
Abstract
Wild blueberries rank very high in anthocyanins (ACNs) and phenolic acids (PA) among other berries. Previous work from our group has documented their beneficial effects. In this study, human umbilical vein endothelial cells were used, and ACNs, PAs, and their combination (ACNs:PAs) at concentrations of 0.002, 8, 15, 60, and 120 μg/mL were tested for endothelial tube formation. Treatment with ACNs decreased, while treatment with PAs and ACNs:PAs increased overall endothelial cell tube formation compared to control. Endothelial cells exposed to ACNs downregulated gene expression of AKT1 and endothelial nitric oxide synthase (eNOS), while PAs upregulated AKT1 and vascular endothelial growth factor (VEGF) gene expression. Combination of ACNs:PAs decreased gene expression of AKT1 and eNOS, while protein levels of AKT1 increased. In summary, based on the type of wild blueberry extract, angiogenesis is differentially modulated and is concentration dependent. Further experiments will delineate the mechanism(s) of the differential action of the aforementioned extracts on angiogenesis.
Introduction
Blood vessels are responsible for carrying oxygen to all organs of the human body. 1 New blood vessel formation from preexisting ones is known as the process of angiogenesis. 1,2 Angiogenesis, arteriogenesis, and vasculogenesis are the three different processes of blood vessel formation. 3 Hypoxia-inducible factor (HIF)-1α expression is critical to activate angiogenesis in adults. 3 The de novo formation of blood vessels is known as arteriogenesis. 4,5 The in situ formation of blood vessels from vascular progenitor cells and circulating endothelial progenitor cells is known as vasculogenesis. 6,7 The ability to control pathways of angiogenesis can be used as a potential therapeutic tool. Nearly four decades ago, clinical trials have shown that inhibition of angiogenesis can be a potential therapeutic target for cancer and other diseases. 8 Angiogenesis can have a critical role in diseases where promotion of this process is an important mechanism such as in ischemic disorders, wounds, burns, and others; however, more evidence is required to delineate the molecular mechanisms of angiogenesis. 8
The lowbush blueberry (Vaccinium angustifolium) is one of the richest sources of polyphenols. 9 Wild blueberries (WB) have low concentrations of minerals and antioxidant vitamins but high concentrations of bioactive compounds such as phenolic acids (PAs) and anthocyanins (ACNs), commonly found in food sources such as fruits and vegetables which confer antioxidant activity. 10,11 Wild blueberry ACNs such as delphinidin, malvidin, cyanidin, peonidin, and petunidin are in highest concentration. Growing practices, such as harvesting methods, location, and weather conditions, influence phenolic content. Studies have documented that consumption of WB (V. angustifolium) can reduce inflammation and benefit patients with cardiovascular disease and many other chronic diseases. 9 –16 There is lack of studies on the effect(s) of ACNs and PA fractions from different berries on physiological processes such as angiogenesis, cell migration, and wound healing. 17 –22
During cell proliferation and other important cellular functions such as cell migration, cell growth, and metabolism, protein kinase B (AKT) plays a key role. 23,24 It is a member of AGC kinases, and its three known isoforms of AKT are critical in the cardiovascular system. 24 Upstream regulators of AKT are vascular endothelial growth factor (VEGF), platelet derived growth factor (PDGF), epithelial growth factor, and basic fibroblast growth factor (bFGF). 24 –26 During angiogenesis, AKT regulates the secretion of VEGF; and during endothelial cell migration, which is a critical function of angiogenesis, AKT also regulates VEGF through the AKT-PI3K pathway. 27 –29 Moreover, endothelial nitric oxide synthase (eNOS) which is a key player in angiogenesis and vascular permeability can be directly phosphorylated by AKT. 30
Even though previous studies have documented the antiangiogenic effect of ACNs on angiogenesis, 20,31 –36 evidence on the role of PAs on the above process is scant and conflicting. An in vitro study documented that human umbilical vein endothelial cells (HUVECs) treated with 0.1, 1, and 2 μg/mL of ACNs from black beans inhibited tube formation after cells were induced by VEGF and showed inhibitory effects in a wound healing assay. 37 Furthermore, purple rice fractions were found to inhibit ERK 1/2 and p38 pathways leading to inhibition of cell proliferation and migration with greater impact on VEGF-induced angiogenesis. 31 In addition, the anthocyanin nasunin, isolated from eggplant peelings, was documented to suppress microvessel outgrowth (10–100 μM) in an ex vivo angiogenesis experiment using rat aortic rings 36 but had no effect on tube formation and chemotaxis. 36
Recently, studies in our laboratory documented the differential effects of ACNs and PAs on endothelial cell migration. 38 Significant inhibition of endothelial cell migration in a collective cell migration setup was documented with the ACN fraction, while the PA fraction promoted cell migration by significantly increasing the speed of endothelial cells. Finally, a combination of both ACNs:PAs significantly increased endothelial movement. 39
In this study, we identified the effects of ACN and PA fractions, as well as their combination (ACNs:PAs), from wild blueberry powder and demonstrated the critical role they play in angiogenesis through the PI3K pathway.
