
Abstract
Hibiscus sabdariffa L. (Hs) calyxes, rich in organic acids, are included in diets in different countries. In recent years, some phytochemicals have been shown to reduce bacterial virulence at sublethal concentrations by interfering with quorum sensing (QS) systems. Therefore, in this study the antivirulence properties of Hs calyxes and two γ-lactones (hibiscus acid [HA] and its methyl ester) in Pseudomonas aeruginosa were analyzed. Acetone and methanol extracts of Hs showed anti-QS activity by inhibiting violacein production (60% to 80% with 250 μg/mL). In molecular docking analysis, the γ-lactones registered a good binding score, which suggests strong interaction with the active site of LasR protein. To verify their effect in vitro, they were isolated from Hs and evaluated in six QS-regulated phenotypes, as well as in ExoU toxin that is released by the type III secretion system (T3SS). At 500 μg/mL they reduced alkaline protease (29–52%) and elastase (15–37%) activity, biofilm formation (∼75%), and swarming (50%), but there was no effect on pyocyanin production, hemolytic activity, or type III secretion. In a mouse abscess/necrosis model, HA at sublethal concentrations (15 and 31.2 μg/mL) affected infection establishment and prevented damage and systemic spread. In conclusion, HA is the first molecule identified with antivirulence properties in Hs with the potential to prevent infections caused by P. aeruginosa.
Introduction
H
Antivirulence is a broad concept that consists in interrupting the production of factors responsible for damage and establishment of pathogenic bacteria, without affecting bacterial viability. 6 Different targets have been proposed, such as biofilms, swarming, two-component systems, fimbriae, pili, secretion systems, pigments, and proteases, among others. 7 However, one of the most promising targets is the inhibition of quorum sensing (QS) systems, as it has a central role in the regulation of several virulence factors. 8 QS is the mechanism that allows bacteria to communicate and exhibit multicellular behaviors. 9 Bacterial communication is necessary for virulence factors to be expressed in a collective and coordinated manner in the population, maximizing success of infectious processes. 8 Pseudomonas aeruginosa is one of the main bacterial models where QS and its role in virulence have been studied. 10 Four QS systems have been identified in this bacterium that activate the expression of virulence genes through an intricate regulatory network between them. 11 Two systems use acyl-homoserine-lactone type autoinducers (LasI-LasR and RhlI-RhlR), whereas the rest use 2-heptyl-3-hydroxy-4-quinolone (PQS) and cis-2-decenoic acid (CDA). 12 These QS systems regulate some virulence mechanisms involved in motility (swarming), iron clearance (proteases, pyoverdine), immune evasion (elastase, alkaline protease), cytotoxicity (pyocyanin), and biofilm formation. However, there are other essential virulence factors, such as the type III secretion system (T3SS), for which regulation by QS is not yet completely clear. 13
Furthermore, it is known that diet plays an important role in health. 14,15 In this regard, it has been suggested that antivirulence molecules (especially QS inhibitors) contained in edible plant species could prevent the establishment and development of chronic infections. 16
Fresh or dried Hs calyxes are used in the preparation of hot and cold beverages and is a common ingredient in several foods. 1 Although their bactericidal properties have been reported, 2,17,18 it is only recently that its antivirulence potential has been investigated. 19,20
Organic extracts of the calyx are characterized by containing a high percentage of organic acids, among which hibiscus acid (HA) (γ-lactone) is the most abundant. 15 It should be noted that, despite the high abundance of acids in the calyx of the plant, not much attention has been paid to them, apart from the fact that HA is not commercially available. 21 Therefore, the objective of this research was to analyze the antivirulence properties of Hs calyxes, as well as HA and its dimethyl ester against P. aeruginosa.
Materials and Methods
Plant material
Two varieties of Hs (“criolla de Guerrero” and “alma blanca”) were analyzed in 2016. Both varieties are cultivated in the state of Guerrero, Mexico, at 18° 20′ 36.6″ N; 99° 30′ 12.6″ W. The calyxes were dried at 50°C (Lab-Line, Ambi Hi-Low Chamber, USA). One hundred grams of powder material was extracted with 900 mL acetone or methanol (JT Baker, USA) for 24 h at room temperature. The process was repeated two more times, supernatant was collected by filtration and the solvent was evaporated.
