
Abstract
Jaboticaba (Myrciaria cauliflora), a Brazilian fruit, is a good source of dietary fiber and phenolic compounds, which are concentrated mainly in the peel. These compounds have been considered promising in prevention and treatment of hypercholesterolemia and hepatic steatosis. In this study, we investigated the effects of 4% jaboticaba peel powder (JPP) supplementation on cholesterol metabolism and hepatic steatosis in livers of rats fed a high-fat (HF) diet. The rats were fed a standard AIN-93M (control) diet or an HF diet containing 32% lard and 1% cholesterol, both with and without 4% JPP. The M. cauliflora peel composition revealed a low-lipid high-fiber content and phenolic compounds. The phenolic compounds in JPP, tentatively identified by high-performance liquid chromatography and mass spectrometry (HPLC-MS/MS) analysis, were confirmed to contain phenolic acids, flavonoids, and anthocyanins. Moreover, JPP presented significant antioxidant activity in vitro and was not cytotoxic to HepG2 cells, as determined by the lactate dehydrogenase (LDH) assay. After 6 weeks of treatment, our results showed that JPP supplementation increased lipid excretion in feces, reduced serum levels of total cholesterol and nonhigh-density lipoprotein cholesterol, decreased serum aspartate aminotransferase (AST) activity, and attenuated hepatic steatosis severity in rats fed the HF diet. Furthermore, JPP treatment downregulated expression of ACAT-1, LXR-α, CYP7A1, and ABCG5 genes. Therefore, jaboticaba peel may represent a viable dietary strategy to prevent nonalcoholic fatty liver disease as the JPP treatment alleviated hepatic steatosis through improvement of serum lipid profiles and modulation of mRNA expression of genes involved in cholesterol metabolism.
Introduction
Nonalcoholic fatty liver disease (NAFLD), a chronic hepatic disease, has become a major public health challenge worldwide, and epidemiological studies estimate that global prevalence of NAFLD is higher than 25%. 1
NAFLD is characterized by liver triacylglycerol buildup in the absence of chronic alcohol consumption and any other liver disease. It incorporates a broad spectrum of disorders, ranging from steatosis to cirrhosis. The molecular mechanisms that promote hepatic steatosis are still being clarified. Multiple factors are involved in NAFLD development and progression, such as oxidative stress, inflammation, fibrosis, and changes in gut microbiota. However, increased fatty acid influx from adipose tissue and alterations in adipokine secretion patterns associated with insulin resistance are considered to be the main initiating factors. 2,3
Dysregulation of cholesterol metabolism also plays a recognized role in NAFLD. Cholesterol homeostasis is maintained by a complex mechanism involving biosynthesis, uptake, and excretion. The endogenous synthesis of cholesterol is mainly controlled by the transcription factor, sterol regulatory element-binding protein-2 (SREBP-2), which is able to bind to sterol response elements located in the promoter region of genes related to cholesterol synthesis, activating the transcription of certain genes, such as 3-hydroxy-3-methylglutaryl-CoA reductase (HMGCR) and low-density lipoprotein receptor (LDL-R), and promoting increased intracellular cholesterol concentration. 4
Furthermore, bile acid synthesis by the enzyme, cholesterol-7α-hydroxylase (CYP7A1), is a key pathway in controlling cholesterol homeostasis. CYP7A1 hepatic expression is suppressed by increased bile acid levels and this feedback mechanism might be mediated by the liver X receptor (LXR). Likewise, LXR can modulate the expression of the ATP-binding cassette subfamily G transporters, ABCG5 and ABCG8, which are glycoproteins synthesized in the endoplasmic reticulum and involved in reverse cholesterol transport. 5 –7
Considering that pharmacological options for NAFLD treatment are limited, foods containing bioactive compounds capable of preventing or reducing this disease have been described. 8 In this context, jaboticaba (Myrciaria cauliflora) is a fruit of the Brazilian flora and it contains high levels of dietary fiber and phenolic compounds concentrated mainly in its peel, which are able to promote health benefits. 9,10 Thus, the jaboticaba peel has attracted much attention of researchers because its consumption has been associated with antioxidant, anti-inflammatory, antimicrobial, hypolipidemic, and hepatoprotective effects. Moreover, a beneficial effect on insulin signaling was observed in in vitro, animal, and clinical studies, 11 –17 making it a promising candidate for use in dietary therapies for NAFLD.
However, a clarification of how the jaboticaba peel exerts its hypocholesterolemic and hepatoprotective effects has not been presented so far. In the present study, we proposed that M. cauliflora peel supplementation might prevent NAFLD in rats fed a high-fat (HF) diet. Moreover, we investigated whether bioactive compounds from the jaboticaba peel, such as dietary fiber and phenolic compounds, could exert their biological effects on cholesterol metabolism pathways.