The
Materials and Methods
Culture of endothelial cells
Cultures of HUVECs were prepared as described previously in detail. 38 In brief, cells were cultured in F-12K medium supplemented with 10% fetal bovine serum, 1% penicillin–streptomycin solution, heparin 0.1 mg/mL, and endothelial cell growth supplement 0.03 mg/mL. The culture conditions for the cell line was 95% air, 5% carbon dioxide (CO2), 90% relative humidity, and 37° C temperature.
WB powder analysis and extraction of ACNs and PAs
Extraction of ACNs and PAs was performed as described. 38 In brief, WB were provided by Wyman's (Cherryfield, ME) in a freeze-dried powder form (FutureCeuticals, Momence, IL). The following fractions were extracted from the freeze-dried powder: (1) Phenolic-rich fraction (ethyl acetate soluble, containing mainly chlorogenic acid), (2) Anthocyanin-rich fraction (methanol soluble fraction, containing mainly ACNs), and (3) Water soluble fraction. High-performance liquid chromatography (HPLC) method was used as previously reported for the analysis of individual ACNs and PAs in the corresponding fractions. 10
AlamarBlue assay
Cell proliferation and cytotoxicity were prepared as previously described in detail (data not presented). 38 Briefly, the cytotoxicity assay was conducted to determine the optimum concentration and exposure time of ACNs and PAs used for all experiments. A broad range of concentrations (0.001–1000 μg/mL for ACNs and 0.001–500 μg/mL for PAs) were tested.
Angiogenesis assay (tube formation)
Based on previously published results on endothelial cell migration,
38
critical concentrations of ACNs, PAs, and ACNs:PAs used were 60 μg/mL for ACNs, 0.002 and 60 μg/mL for PAs, and 8 μg/mL:8 μg/mL and 60 μg/mL:60 μg/mL for ACNs:PAs. Endothelial cells (1 × 104 cells/well) were plated and cultured on Matrigel (BD Biosciences), applied on an IBIDI μ-slide Angiogenesis plate (Ibidi, Martinsried, Germany), and incubated at 37°C for 30 min for gel construction. With this method, cells were induced to form capillary-like tubes. After exposing the cells to anthocyanin, phenolics, and combinations of both fractions, including a control (untreated cells), their effect on tube formation was photographed using an inverted phase-contrast optical microscope (Nikon, TS100) 4 h after exposure, based on the treatment. The
RT-qPCR
HUVECs were treated with ACNs, PAs, and combination of both bioactive compounds for 2 h. For RNA isolation, purification and reverse transcription commercially available kits (Qiagen) were used. Master mix and primers (Invitrogen) for VEGFA (Hs00900055_m1), NOS3 (Hs01574665_m1), AKT1 (Hs00178289_m1), and GAPDH (Hs99999905_m1) for a final 20 μl polymerase chain reaction (PCR) were used.
Immunoblotting analysis
For the detection of AKT1, VEGF, eNOS, and β-tubulin, total protein was extracted from the cells in RIPA lysis and extraction buffer (Thermo Fisher) supplemented with protease/phosphatase inhibitor cocktail (Cell Signaling, 5872). Low fluorescent polyvinylidene fluoride membranes were blotted with anti-Akt1 (1:1000; Cell Signaling, 2938), anti-Phospho-Akt1 (1:1000; Cell Signaling, 9018), anti-Phospho-Akt1 (1:5000; Abcam, ab81283), anti-VEGF (1 μg/mL; Abcam, ab46154), anti-Phospho-eNOS (1 μg/mL; BD Biosciences, 612392), anti-eNOS (1:1000; Cell Signaling, 9572), and anti-β-Tubulin (1:1000; Cell Signaling, 2128). Secondary specific antibodies for either mouse or rabbit IRDye® 800CW Goat anti-Mouse IgG (1:15,000; Li-COR, 925–32310) and IRDye 800CW Goat anti-Rabbit IgG (1:15,000; Li-COR, 925–32211) were used. The Li-COR Odyssey imaging system was used in all immunoblotting experiments. Moreover, for western blot analysis, VEGFA positive control was used (Abcam, ab55566 and R&D Systems, 293-VE-010).
Statistical analysis
For the tube formation/angiogenesis experiments, one-way analysis of variance (ANOVA) was used. Fisher's least significant difference (LSD) test was used as a post hoc. Each experiment was repeated four times (n = 4), and each independent sample per group was tested four times (n = 4). For the real-time PCR and immunoblotting experiments, a two-tailed Mann–Whitney U-test was performed to compare the control to ACNs, while one-way ANOVA was used for the PAs and combination (ACNs:PAs) groups. Fisher's test was used for post hoc comparisons. For gene expression experiments, the control and each treatment group included 10 replicates (n = 10). Finally, for immunoblotting experiments, the control and each treatment group included four replicates (n = 4). All data in the graphs are expressed as mean ± standard error of the mean, and a P value of <.05 was considered significant.