Isolation and characterization of γ-lactones
The methanol (27.4 g) and acetone (2.58 g) extracts of the variety “criolla de Guerrero” were subjected to a separation process by liquid column chromatography (silica gel, 60 Å; Sigma-Aldrich), using hexane, ethyl acetate, and methanol with increasing polarity (JT Baker) as the mobile phase (Supplementary Table S1).
Infrared spectroscopy
Infrared (IR) spectra were obtained in a PerkinElmer Spectrum 200 Fourier transform spectrophotometer (FT-IR) with universal attenuated total reflectance sampling accessory, at a temperature of 25°C ± 2°C. 1 H and 13 C nuclear magnetic resonance (NMR) spectra were recorded using deuterated chloroform (CDCl3, to dissolve HA) or dimethyl sulfoxide (DMSO-d6, to dissolve hibiscus dimethyl ester [HDM]) at room temperature using a Varian NMR, 400 MHz spectrometer. Tetramethylsilane was used as the internal standard.
Molecular docking
The crystal structure of LasR from P. aeruginosa bound to 3-oxo-C12-HSL was obtained from the protein data bank (accession no. 31X3). HA and HDM three-dimensional models were obtained from the PubChem database (National Center for Biotechnology Information) (
Anti-QS activity of organic extracts
Anti-QS activity was determined by the production of the violacein pigment in Chromobacterium violaceum by a protocol previously reported (Supplementary Data). 25
Virulence factor determination in P. aeruginosa
Pyocyanin inhibition, 26 swarming, 27 biofilm formation, 28 as well as elastase, alkaline protease, 26 and hemolytic activities 29 were evaluated as previously reported (Supplementary Data).
Type III secretion assay
The type III protein secretion assay was performed as previously reported. 30 Proteins secreted to the supernatant were precipitated, concentrated, loaded onto a 15% sodium dodecyl sulfate polyacrylamide gel, transferred to a nitrocellulose membrane, and tested against anti-ExoU polyclonal antibodies. The proteins were detected by immunoblot analysis using the Immobilon Western Chemiluminescent HRP Substrate Kit (Millipore) on X-ray films.
Animals and experimental design
Female CD1 mice (4–8 weeks old) were obtained from the Facultad de Estudios Superiores-Cuautitlán, UNAM. Animals were kept in standard conditions (23°C ± 2°C) with a 12-h light–12-h dark cycle and allowed free access to food and water. The indications of regulation for use and care of animals destined for research at the Hospital Infantil de México-Federico Gómez were always followed (HIM2018-002).
Ethics statements
Studies involving animal subjects Generated Statement: The animal study was reviewed and approved by Hospital Infantil de México-Federico Gómez (HIM2018-002).
Bacterial inoculum and preincubation with γ-lactones
Five milliliters of P. aeruginosa overnight culture were adjusted to an O.D600nm of 0.06 and incubated at 6000 g at 37°C until reaching an O.D600nm of 1.0–1.5. Subsequently, 1 mm of the culture was adjusted to an O.D600nm of 0.08 (≈107 colony-forming unit [CFU]/60 μL), centrifuged at 13,000 g for 2 min, and washed twice with phosphate-buffered saline (PBS). Finally, the bacterial pellet was homogenized in 1 mL of injectable solution (Pisa®), the γ-lactones were added (final concentrations of 7.5, 15, 31.25, 125, and 500 μg/mL), and incubated for 25 min for its subsequent inoculation into animals.
In vivo assay
Based on a previous report. 31 Mice were shaved with an electric shaver and excess hair was removed with a topical hair remover (Nair®). Mice were anesthetized with an intraperitoneal injection of sodium pentobarbital (Pisa) 50 mg/kg, then 60 μL of the inoculum (≈107 CFU) incubated with the different treatments was injected into the subcutaneous space on their right flank. Abscess and necrotic areas were measured every 24 h for a period of 96 h. Four days after inoculation, the livers and the soft tissues containing the necrotic area of the mice were excised and homogenized with 3 mL of PBS. Serial dilutions were done in Luria-Bertani plates to count colony-forming units.
Results
Anti-QS activity of Hs calyxes
None of the extracts affected bacterial growth at the concentrations evaluated, but they did reduce violacein production. The acetone extract of the variety “alma blanca” (AB-A) showed the best effect by inhibiting 86% pigment production at 250 μg/mL, similar to that of the positive control treated with the anacardic acid mixture (AA) (95% with 250 μg/mL) (Fig. 1).