Materials and Methods
Jaboticaba peel powder
Jaboticaba fruits (M. cauliflora), Paulista genotype, were harvested in October 2014 in Cachoeira do Campo, Ouro Preto, MG, Brazil (20°23′45.0″S 43°30′34.2″W—SISGEN number: A06098D). They were selected, cleaned, and peeled. Peels were then dried in an air flow oven at 40°C for 24 h. Dry peels were crushed, pulverized, hermetically sealed in plastic bags, wrapped in black packaging, and then stored at −20°C until use.
Jaboticaba peel composition
Proximate composition of nutrients and antioxidant activity in vitro
Proximate composition was determined according to the Association of Official Analytical Chemists. 18 Antioxidant capacity was determined as described by Brand-Williams et al. 19 and it was evaluated by reduction in absorbance of the DPPH (2,2-diphenyl-1-picryl-hydrazyl) radical, with Trolox (6-hydroxy-2,5,7,8-tetramethylchroman-2-carboxylic acid) being used as a standard antioxidant. The results were expressed as Trolox equivalent antioxidant capacity (μM TEAC/g).
Total extractable and nonextractable phenolics in jaboticaba peel
Extractable (E) phenolics were obtained from 0.5 g of jaboticaba peel powder (JPP) by sequential solvent extractions (3 mL of acidic methanol and 3 mL of acetone/water). 20 The remaining insoluble JPP residue was dried (Savant SpeedVac ISS110-115; Thermo Scientific, Pittsburgh, PA) and subjected to alkaline hydrolysis to release nonextractable (NE) phenolics, as previously reported. 21
Total E and NE phenolics were quantified by the Folin–Ciocalteu micromethod 20 against a gallic acid standard curve (0–0.2 mg/mL, expressed as gallic acid equivalent (GAE)/100 g).
Total extractable monomeric anthocyanin (EMA) content was determined by the differential pH method, as previously described by Giusti and Wrolstad, 22 and expressed as mg of cyanidin-3-glucoside equivalents per 100 g of JPP. Nonextractable polymeric proanthocyanidin (NEPA) was quantified using procedures as previously described. 23
High-performance liquid chromatography and mass spectrometry analysis of JPP phenolics
High-performance liquid chromatography and mass spectrometry (HPLC-MS/MS) analysis of the JPP extractable phenolic fraction was performed using a Finnigan Surveyor HPLC coupled to a Surveyor PDA detector and it was analyzed in the negative ionization mode. 24 Gradient separations were performed using a Sunfire (Waters, MO) C18 column (250 × 4.8 mm, 5 μm) at room temperature. Injections were made into the column by use of a 50-μL sample loop. Mobile phase A consisted of 0.1% formic acid in water, and mobile phase B consisted of 0.1% formic acid in methanol run at 0.4 mL/min.
A gradient was run from 0% to 10% phase B in 3 min and changed to 30% phase B in 20 min, from 30% to 50% phase B in 40 min, from 50% to 70% phase B in 55 min, and from 70% to 100% phase B in 65 min before returning to initial conditions in 3 min. Source and capillary temperatures were set at 300°C, source voltage was set at 3.50 kV, capillary voltage was set at −42 V, and collision energy for the MS/MS analysis was set at 35 eV. The instrument was operated with sheath gas and auxiliary gas (N2) flow rates set at 40 and 5 U/min, respectively. Tentative identification of phenolics based on spectrum and mass was performed using full-scan, data-dependent MS2 scanning from m/z 300 to 800. At least three independent phenolic extractions were performed and analyzed independently by HPLC-MS/MS.
Lactate dehydrogenase assay
The cytotoxicity potential of JPP was assessed by lactate dehydrogenase (LDH) leakage into the culture medium of a human hepatoma cell line (HepG2) acquired from the Cell Bank of the Federal University of Rio de Janeiro. HepG2 cells were cultured in DMEM supplemented with 10% fetal bovine serum, 100 U/mL penicillin, and 100 μg/mL streptomycin and maintained in a humidified atmosphere containing 5% CO2 at 37°C. Briefly, 0.5 × 105 cells were seeded in 12-well plates (1 mL of cell solution/well) 24 h before the treatments. Posteriorly, the culture medium was removed and serum-free medium without phenol red was added containing different concentrations of JPP (25–800 μg/mL). After 24 h of treatment, the medium was aspirated and centrifuged at 200 g for 5 min to obtain a cell-free supernatant.
Cells from the positive control group (PC) were treated with 1% Triton 100 × . LDH activity was analyzed using a commercial kit (Labtest Diagnóstica) following the manufacturer's recommendations, and LDH activity was expressed as U/L.