Results
Wild Blueberry Powder ACN and PA fraction characterization
Analysis of the PA and ACN fractions was conducted by HPLC. 38 The PA fraction was primarily composed of chlorogenic acid (10.23 ± 1.8 mg/mL) with traces of ferulic and caffeic acids. The ACN fraction was composed of malvidin glucosides (26.5%), malvidin galactoside (14.8%), delphinidin glucoside (8.9%), petunidin glucoside (8.2%), and cyanidin glucoside (7.4%). Total ACN concentration was 45.11 ± 0.35 mg/mL.
Angiogenesis assay
The tube formation in vitro assay was first standardized for the HUVEC line. After determining the appropriate cell density and incubation time, experiments were performed in quadruplicate for each treatment. Cells treated with ACNs at 60 μg/mL were not able to form a complete tube network after 4 h of incubation time (Fig. 1).
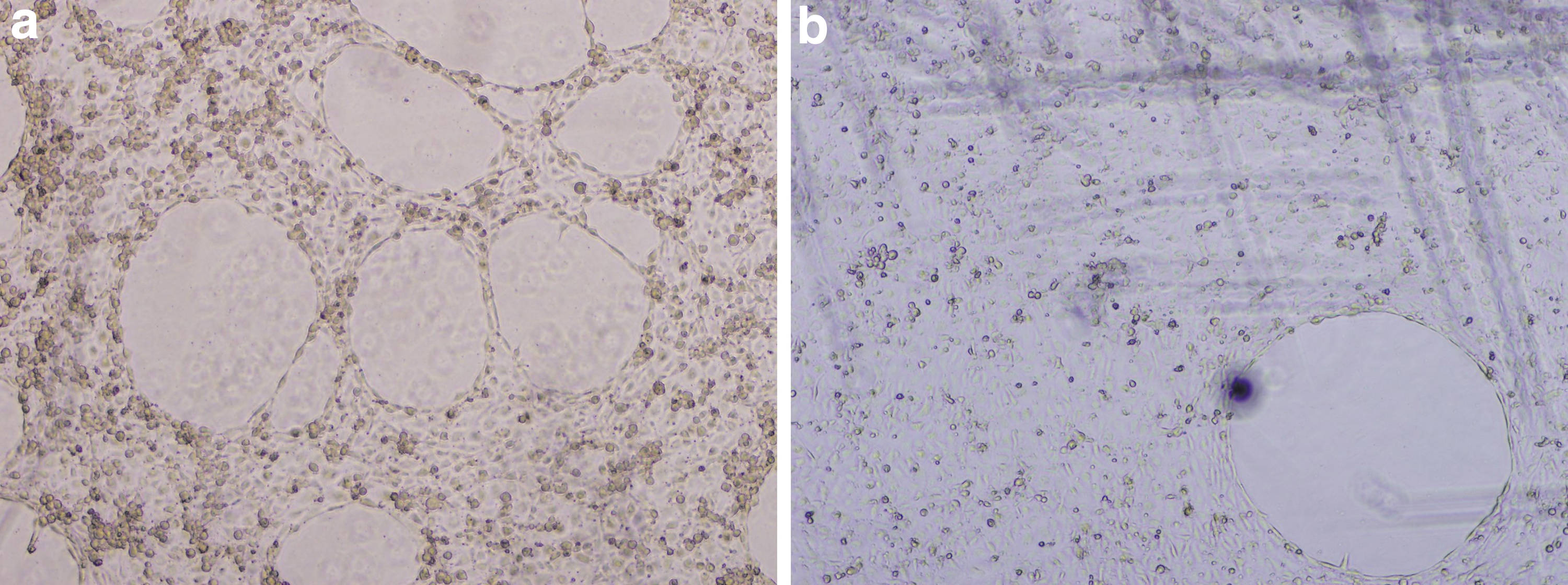
Untreated HUVECs
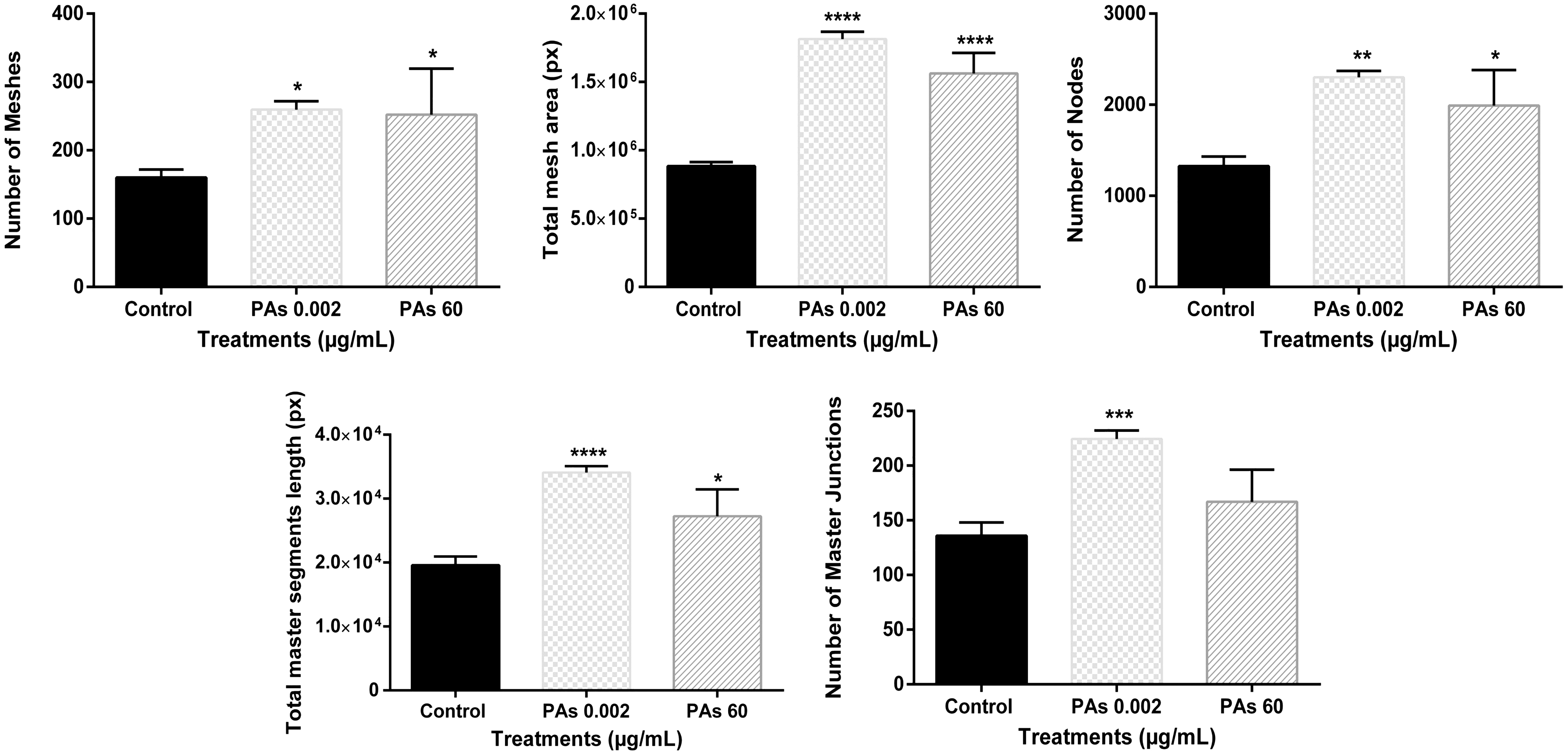
Two parameters from the five tested documented decreased numbers compared to the control (Fig. 2). These were the number of the meshes and the total area of the meshes that are critical and of high importance for the formation of a complete endothelial tube network.

Parameters of endothelial tube formation integrity after cells were treated with ACNs at 60 μg/mL. Treatment with ACNs at 60 μg/mL documented decreased number of meshes (*: P ≤ .05) and area of meshes (**: P ≤ .01) compared to the control.
Cells treated with PAs at 0.002 and 60 μg/mL formed a complete tube network (Fig. 3), which was also quantified (Fig. 4).
