their generous donation of lycopene (LycoVit® dispersion 10%) for this study.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.