
Abstract
Dysmenorrhea is painful menstrual periods, which affects 25% of women within reproductive age and has a prevalence of 67.2–90.0%. Current treatment has several adverse effects and can be ineffective once the pain is initiated. Thymus vulgaris traditionally used for pain management was investigated in this study for its activity on uterine contraction in the nonpregnant uterus, as a parameter for dysmenorrhea. The dried leaves of T. vulgaris were macerated in water, and the resulting aqueous extract was investigated on the isolated mouse uterus. Parameters investigated included spontaneous contractions, oxytocin-induced contractions, and high potassium chloride (KCl; 80 mM)-induced tonic contractions. Mass spectrometric analysis of the thyme extract was also performed using liquid chromatography–high-resolution Fourier Transform mass spectrometry. Thyme extract inhibited the amplitude and frequency of spontaneous and oxytocin-induced uterine contractions. It also inhibited KCl-induced tonic contractions. The activities observed suggest that T. vulgaris inhibits uterine contractions through blockade of extracellular voltage-gated calcium channels. Secondary metabolites detected included compounds belonging to chlorogenic phytochemical class and flavonoids, which are known to have activities on extracellular calcium blockade. This study has shown that aqueous T. vulgaris extract, also known as thyme, inhibits contractions of the nonpregnant uterus and can be a lead plant in the drug discovery process for the management of dysmenorrhea.
INTRODUCTION
There are smooth muscle cells in the uterus, which are responsible for contraction and relaxation. 1 This contraction of the uterus has a myogenic origin and occurs even when the uterus is denervated. 2 The nonpregnant uterus experiences contractions that may be spontaneous or agonist induced. These contractions of the nonpregnant uterus are necessary for sperm transport to the oviduct from the cervix, for movement of the ovum to the uterine cavity, and to expel sloughed materials in the early follicular phase in women. 3 Contractions of the nonpregnant uterus also assist in implantation. 4 Excessive or abnormal contractility of the nonpregnant uterus can cause pain often referred to as dysmenorrhea, and in the pregnant uterus, contractions can lead to preterm labor or abortion. 2,5
Dysmenorrhea is painful menstrual periods 6 that affects women of reproductive ages. It affects about 25% of women in reproductive age, with a prevalence of 67.2–90.0%. 7 Dysmenorrhea is classified into primary and secondary dysmenorrhea. 8 Primary dysmenorrhea occurs due to an increase in prostaglandin release, particularly PGF2α , resulting in vascular constriction and myometrial contraction. Blood flow to the endometrium becomes insufficient, and the women experience pain. 9 Secondary dysmenorrhea on the other hand occurs due to gynecological pathologies such as uterine leiomyoma and other uterine pathologies. 9
Nonsteroidal anti-inflammatory drugs are currently utilized for clinical management of dysmenorrhea, 10 but are not without their adverse effects, which include hepatic, renal, as well as cardiovascular disturbances. 11 In the search for new and more effective therapies, Thymus vulgaris was investigated in this study.
T. vulgaris, commonly known in culinary as thyme, is an aromatic herb known for the distinctive flavor it adds to foods. 12 It is a flowering plant that belongs to the family Lamiaceae and is indigenous to Southern Europe, 13 but widely distributed around the world. It is a small perennial shrub that grows up to 15–30 cm in height. 13
Thyme is used as a culinary herb and as herbal medicine. 14 The flowers are pale violet and are two-lipped shaped, occurring about 5 mm long with a hairy glandular calyx. 14 The leaves range from oval to rectangular shapes. 13 Thyme also consists of numerous beneficial activities, which include, but are not limited to, antioxidative, antimicrobial, carminative, and antiseptic effects. 15 It has also been used traditionally for treating pain. 14 T. vulgaris was therefore investigated on nonpregnant uterine contractions, to evaluate its potential in the management of dysmenorrhea.
MATERIALS AND METHODS
Plant extract preparation
Dried thyme leaves were purchased from a local market in Benin City, Edo State, Nigeria. The dried thyme leaves (500 g) were powdered using an electric blender and macerated in water (2.5 L) at room temperature (30°C ± 2°C) for 24 h. During the period of maceration, the mixture was continuously stirred. After 24 h, the extract was filtered and concentrated in an oven set at 40°C. The aqueous leaf extract of Thymus vulgaris (TL) was stored in airtight containers at 4°C until needed.