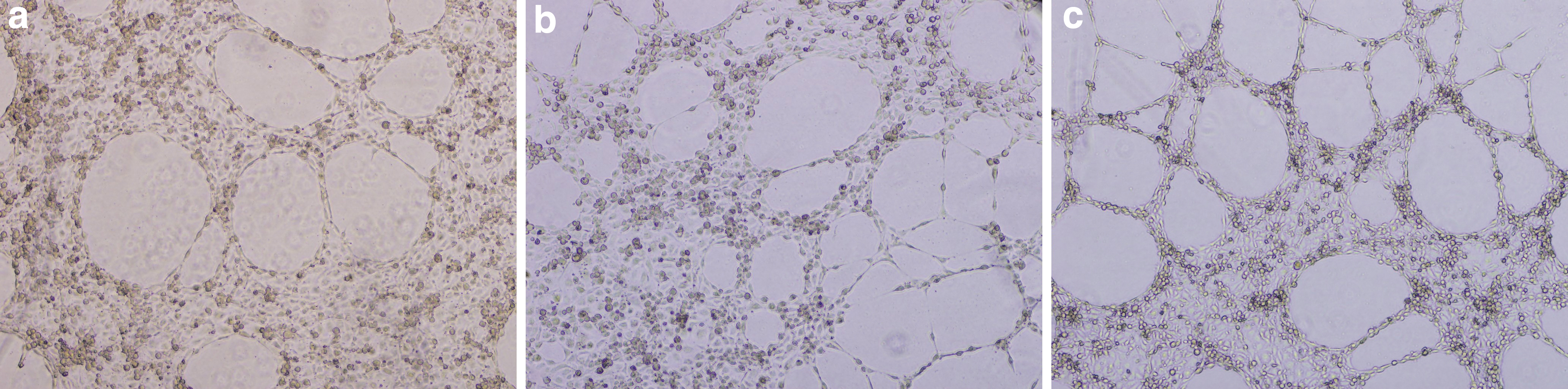
Untreated HUVECs
Parameters of endothelial tube formation integrity after cells were treated with PAs at 0.002 and 60 μg/mL. PAs at 0.002 μg/mL significantly increased all parameters measured for endothelial tube formation compared to the control. PAs at 60 μg/mL significantly increased all the parameters but one (the number of the master junctions) compared to the control. *P ≤ .05, **P ≤ . 01, ***P ≤ .001, ****P ≤ .0001.
PAs at the lowest concentration tested, documented the most significant effect on the endothelial tube network (Fig. 4). Number of meshes (P ≤ .05), area of the meshes (P ≤ .0001), number of nodes (P ≤ .01), total master segment length (P ≤ .0001), and number of master junctions (P ≤ .001) significantly increased compared to the control (Fig. 4). Moreover, exposure to PAs at 60 μg/mL increased significantly number of meshes (P ≤ .05), area of the mesh (P ≤ .0001), number of nodes (P ≤ .05), and total master segment length (P ≤ .05) compared to the control (Fig. 4).
Finally, cells treated with ACNs:PAs formed a complete endothelial network (Fig. 5). ACNs:PAs at 8 μg/mL documented no statistically significant differences compared to the control on any of the five parameters of endothelial tube network integrity (Fig. 6). However, ACNs:PAs at 60 μg/mL increased the total mesh area (P ≤ .0001) and the total master segment length (P ≤ .01) compared to control (Fig. 6).