Quorum sensing inhibitory activity of organic extracts from Hs calyxes. The black bars indicate violacein production and white bars indicate growth calculated by the viable cell counting method. Control, dimethyl sulfoxide, (vehicle). 25 Bars represent mean ± standard error. Significant statistical difference between mean values was determined by two-tailed Student's t-test, *P < .05. AA, anacardic acid mixture; AB-A, “alma blanca”-acetone extract; AB-M, “alma blanca”-methanol extract; CG-A, “criolla de Guerrero”-acetone extract; CG-M, “criolla de Guerrero”-methanol extract; Hs, Hibiscus sabdariffa L.
Docking analysis of LasR protein with γ-lactones
Molecular docking analysis suggested that γ-lactones can efficiently interact with the LasR protein-binding site (Fig. 2). To determine the theoretical binding affinity and stability of the complexes coupled between lactones and the LasR protein, energy minimization was performed revealing that the LasR protein had the lowest binding energy when coupled to HA (B.E. −7.39 kCal/mol, Ki 1.66 μM), even lower than that of the natural autoinducer (3-oxo-C12-HSL). While HA deprotonation further facilitates coupling (B.E. −7.26 to −9.06 kCal/mol, Ki 0.228 to 4.51 μM) (Fig. 2 and Table 1). These results indicate that HA can interact with the LasR protein efficiently and suggests a possible mechanism of action related to competition for the LasR active site. Interestingly, the analysis indicated that HA methylation reduces the binding efficiency (B.E. −6.63 kCal/mol, Ki 13.79 μM), however, its binding energy value remains in the same order of magnitude than that recorded for 3-oxo-C12-HSL.

Slice of the binding site of the LasR receptor protein showing the interactions with the autoinducer 3-oxo-C12 homoserine lactone
Molecular Docking Parameters of Hibiscus Acid and Hibiscus Dimethyl Ester Isolated from Hibiscus sabdariffa L. Against LasR Receptor Protein of Pseudomonas aeruginosa
B.E., binding energy (more negative scores indicate higher binding affinities); HA, hibiscus acid; HDM, hibiscus dimethyl ester.
Isolation and characterization of γ-lactones
Since HA and HDM are not commercially available, they were isolated from Hs calyxes. Identification was made by comparative analysis of the NMR and IR values previously reported. 17,32
HA, (2S,3R)-tetrahydro-3-hydroxy-5-oxo-2,3-furandicarboxylic acid: white solid; m.p. 188.74–193.03°C. IR νmax cm−1: 3410, 1797, 1742. 1 HNMR (acetone-d6, 400 MHz) δH: 5.34 (1H, s, CH-COOH), 3.25 (1H, d, J = 17.2 Hz, CHa Hb COHCOOH), 2.80 (1H, d, J = 17.2 Hz, CHa Hb COHCOOH). 13 C-NMR (acetone-d6, 400 MHz) δC: 173.2, 172.3, 167.1, 82.9, 78.4, 42.2.
HDM, dimethyl (2S,3R)-tetrahydro-3-hydroxy-5-oxo-2,3 furandicarboxylate: white solid: m.p. 128–129.54°C. IR νmax cm−1: 3527, 1798, 1741. 1 HNMR (CDCl3, 400 MHz) δH: 5.14 (1H, s, CH-COOMe), 3.95 (3H, s, CH-COOMe), 3.84 (3H, s, COH-COOMe), 3.09 (1H,d, J = 17.5 Hz, H aHbC-C = O), 2.88 (1H,d, J = 17.5 Hz, Ha Hb C-C = O). 13 C-NMR (DMSO-d6): 172.7, 170.8, 166.1, 81.94, 77.93, 53.11, 52.39, 41.55.
Effect of γ-lactones on production of virulence factors
γ-lactones did not exhibit bactericidal activity in any of the evaluated doses (Fig. 3A), but they did affect production of some virulence factors. HA inhibited protease activity by 29.1% at 500 μg/mL (P < .05), but contrary to what was expected, HDM methylations favored a better activity, inhibiting it by 36.2% and 52.4% at 250 and 500 μg/mL, respectively (P < .05) (Fig. 3B). Only HA reduced elastase activity by 37% at 500 μg/mL (P < .05), whereas with HDM the effect was halved at the same concentration and did not show a dose–response effect (Fig. 3C). In contrast, there was no inhibitory effect of γ-lactones on hemolytic activity, pyocyanin production, and the secretion of the effector protein ExoU (Fig. 4).