Animals
Nonpregnant mature female albino mice weighing between 20–30 g were utilized in this study. The mice were obtained from the Animal Unit of the Department of Pharmacology and Toxicology, Faculty of Pharmacy, University of Benin, Edo State, Nigeria. They were housed in standard cages at an environmentally controlled room temperature of ∼27°C ± 5°C and lighting conditions. Ethical consent was obtained before the start of the experiments from the Faculty of Pharmacy Ethics Committee, University of Benin, Nigeria (EC/FP/016/04). Handling of animals was according to standards of the public health service policy on humane care and use of laboratory animals. 16,17 The animals were fed a standard animal pellet diet and had access to clean water ad libitum.
Contractility studies
Tissue preparation
Diethylstilboestrol (DES; 1.0 mg/kg p.o.), constituted in Tween 80 and distilled water (1:1), was administered to each mouse, 24 h before the experiments.
18
On the experiment day, vaginal lavage was obtained with the aid of a pipette (0.1 mm tip). The lavage was placed on a clean glass slide, dried at room temperature, and fixed with ethanol. The slide was subsequently stained with a drop of Gentian violet and observed under a microscope to ascertain the estrous cycle stage.
5,19
Mice found to be in proestrus, and estrous stages of the estrous cycle were selected and euthanized by cervical dislocation. The uterine horns were rapidly, but carefully excised and immediately placed into warmed and aerated De Jalon's physiological salt solution in a Petri dish. The composition of the physiological salt solution (PSS) in M was as follows: NaCl 154.00, NaHCO3 5.95,
Experimental procedures
On mounting in the organ baths, the uterine tissues were equilibrated under resting tensions of 4.90 mN for 30–45 min or until regular contractions were achieved. The frequency and force of uterine contractions occurring in the longitudinal muscle layers of the uterus 20 were recorded through connection to a 7003E-isometric force transducer (Ugo Basile, Varese, Italy) connected to a digital recorder 17400 data capsule consisting of an inbuilt bridge amplifier (Ugo Basile).
Experiment on the effect of TL on spontaneous uterine contraction
The activity of TL on the spontaneous contraction of the uterine smooth muscle was investigated. 21 This was achieved through cumulative additions of TL (0.0003–1.3332 mg/mL) to the isolated uterus which showed regular spontaneous contractions. The investigation utilized 3-min contact time of TL with the uterine tissue for each concentration of TL. 22
Experiment on the effect of TL on oxytocin-induced uterine contraction
The activity of TL on oxytocin (OT)-induced uterine contraction was also investigated. 21 TL (0.0012–1.3332 mg/mL) was added cumulatively to the isolated uterus in the presence of OT (60.00 pg/mL). The concentrations of TL used in this protocol utilized median to maximal contractions derived from TL's activity on the spontaneous uterine contraction studies. The investigation again utilized a contact time of 3 min with TL and the uterine tissue for each concentration.
Experiment on the effect of TL on high potassium chloride-induced uterine contractility
To assess the activity of TL on voltage-gated Ca2+ channels, TL was investigated in the presence of high potassium chloride (KCl; 80 mM). KCl (80 mM) was added to the bath containing the uterine tissues for 5 min, and without washing, the cumulative addition of TL (0.014 16.32 μg/mL) was performed and responses recorded. Lack of sufficient extract quantities at this stage prevented exact replication of concentrations used in previous protocols.