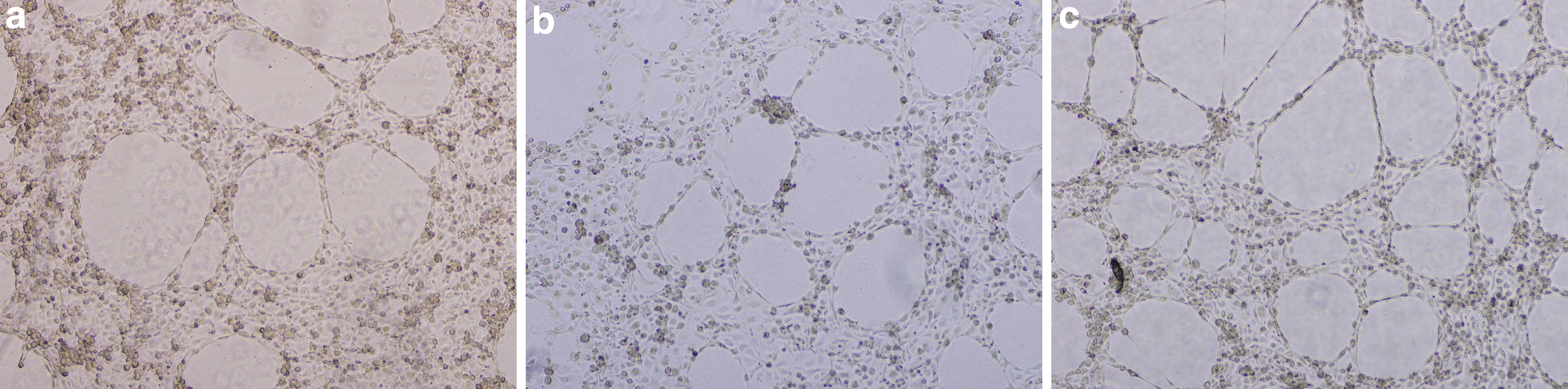
Untreated HUVECs
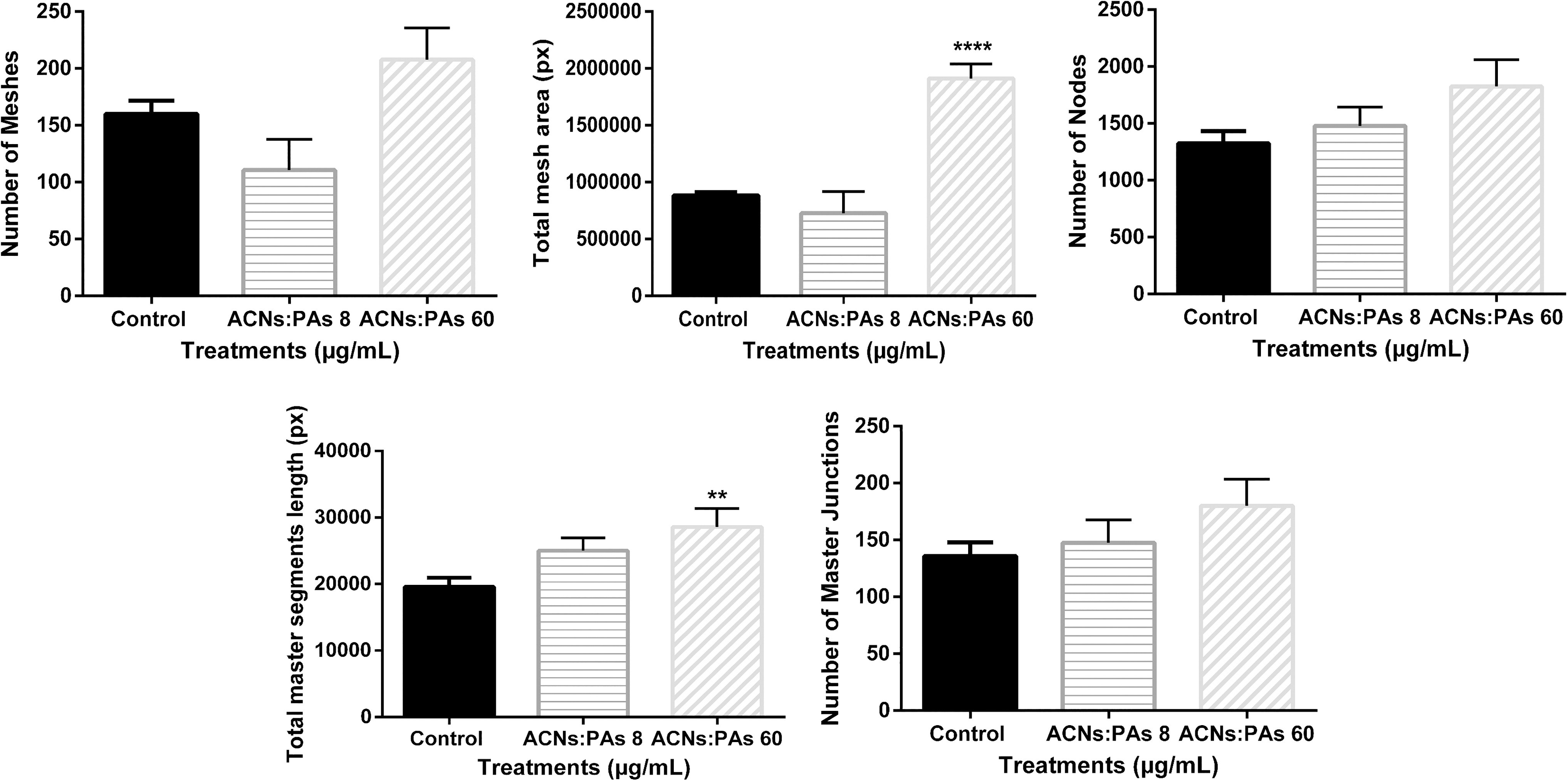
Parameters of endothelial tube formation integrity after cells were treated with a combination of both ACNs and PAs at 8 μg/mL:8 μg/mL and 60 μg/mL:60 μg/mL. ACNs:PAs at 60 μg/mL:60 μg/mL increased the total mesh area (****: P ≤ .0001) and total master segment length (**: P ≤ .01) compared to the control.
Summary from tube formation assay shows ACNs (60 μg/mL), PAs (60 μg/mL), and combination of both fractions (60 μg/mL:60 μg/mL) in Figure 7. Pictures were obtained after 4 h of treatment.

Summary of three different treatments at the same concentration (
RT-qPCR analysis
Gene expression was conducted for AKT1, eNOS, and VEGF (Fig. 8), and results are presented 2 h post treatment. ACNs at 60 μg/mL documented no effect on VEGF gene expression levels. Exposure of cells to PAs at 0.002 μg/mL (P ≤ .01) and 60 μg/mL (P ≤ .05) resulted in an increased gene expression compared to the control. Similar to the effect of PAs, combination at ACNs:PAs 8 μg/mL (P ≤ .05) and ACNs:PAs 60 μg/mL (P ≤ .05) resulted in upregulation of VEGF gene expression levels compared to the control.

Gene expression for VEGF, eNOS, and AKT1. The values are reported as fold change corrected to GAPDH housekeeping gene (*: P ≤ .05) (**: P ≤ .01) (***: P ≤ .001) (****: P ≤ .0001). eNOS, endothelial nitric oxide synthase; VEGF, vascular endothelial growth factor.