Animals and diets
Thirty-two, 4–5-week-old, female Fischer rats weighing ∼140 g were obtained from the Laboratory of Experimental Nutrition at the School of Nutrition of the Federal University of Ouro Preto (UFOP). The rats were housed individually under a 12-h light–12-h dark cycle under temperature-controlled conditions, with food and water ad libitum. Animal procedures were approved by the Ethics Committee for Animal Research of the University (Protocol N° 2014/53).
Initially, rats were divided into two experimental groups: the control (C) group fed the AIN-93 M standard diet 25 and the HF group that received an HF diet (32% lard and 1% cholesterol). After 2 weeks, the C and HF groups were subdivided into C with jaboticaba supplementation (CJ) and HF with jaboticaba supplementation (HFJ) groups, respectively. The CJ and HFJ groups received 4% JPP in their diet (Table 1). After 6 weeks, the rats were fasted for 12 h, anesthetized with isoflurane, and euthanized by exsanguination. Blood was collected in polypropylene tubes and centrifuged at 3000 g for 15 min. Serum was removed and stored at −80°C. The liver was collected and weighed. The small hepatic lobe was stored in buffered formalin for histopathological analysis, and the rest of the liver was submerged in liquid nitrogen and immediately stored at −80°C.
Composition of the Experimental Diets (g/kg Diet)
Mineral and bvitamin mixtures as recommended by the AIN-93M rodent diet.
Conversion factors: protein, 4 kcal/g; fat, 9 kcal/g; and sugars, 4 kcal/g.
HF, high-fat; JPP, jaboticaba peel powder.
Biochemical analysis of serum biomarkers associated with NAFLD pathogenesis
Triglycerides (TG), total cholesterol (Total-c), high-density lipoprotein cholesterol (HDL-c), and the serum activities of alanine aminotransferase (ALT) and aspartate aminotransferase (AST) were quantified using kits from Labtest (Lagoa Santa, MG, Brazil). Non-HDL cholesterol (non-HDL-c) was calculated as the difference between Total-c and HDL-c.
Fecal analysis
Total lipids were extracted from feces using the chloroform/methanol solution (2:1, vol/vol), as described by Folch et al. 26 Total fecal lipids were obtained by solvent evaporation.
Liver morphology
Liver samples were fixed in 10% buffered formalin and embedded in paraffin. Tissue sections (4 μm) were cut with a microtome, semiautomatic, mounted, and stained with hematoxylin and eosin (H&E) on precleaned and degreased glass slides. Photomicrographs were taken using a Leica DM5000 microscope coupled to a digital camera at 400 × magnification. In the liver, steatosis was measured as described by Liang et al. 27 and inflammation was evaluated by counting the number of inflammatory cells using Leica QWin Plus software.
Real-time quantitative reverse transcription–polymerase chain reaction assay
Total RNA was isolated from the liver using the Total RNA Isolation System (Promega Corporation, Madison, WI) according to the manufacturer's instructions. cDNA was synthesized from 2 μg of total RNA with random primers using the High-Capacity cDNA Reverse Transcription Kit (Applied Biosystems, Foster City, CA). Quantitative real-time polymerase chain reaction was performed using the Power SYBR® Green PCR Master Mix reagent (Applied Biosystems). The polymerase chain reaction conditions were as follows: 50°C for 2 min, 95°C for 10 min, and then 40 cycles at 95°C for 15 s and 60°C for 1 min. The specificity of the products obtained was confirmed by melt analysis of amplified product dissociation curves.
The data obtained were analyzed using the comparative Cq method. Target gene expression was determined relative to the expression of the endogenous 18S ribosomal RNA gene. All analyses were performed in triplicate. Primer sequences used are presented in Supplementary Table S1.
Statistical analysis
Statistical analysis was performed using GraphPad Prism, v.6.00, for Windows (GraphPad Software, San Diego, CA). Normality of data was tested using the Kolmogorov–Smirnov test. Data were analyzed by one-way analysis of variance (ANOVA), followed by Tukey's post hoc test, and all data are expressed as the mean ± standard error of the mean (SEM). Nominal data (macrovesicular and microvesicular steatosis) were evaluated by the chi-square test. Results were considered statistically significant when P < .05.
Results
Analysis of JPP
The proximate composition of JPP revealed high levels of carbohydrates and dietary fiber, mainly in the insoluble fraction (26.58%), and low levels of lipids (<1 g/100 g of JPP). The content of total phenolic compounds was 14.59 g GAE/100 g of JPP, represented by 12.10 g GAE of E and 2.49 g of NE phenolics. In addition, the EMA and NEPA levels in JPP were 0.82 g C3G/100 g and 1.38 g C3G/100 g, respectively (Table 2).