Liquid chromatography–high-resolution Fourier Transform mass spectrometry identification of constituents in TL
Mass spectrometric analysis using liquid chromatography–high-resolution Fourier Transform mass spectrometry (LC-HRFTMS) analysis was performed (DIONEX, Sunnyvale, CA, USA). The Dionex Ultimate-3000 was coupled to a ThermoScientific Exactive Orbitrap system (Thermo Fisher Scientific [Bremen] GmbH, Bremen, Germany). The ACE column used was of the dimensions C18 75 × 3.0 mm (Hichrom Ltd., Reading, United Kingdom). The elution flow rate was 300 μL/min with water (A) and acetonitrile (B) in formic acid (0.1%). A gradient flow beginning at B (10%) and gradually increasing to B (100%) within 30 min was utilized. The mobile phase was kept at 100% B for 5 min, followed by equilibration of the column with B (10%). The ProteoWizard was used to slide data files into positive and negative datasets, 23 and MZmine 2.10 was used for data mining. 24,25 The centroid mass detector enabled peak detection at a noise level of 1000. Peak lists were generated from the mass lists using the chromatogram builder, with the following parameters: minimum period of 0.2 min, a minimum height of 10,000, and mass to charge ratio (m/z) tolerance of 0.0001 m/z or 5 ppm.
Local minimum search algorithm enabled chromatogram deconvolution with the following parameters: threshold (90%), minimum relative height (5%), minimum absolute height (10,000), minimum retention time (RT) range (0.4 min), peak duration range (0.2–5.0 min), and a minimum ratio of peak top/edge (2). De-isotoping of peak list was achieved using the isotopic peaks grouper at an m/z tolerance of 0.001 m/z or 5 ppm, RT tolerance of 0.1 min (absolute), and a maximum charge set at 2. The most intense isotope was taken as the representative isotope. The alignment function merged the peak lists, and the weight for m/z and RT was 20, while the RT tolerance was 5%. Peak finder gap filled the aligned peak lists, with an intensity tolerance of 1% and RT tolerance of 0.5 min (absolute). 26
Adducts were identified, together with other complexes that may have formed. The chemical formulae of the peaks were predicted using the formula prediction tool developed by MZmine. Hits from the database were accessed using ChemBioFinder version 13 (PerkinElmer Informatics, Cambridge, United Kingdom).
Data analysis
GraphPad Prism (version 7.03; GraphPad Software, Inc., San Diego, CA, USA) was used to analyze data. The amplitude (force) and the frequency of contractions were computed as means within the 3-min contact time. Data were obtained as percentages of control applications (control = 100% or contractions in the absence of extract) where necessary and variations in force (amplitude) or frequency were analyzed with respect to control (100%). All data shown are denoted as mean ± standard error of the mean, and n represents the number of mice used for each experimental protocol. Significance was evaluated using appropriate t-tests for the OT and potassium chloride data, while one-way analysis of variance with Dunnett's post hoc was used for the spontaneous contraction data. P values ≤.05 were taken as the least significance.
Datasets with sufficient data points were used to compute mean log concentration–response curves and analyzed by fitting data to a variable slope logistic equation, using the following equation values: Y = Bottom + (TopBottom)/[1 + 10^((LogIC50-X) × HillSlope)], where Y represents response initiating at the bottom and continuing to the Top in sigmoid shape, X represents the logarithm of the concentration, and IC50 represents the concentration producing responses halfway between Top and Bottom.
RESULTS
The activity of TL on spontaneous uterine contraction
TL inhibited the spontaneous uterine contractions of the nonpregnant uterus in a concentration-dependent manner (Fig. 1A). This was evident in the marked inhibition of the amplitude (Fig. 1B) and frequency (Fig. 1C) of the spontaneous uterine contraction. The higher concentrations of TL were more effective in inhibiting spontaneous uterine contractions (Fig. 1).

Activity of TL on spontaneous uterine contractions.
The activity of TL on OT-induced uterine contraction
TL produced inhibition of OT-induced uterine contraction (Fig. 2A). TL inhibited the amplitude of OT-induced contraction with TL (1333.2 μg/mL), producing significant inhibition (P < .001) of OT (Fig. 2B). TL also produced significant inhibition (P < .05; .01; .001) at all concentrations used on the frequency of OT-induced contractions (Fig. 2C).