At 60 μg/mL (P ≤ .001), ACNs reduced gene expression for eNOS but no significant effect was documented with PAs. ACNs:PAs at 8 μg/mL (P ≤ .0001) and 60 μg/mL (P ≤ .001) resulted in decreased eNOS gene expression levels.
Gene expression of AKT1 at all tested concentrations resulted in a significant effect. Treatment of endothelial cells with ACNs at 60 μg/mL (P ≤ .01) resulted in upregulation of AKT1, and exposure to PAs at 0.002 μg/mL (P ≤ .05) and 60 μg/mL (P ≤ .01) increased AKT1 expression compared to the control. Combination of both fractions ACNs:PAs at 8 μg/mL (P ≤ .001) downregulated AKT1 gene expression, while ACNs:PAs at 60 μg/mL had no effect at 2 h post treatment compared to control.
Immunoblotting analysis
Western blot analysis documented significant increases in AKT1 (Fig. 9), 2 h post treatment with PAs at 0.002 μg/mL (P ≤ .01) and ACNs:PAs at 60 μg/mL (P ≤ .001). No significant differences were detected in VEGF or eNOS protein levels with exposure to the above fractions.

Western blot analysis for AKT1. (**: P ≤ .01) (***: P ≤ .001).
Discussion
In this study we examined the effects of different fractions from WB on endothelial in vitro tube formation, protein synthesis, and gene expression of AKT1, eNOS, and VEGF. This novel study documented that different fractions extracted from WB at specific concentrations have differential impacts on the integrity of endothelial network development.
In previous work conducted by our group we documented that ACNs are able to inhibit cell migration while PAs and combination of ACNs:PAs enhance the speed of endothelial cell migration even at low concentrations (0.002 μg/mL). 38 This is highly associated with angiogenesis.
In this study we report the effect of ACN and PA fractions and their combination on angiogenesis through the tube formation assay. This particular assay is simple, rapid, reliable, can generate quantitative results, and is overall better than any other in vitro assay to assess angiogenic regulators. 42 –44 HUVECs treated with ACNs at 60 μg/mL failed to form endothelial cell network by significantly decreasing the number of meshes (P ≤ .05) and the total area of the mesh (P ≤ .01) compared to the control. In addition, and in support of our observations above, ACNs were found to inhibit AKT1 (P ≤ .01) and eNOS (P ≤ .001) gene expression 2 h post treatment. AKT1 regulates the secretion of VEGF during cell migration, which is a critical cellular process during angiogenesis. 27 –29 Moreover, AKT1 can activate eNOS by direct phosphorylation, a pivotal step during angiogenesis and vascular permeability. 30
Thus, our results documented inhibition of angiogenesis after endothelial cell exposure to the ACN fraction. This finding agrees with most of previous studies 20,31 –35 using commercially available ACNs found to be in high concentrations in food/berry sources. An in vitro study documented that HUVECs treated with 0.1, 1, and 2 μg/mL of ACNs from black beans inhibited tube formation after cells were induced by VEGF. 37 HUVECs were not able to develop a complete endothelial network, and they showed inhibitory effect in a wound healing assay. Purple rice fractions were documented to inhibit ERK 1/2 and p38 pathways by inhibiting cell proliferation and migration with greater impact on VEGF-induced angiogenesis. 31 Tube formation experiments evaluated by measuring the tube area, length, joint, and path documented an inhibitory effect of purple rice fractions with concentrations from 3 to 30 μg/mL. 31
Other in vitro studies confirmed the antiangiogenic properties and antioxidant activities of bilberry main anthocyanidins (delphinidin, cyanidin, and malvidin) on tube formation assay using HUVECs and documented inhibition of the tube formation network when cells were treated with concentrations from 1 to 3 μM. 45 Among different (commercially available) ACNs tested (cyanidin, delphinidin, malvidin, pelargonidin, peonidin, and petunidin), delphinidin had the strongest antiangiogenic effect. 33 Delphinidin inhibited VEGF-dependent tyrosine phosphorylation of VEGFR-2 in a dose- and time-dependent manner. 33 In addition, the anthocyanin nasunin, isolated from eggplant peelings, was documented to suppress microvessel outgrowth (10–100 μM) in an ex vivo angiogenesis experiment using rat aortic rings. 36
For PAs, we documented a different pattern from ACNs. Exposure of HUVECs to 0.002 μg/mL PAs was able to form an integral cell network, and all parameters of angiogenesis (number of meshes, total area of the mesh, number of nodes, total master segment length, and number of master junctions) were significantly increased. Same results were documented at 60 μg/mL, only without the significant effect on the number of the master junctions. Gene expression of AKT1 and VEGF increased with PAs at 0.002 and 60 μg/mL, and protein levels of AKT1 also increased at 0.002 μg/mL (P ≤ .01). Finally, ACNs:PAs upregulated VEGF gene expression significantly 2 h post treatment (P ≤ .05, P ≤ .01).