Concentration-dependent effects of γ-lactones on inhibiting Pseudomonas aeruginosa growth and virulence factors.
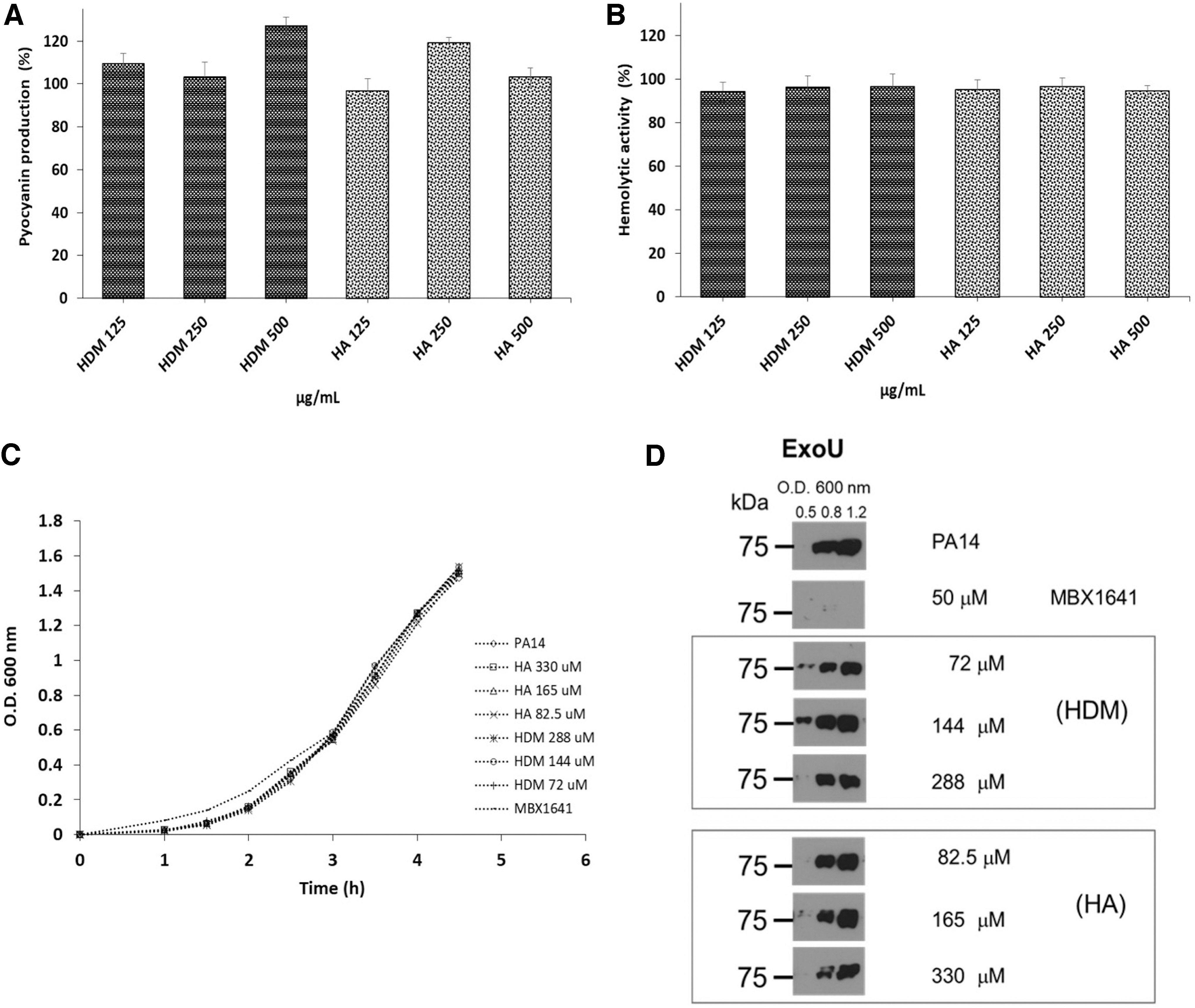
Effect of γ-lactones on the production of P. aeruginosa virulence factors.