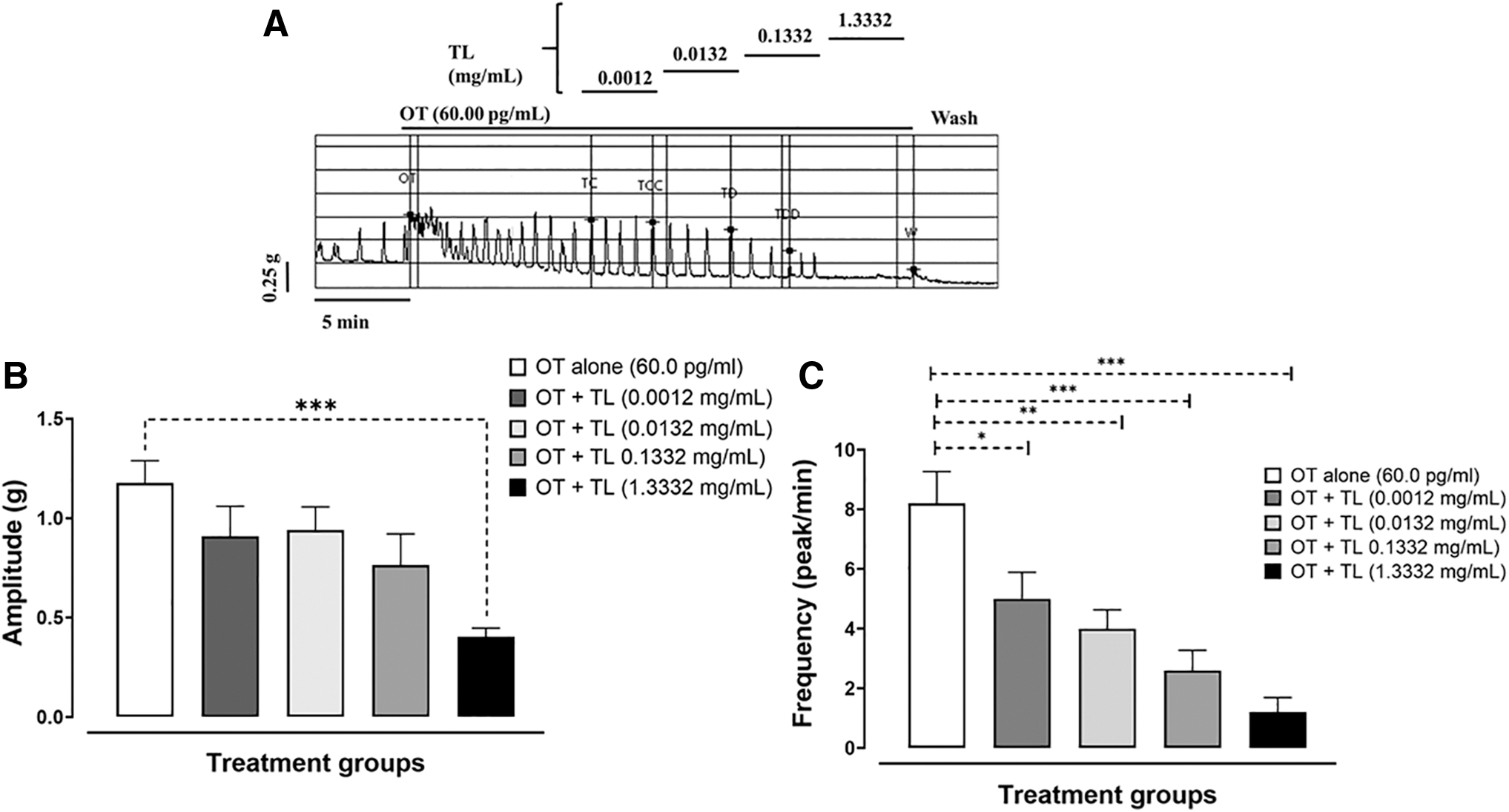
Activity of TL on OT-induced uterine contraction.
The activity of TL on high KCl-induced uterine contractions
TL produced minimal inhibition of high KCl-induced tonic contractions (Fig. 3A). The inhibition induced by TL was evident at 1.632 and 16.332 μg/mL (Fig. 3B).

Activity of TL on high KCl-induced uterine contraction.