Limited and contradicting results exist on the role of PAs on angiogenesis. Some studies have documented proangiogenic and others antiangiogenic effects. Endothelial cells (HUVECs) treated with chlorogenic acid (10 μM) 46 documented significant inhibition of hypoxia-induced network formation, reduction of HUVEC mobility, and inhibition of cell invasion. Yet, ferulic acid (10−6–10−4 M) from commercial sources, similar to chlorogenic acid (under the same classification), induced cell migration and tube formation of HUVECs 47 by upregulating VEGF and PDGF pathways. Increased percentage of neovascularization was observed with ferulic acid (10−6–10−5 M) using chick chorioallantoic membrane (CAM) assay. 47 In contrast, p-coumaric acid (0.5 and 1.0 mM) from commercial sources suppressed sprouting of endothelial cells in rat aortic rings and inhibited tube formation and migration of ECV304 cells. 48 Moreover, other in vitro study using ellagic acid (2.5–20 μM) documented inhibition of angiogenesis in HUVECs through inhibition of VEGFR-2. 49 Hence, it has been documented that both anti- and proangiogenic properties of PAs, either as individual compounds or as extracts, can have a possible involvement of VEGF, PDGF, and HIF-1a signaling pathways. The majority of studies tested concentrations far above the physiological levels in the blood stream and used single phenolic compounds. In our wild blueberry extracted PA fraction, the major PA found was chlorogenic acid, with caffeic and ellagic acids found in trace amounts.
Finally exposing the HUVECs to a combination of ACN and PA extracts resulted in increased total mesh area and total master segment length when cells were treated with 60 μg/mL:60 μg/mL with increased gene expression of VEGF and decreased gene expression for AKT1 and eNOS at 2 h post treatment. Even though gene expression of AKT1 decreased compared to the control, increased protein synthesis was documented at 2 h post treatment at 60 μg/mL:60 μg/mL. To our knowledge, combination of both fractions on angiogenesis, evaluating the effect on AKT1, eNOS, and VEGF, has not been reported in the past.
These novel findings are supported by recently published data from our laboratory of similar effects of wild blueberry ACNs, PAs, and ACNs:PAs on endothelial cell migration. 38 ACNs (60 μg/mL) were documented to have an inhibitory effect on cell migration, while PAs (0.002, 60, and 120 μg/mL) and ACNs:PAs (8 μg/mL:8 μg/mL and 60 μg/mL:60 μg/mL) induced endothelial cell migration. Moreover, gene expression of genes (RHOA and RAC1) critical for endothelial cell migration increased rapidly 2 h post treatment. Protein levels of RHOA and RAC1 were also affected 2 h post treatment with PAs and ACNs:PAs; however, no significant change was documented with ACN treatment. 38 Since cell migration is a precursor of angiogenesis the findings in the present study are in harmony with our previous work. 38
The present study examined for the first time the effects of wild blueberry fractions on HUVECs after acute exposure. We examined a range of low and high concentrations of ACNs and PAs, as well as a combination of both, on cell cytotoxicity, angiogenesis, gene expression, and protein synthesis of AKT1, eNOS, and VEGF. Some of the concentrations used in our experiments (ACNs, PAs, and combination at 0.002 and 8 μg/mL) were similar to the physiologically reported concentrations that have been detected in the blood circulation. 50 Scientists have documented that polyphenolic aglycones in plants can be found in concentrations ranging from low μM to mM. 50 ACNs can be found after consumption of food in the plasma at 274 nM. 51
The stability and structural and chemical properties of the evaluated fractions in the present study can possibly explain the differential effects observed on several angiogenesis parameters and gene expression of molecules directly related to formation of endothelial tube networks in vitro. Moreover, degree of glycosylation, oxidation, cell receptor availability, formation of polymeric molecules, and existence of stereoisomers can also be considered as possible factors of differential effects. 50
Finally, our results suggest that ACNs, PAs, and combinations of both have differential effects on different parameters on the endothelial tube network, gene expression, and protein synthesis of important molecules such as AKT1, eNOS, and VEGF. Preclinical studies are underway in our laboratory to validate the results we observed with our in vitro system with an in vivo model. Angiogenesis is a complex phenomenon, which plays a key role in health and disease. Unraveling the significant effects of wild blueberry extracts on signaling pathways that regulate angiogenesis could be of critical importance for possible novel clinical treatment approaches.
Footnotes
Acknowledgment
The authors thank FutureCeuticals (Momence, IL) for providing the wild blueberries.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This project was supported by the USDA National Institute of Food and Agriculture, Multistate Project No. ME0- 31910 through the Maine Agricultural & Forest Experiment Station. Maine Agricultural and Forest Experiment Station Publication No. 3752 and by the Wild Blueberry Association of North America.