The best antivirulence activities of the entire study were observed on swarming motility and biofilm formation. γ-lactones significantly reduced biofilm formation by about 75% at 500 μg/mL (P < .05) (Fig. 3D), whereas for swarming, HA showed better activity with respect to its methylated form, reducing it by 50% at 250 μg/mL and 24.9% at 125 μg/mL (P < .05) (Fig. 3E, F). Interestingly, HDM at concentrations of 125 and 250 μg/mL stimulated swarming motility by about 20% (Fig. 3E).
HA displays an antibacterial effect and reduces damage and dispersal of P. aeruginosa
Subcutaneous inoculation of ∼107 CFU induced the formation of 163.4 mm2 abscesses and 22.9 mm2 necrotic area, on average (Fig. 5A, B), as well as 7% mortality (data not shown). Incubation of the bacterial inoculum with 15 μg/mL HA for 25 min before inoculation, reduced the area of the abscesses to an average of 44.1 mm2 (P < .05), whereas with 31.2 μg/mL, it prevented their formation (P < .05) (Fig. 5A). Similarly, with these concentrations of HA, necrotic tissue did not develop (P < .05) (Figs. 5B and 6), and preincubation of the bacteria with HA did not affect inoculum viability (Fig. 5D).

Effect of HA on P. aeruginosa infection in an animal model. CD1 mice were infected in the subcutaneous space with ∼107 CFU of PA14, previously incubated for 25 min with HA in injectable solution.

Images of animals from a representative experiment subcutaneously infected with P. aeruginosa treated with HA. Maximum abscess formation occurred 24 h postinoculation, whereas necrosis formation occurred at 48 h. In the control group (PA14-V), the bacteria were inoculated through an injectable solution, whereas in the experimental groups (PA14-HA) the inoculum was incubated for 25 min with the different concentrations of HA.
The bacteria established in the inoculation area at an average of log 8.7 CFU/g of tissue 4 days postinoculation, whereas with previous incubation of HA at 7.5 μg/mL it was reduced to log 8 CFU/g and with 15 μg/mL to log 4.33 CFU/g of tissue (P < .05). Interestingly, at a concentration of 31.2 μg/mL, P. aeruginosa establishment completely halted (Fig. 5C).
HA prevented systemic dispersion in all the concentrations evaluated since bacteria were not found in the liver of the animals; in contrast, the control group had an average of log 3.8 CFU/g of tissue (Fig. 5C). Likewise, 100% survival of the inoculated animals previously treated with HA was recorded (Fig. 6). Finally, it should be noted that HA at concentrations of 125 and 500 μg/mL also counteracted the damage caused by the bacteria (Fig. 7A, B), as well as their establishment and dispersion (Fig. 7C), while HDM, at the same concentrations, was not significantly different from the control group without treatment (Fig. 7A–C). However, the effect of HA at these concentrations was attributed to a bactericidal effect caused by a decrease in pH (2.7) in the bacteria preincubation process (Fig. 7D and Supplementary Table S2).

Effect of γ-lactones on P. aeruginosa infection in an animal model at concentrations of 125 and 500 μg/mL. The same experimental conditions mentioned in Figure 5 were used.
Discussion
Many anti-QS molecules have been reported to act by interfering with acyl-homoserine lactones bonding to homologous LuxR receptors. 5,33 In this study, our docking analysis suggested a strong interaction of HA with the active site of the LasR protein. This prediction was partially corroborated in vitro with elastase activity, biofilm formation, and swarming motility. However, for alkaline protease activity, this trend was not observed and there was no effect on hemolytic activity and pyocyanin production. In this regard, although the antivirulence properties of γ-lactones have been poorly explored, an HDM-like molecule called pantolactone ((3R)-3-hydroxy-4,4-dimethyloxolan-2-one) was reported to inhibit the production of pyocyanin. 34 Therefore, this indicates that small structural changes (the position of the methyl groups) favor reduction of some virulence factors in vitro. However, in swarming, acid groups in γ-lactone need to exert a better effect.