Identified secondary metabolites in TL
The LC-HRFTMS results and database search (using Dictionary of Natural Products) enabled the detection of 18 significant compounds (Fig. 4). The identified compounds belonged to a diverse range of phytochemical classes, including polyols, flavonoids, flavones, plant lipids, and triterpenoids, with the majority of detected compounds belonging to the flavonoid class (Table 1) and ionizing in the negative mode (Fig. 5).
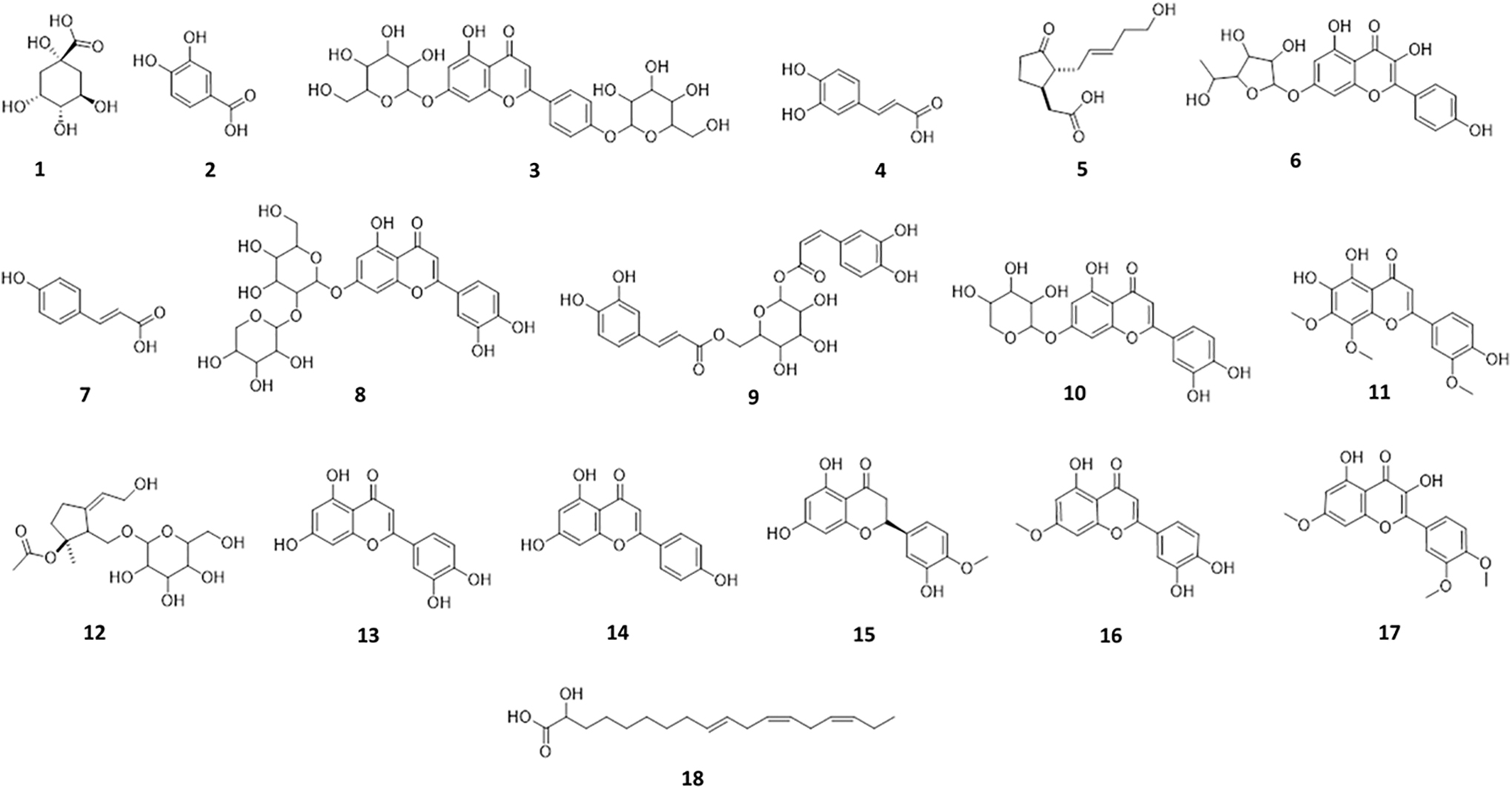
Secondary metabolites

Total ion chromatogram for TL in negative ionization modes showing identified metabolites
The Identified Compounds in Thymus vulgaris
DBE, double bond equivalence; MF, molecular formula; m/z, mass to charge ratio; RT, retention time.
DISCUSSION
T. vulgaris inhibited the uterine contraction of the nonpregnant uterus in this study. There is increased uterine contraction frequency during the follicular phase (proestrus/estrus in mice) of the reproductive cycle in women, which decreases by the luteal phase (metestrus/diestrus in mice) of the reproductive cycle. 2 The reverse, however, occurs with amplitude (force) of contractions, which increases in the luteal phase and decreases during menstruation. 4 Mice in proestrus and estrus phases were used in this study, and these stages are expected to exhibit a high amplitude of contractions, and this will also enable the effect of the extract being investigated to be observed. TL decreased the spontaneous uterine contractions in this study, and this decrease was more pronounced at the higher concentrations of TL. Spontaneous contractions are due to Ca2+ influx 27 mediated through prostaglandin involvement. 28 The sources of Ca2+ include both extracellular and intracellular sources. 29 That TL inhibited spontaneous contraction suggests TL may inhibit Ca2+ influx.
TL also inhibited OT-induced contraction. OT is a potent uterine contraction agonist. When OT interacts with its receptor in the uterine smooth muscle, the phospholipase C system is activated, which subsequently results in Ca2+ mobilization. 30 Recently, other mechanisms have been shown to be activated by OT, which also results in Ca2+ mobilization, and these include the interaction of OT with sphingosine pathway, nicotinamide adenine dinucleotide phosphate pathway, and gamma-aminobutyric acid pathways. 31 Invariably Ca2+ is mobilized in OT-stimulated contraction. That TL inhibited OT-induced uterine contraction also supports that TL may have inhibited Ca2+ mobilization.
Further investigation in this study showed that TL inhibited high KCl-induced uterine contraction. High KCl activates and opens L-type voltage-gated calcium channels (VGCCs). 32,33 Opening of VGCCs by high KCl causes sustained depolarization, 34 seen in this study as sustained tonic contractions. This, therefore, implies that the sole source of high KCl-induced contraction is from extracellular Ca2+. Extracellular Ca2+ activates Ca2+ intracellular release in OT-induced contraction. VGCCs are necessary for the phasic spontaneous and OT-induced contractions, as well as for high KCl-induced tonic contractions. This, therefore, suggests that TL inhibits extracellularly mediated Ca2+ release.
Mass spectrometric studies enabled the identification of secondary metabolites in TL. The probable involvement of the identified metabolites with the activity of TL on the uterus is described. Quinic acid (1,3,4,5-tetrahydroxy cyclohexane carboxylic acid) belongs to alicyclic organic acids or the chlorogenic acid group found in higher plants. 35 The compound quinic acid exists in high amounts in tea, 36 coffee, 37 and some fruit plants. 38 Other chlorogenic compounds found in TL include caffeic acid, protocatechuic acid (3,4-dihydroxybenzoic acid), 39 and p-hydroxycinnamic acid. In plants, quinic acids act as defense metabolites 39 and are the most dominant compounds of the chlorogenic acids. 39 They also have antioxidant activity; however, they achieve low peak plasma concentrations on consumption, and their antioxidant activities are therefore low compared to antioxidants such as ascorbic acid or alpha-tocopherol. 40 On consumption, caffeic acid is converted to ferulic acid and isoferulic acid. Ferulic acid, together with protocatechuic acid, exerts anti-inflammatory effects in vein endothelial cells. 41 Protocatechuic acid modulates IL-6 and vascular adhesion molecule-1, which are vital inflammatory mediators. 40
A caffeic acid derivative, dihydrocaffeic acid, reduces the secretion of the proinflammatory cytokines, TNF-α, IL-1β, and IL-6. 42 Caffeic acid and ferulic acid also inhibit platelet activation. 43 While there have been no direct studies on chlorogenic acids on uterine smooth muscle activity, it is known that proinflammatory cytokines promote smooth muscle contraction 44 and inhibition of these cytokines by these chlorogenic acids found in TL can promote the activity of TL on cytokine-mediated smooth muscle contractility. Proinflammatory cytokines are also involved in dysmenorrhea in the nonpregnant uterus 45 and are released through activated macrophages, 46 which then stimulate prostaglandin release leading to excessive uterine contractility. 47 The activity of the TL compounds on cytokines may, therefore, contribute to the activity of TL in this study.