Swarming is a phenomenon of collective behavior controlled by QS, which involves the movement of bacteria by flagella on semisolid surfaces using biosurfactants. 35 In P. aeruginosa virulence is higher under swarming conditions, where production of QS-regulated phenotypes (proteases, siderophores, and phenazines) increases, and positive regulation of T3SS gene expression and its effectors is induced, 36 although this regulation seems to be independent of QS systems. 30 Our results suggest that HA acts mainly on some elements of the swarming regulation cascade and has no effect on T3SS activity. In this sense, although it is feasible for HA to inhibit swarming by interference with the LasR active site, it is possible for it to act at other levels of regulation beyond QS, 37 because it is known that other virulence regulators are involved in the expression of swarming, such as two-component systems 38 and temperature sensor systems, 39 among others. 40,41
The difference in activity between HA and its methylated form was also recorded in in vivo assays, where only HA showed antibacterial activity. At the highest evaluated doses, it has a bactericidal effect, which is attributed to a strong decrease in pH. However, at concentrations below 31.25 μg/mL, there is no effect of acidity on the viability of the bacterial inoculum (Supplementary Table S2). Clearly, at these sublethal concentrations, pretreatment of the inoculum with HA blocks the virulence of P. aeruginosa, as it reduces damage and systemic spread in animals. We must mention that, to achieve this effect, HA needs to dissociate partially in the solution, since if the pretreatment is carried out in a buffered solution, the antivirulence activity does not appear (PBS) (Supplementary Table S2 and Supplementary Fig. S1). Docking evaluations in the LasR-binding site with the protonated structure of HA (mainly in position β) support these results (Fig. 2 and Table 1). When the formal charge was assigned to both carboxylates of HA, the form that would be expected at physiological pH, the theoretical Ki diminished seven-fold with respect to the protonated form. Thus, these results suggest that the charge may play an important role in the binding of HA to LasR receptor. In this regard, it has been reported that a slightly acidic environment can enhance the bactericidal effect of some essential oils 42 and organic acids, 43 but in the case of the activity of an antivirulence molecule, as far as we know, this is the first report.
The bactericidal mechanism of some weak organic acids is related to its high lipid permeability in its uncharged state, 44 which favors its entry into the bacteria where it dissociates and causes a decrease in the pH of the cytoplasm. 45 The accumulation of anions inside the bacteria generates osmotic stress and the inhibition of some metabolic processes. 46
In the case of antivirulence activity, it is also necessary for HA to diffuse into the cytoplasm so it can interact with the LasR receptor protein or other possible virulence regulatory elements not analyzed in this investigation. Therefore, other possible antivirulence mechanisms involved in the HA effect at sublethal concentrations in vivo remain to be explored. In this context, some organic acids, such as lactic, acetic, and propionic acids, have recently been reported to inhibit various QS-regulated virulence factors, mainly in Salmonella and Escherichia coli. 47,48
Furthermore, it has been suggested that any natural food source is unlikely to have biologically relevant amounts of quorum quenching (or antivirulence) molecules to eliminate established infections. 16 However, if consumed constantly, the natural food source may help prevent development of chronic infections. 14,49 Hence, we are in the process of understanding the antimicrobial properties of Hs at sublethal concentrations. For example, the antivirulence activity of the aqueous extract of Hs calyxes has recently been reported on Staphylococcus aureus 19 and enteropathogenic E. coli. 20 Finally, unlike other reported antivirulence phytochemicals, it is likely that HA can exert this property when ingested in the diet, since it is soluble in water, it is in high concentrations 15 and its intake is considered safe. 50
In conclusion, HA is the first antivirulence molecule identified in Hs calyxes, whose biofilm and swarming inhibitory effects, as well as antibacterial properties observed in vivo, indicate that it has potential for preventing bacterial infectious diseases.
Footnotes
Acknowledgments
I.C.-J. was supported by Cátedras-CONACyT program. The authors acknowledge Héctor Quezada-Pablo, PhD and facilities granted by the Hospital Infantil de México-Federico Gómez (HIM2018-002). They also acknowledge Norma Espinosa Sánchez, PhD for technical assistance.
Author Disclosure Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationship that could be construed as a potential conflict of interest.
Funding Information
This work was supported by grants from Scientific Development Projects for Solving National Problems/CONACyT Mexico no. 2015-01-402. H.C.-L. thanks CONACYT for her doctoral scholarship (449277). R.G.-C. is funded by CONACYT grant CB 2017–2018 number A1-S-8530.
Supplementary Material
Supplementary Data
Supplementary Figure S1
Supplementary Table S1
Supplementary Table S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.