The flavonoid apigenin chemically known as 4′,5,7-trihydroxyflavone, 48,49 as well as other apigenin derivatives, was also identified in TL. Apigenin is a natural flavonoid found in plants and is often linked to glucose moieties through hydroxyl groups (O-glycosides) or directly to carbon (C-glycosides). 50,51 Apigenin makes up one of the five most common flavonoids 48 and is metabolized to luteolin, 39 which was also identified in TL in this study. Apigenin has been reported to have several biological activities, some of which include anti-inflammatory 52,53 and estrogenic activities. 54 Apigenin is a weak phytoestrogen and binds weakly to the estrogen receptors ERα and ERβ, with a higher affinity for ERβ. 55 It also appears to block progesterone activity 56,57 independent of the estrogen receptor 58 and has been reported to interfere directly with the signaling of progesterone. 57,59
Apigenin is also known to decrease implantation sites in mouse models. 60 The apigenin metabolite, luteolin, is also considered a phytoprogestin, 61 and although it has been reported to antagonize progesterone receptor, 62 recent studies show that apigenin has mixed agonist activity on progesterone receptor. 58 Apigenin and luteolin can modulate the signaling of sex hormones.
Progesterone inhibits myometrial contractility in the pregnant uterus and promotes uterine quiescence during gestation until term. 63 Inhibition of contractility by progesterone can also occur in the nonpregnant uterus, as has been shown that dihydrogesterone, a progesterone derivative, is useful in alleviating dysmenorrhea. 64 The activities of apigenin and luteolin components of TL may also contribute to the effect of TL on the uterus; however, their contribution may be minimal due to the mixed biologic activities.
Jasmonic acid, an α-linolenic acid derivative in plants that also functions as a plant defense metabolite, 65 was also detected in TL. Jasmonic acid interacts with 5HT1B 66 and inhibits nitric oxide production and proinflammatory mediators, 67 while it also selectively activates sarcoplasmic reticulum (SR) Ca2+-adenosine triphosphatase (Ca2+-ATPase). 68 Although not clear how jasmonic acid interacts with 5HT1B, it is known that 5HT1B activation results in increased Ca2+ release and suppression of cAMP, 69 which will lead to smooth muscle contraction. Therefore, if 5HT1B was blocked by jasmonic acid in the uterus, this will cause inhibition of smooth muscle contractility. Inhibition of nitric oxide production 70 as well as inhibition of proinflammatory mediators can also contribute to uterine smooth muscle inhibition.
The SR Ca2+-ATPase is a primary high-affinity Ca2+ transporter in the myometrium, which, when activated, keeps the levels of Ca2+ low. 71 The biological activities of jasmonic acid suggest that jasmonic acid may contribute to the inhibitory action of TL in this study.
Thymonin (5, 6, 4′-trihydroxy-7, 8, 3′-trimethoxyflavone) is a flavonoid found in T. vulgaris. 72 Thymonin, along with luteolin and apigenin, was reported to inhibit smooth muscle contraction in the rat ileum and vas deferens through inhibition of extracellular calcium, 73 just as was observed in this study.
Ajureptoside belonging to the iridoid glycoside phytochemical class 74 was also identified in TL. The biological activity of ajureptoside, as relates to uterine contraction or similar physiological system, is currently unknown. Rhamnorobin (kaempferol 7-α-L-rhamnofuranoside) 75 and quercetin (2-(3,4-dihydroxy phenyl)-3,5,7-trihydroxy-4H-chromen-4-one)) were also identified in TL. Kaempferol is a flavonol reported to have uterotrophic and estrogenic effects 76 and inhibits cAMP-phosphodiesterase, 77 causing inhibition of uterine contraction. Kaempferol has also been shown to inhibit KCl-induced uterine contractions, 78 supporting the activity of TL in this study. Quercetin also inhibited uterine contractions in rats, which was described to have occurred possibly through VGCCs. 79
TL possesses secondary metabolites that contribute to its activity in relaxation of uterine contraction with the flavonoids showing a strong connection to blockade of extracellular Ca2+ release. Further confirmatory studies are, however, required. A schematic diagram showing the action of TL on the myometrium is shown in Figure 6. A limitation to this study was that TL was not investigated on calcium channels and the use of some receptor blockers to further clarify the mechanism of action. This is due to extract exhaustion during the period of study. In addition, studies quantifying the amount of thyme leaves that can be consumed to produce the desired effect were not performed. These are, however, being proposed for future investigations.
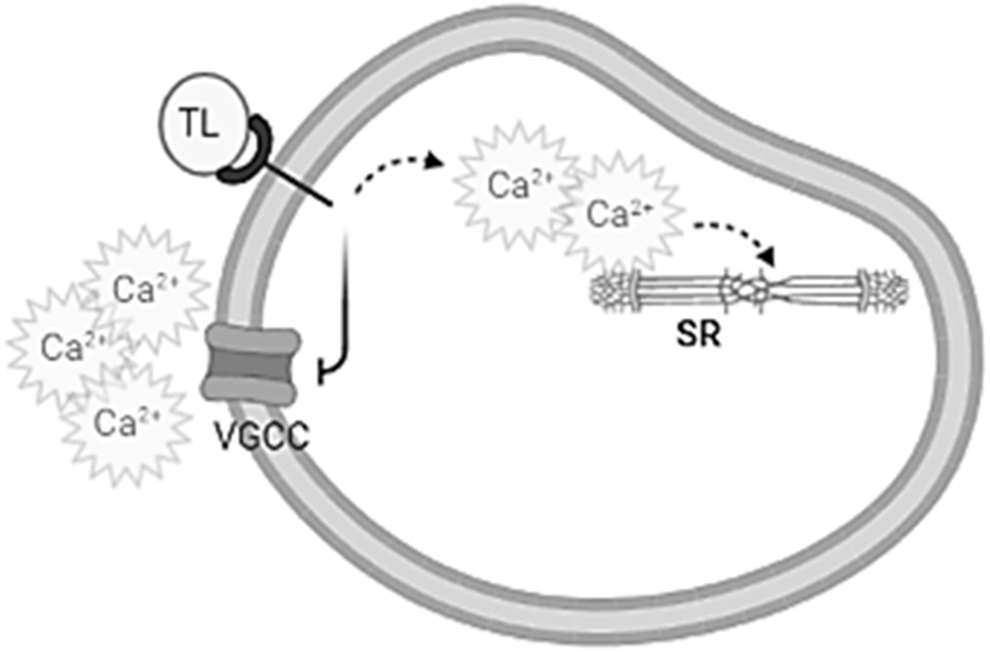
Schematic diagram showing deduced physiological pathways involved in activity of TL on uterine contractility. SR, sarcoplasmic reticulum; VGCC, voltage-gated calcium channel.
This study has shown that aqueous T. vulgaris extract relaxes uterine contractions in the nonpregnant uterus, possibly through blockade of extracellular Ca2+ release. This activity, therefore, makes T. vulgaris useful in counteracting dysmenorrhea.
Footnotes
AUTHORS' CONTRIBUTIONS
E.E.B. performed concept design, literature search, experimental studies, data analysis, article preparation, review, and editing; C.H.K. performed plant material collection, literature search, data acquisition, and article preparation; O.O. performed data acquisition and article review; U.B.E.-O. performed data acquisition and article preparation; and R.E.-E. performed data analysis, article preparation, review, and editing.
ACKNOWLEDGMENT
The authors wish to acknowledge Ms. Glory Uyi Omogiade for her technical assistance during this research.
AUTHOR DISCLOSURE STATEMENT
No competing financial interests exist.
FUNDING INFORMATION
No funding was received for this research.