Centesimal Composition, Phenolic Compounds, and Antioxidant Activity of Jaboticaba Peel Powder
Analyses were performed in triplicate and data are presented as mean ± SEM.
Carbohydrates = 100 − (moisture + protein + lipids + total ash + total dietary fiber).
Nitrogen conversion factor in protein: 6.25.
Total monomeric anthocyanin was dosed using the extractable phenolic fraction.
Total polymeric proanthocyanidin was dosed using the nonextractable phenolic fraction.
C3G, cyanidin-3-glucoside; E, extractable phenolic; GAE, gallic acid equivalent; I, insoluble dietary fiber; NE, nonextractable phenolic; PCN, proanthocyanidin; S, soluble dietary fiber; SEM, standard error of the mean; TEAC, Trolox equivalent antioxidant capacity.
Furthermore, the tentative identification of phenolic compounds in the JPP extractable phenolic fraction was performed by identifying the m/z as well as UV-visible spectrum (Supplementary Table S2 and Supplementary Fig. S1). Even though compound identity was not confirmed with pure standards, the HPLC-MS/MS analysis allowed tentative identification of the compounds presented in Supplementary Table S2 based on the UV-visible spectrum and mass fragments. Phenolics in JPP included a mixture of phenolic acids (ellagic acid), anthocyanins (delphinidin-3-O-glycoside and cyanidin-3-O-monoglucoside), and flavonoids (flavonol type, quercetin-3-glucoside, and quercetin-3-rhamnoside).
The antioxidant capacity, as determined spectrophotometrically through the DPPH method, was 797.73 ± 8.02 μM TEAC/g (Table 2).
LDH is an intracellular enzyme and its presence in the extracellular medium indicates cellular damage. In this study, LDH assay performed using HepG2 cells that were incubated for 24 h with 25, 50, 100, 200, 400, 600, and 800 μg/mL JPP revealed that none of the concentrations evaluated showed cytotoxicity since LDH activity (U/L) in JPP concentrations was not statistically different from the control group (C) (Fig. 1).
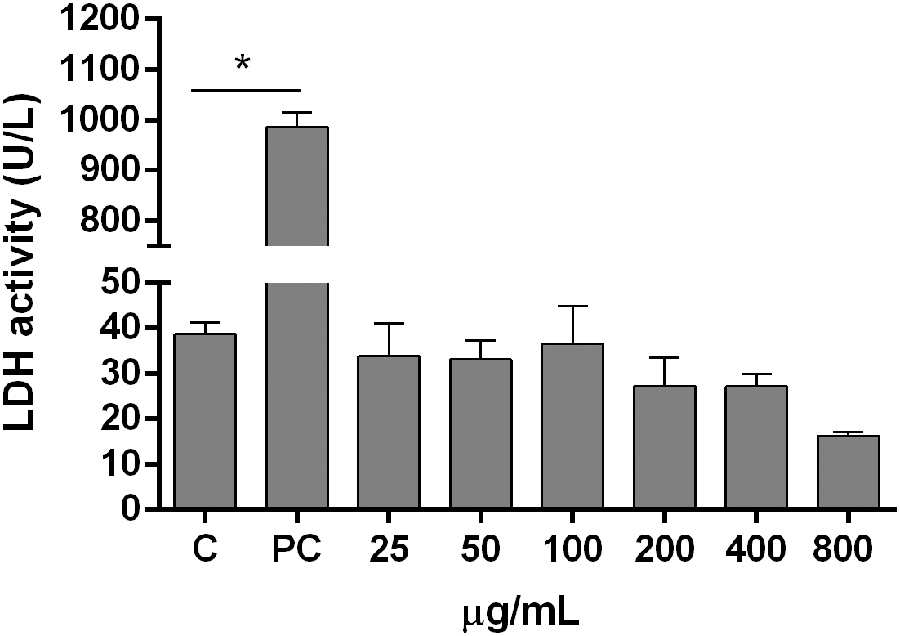
LDH activity in HepG2 cells treated with Myrciaria cauliflora peel (25–800 μg/mL) for 24 h. The bars represent the mean ± SEM (n = 6), data were compared with the C group. Statistical analysis was performed with one-way ANOVA. *Indicates statistical significance (P < .05). In the PC, the cells were lysed in 1% Triton 100 × . ANOVA, analysis of variance; C, control group; LDH, lactate dehydrogenase; PC, positive control group; SEM, standard error of the mean.
Effects of the HF diet and JPP treatment on obesity biomarkers
Final body weight, weight gain, and liver weight were similar among HF and HFJ groups and higher than the control group. JPP intake did not alter these parameters. However, food intake was lower in the HF groups compared with the control groups (Table 3 and Fig. 2).
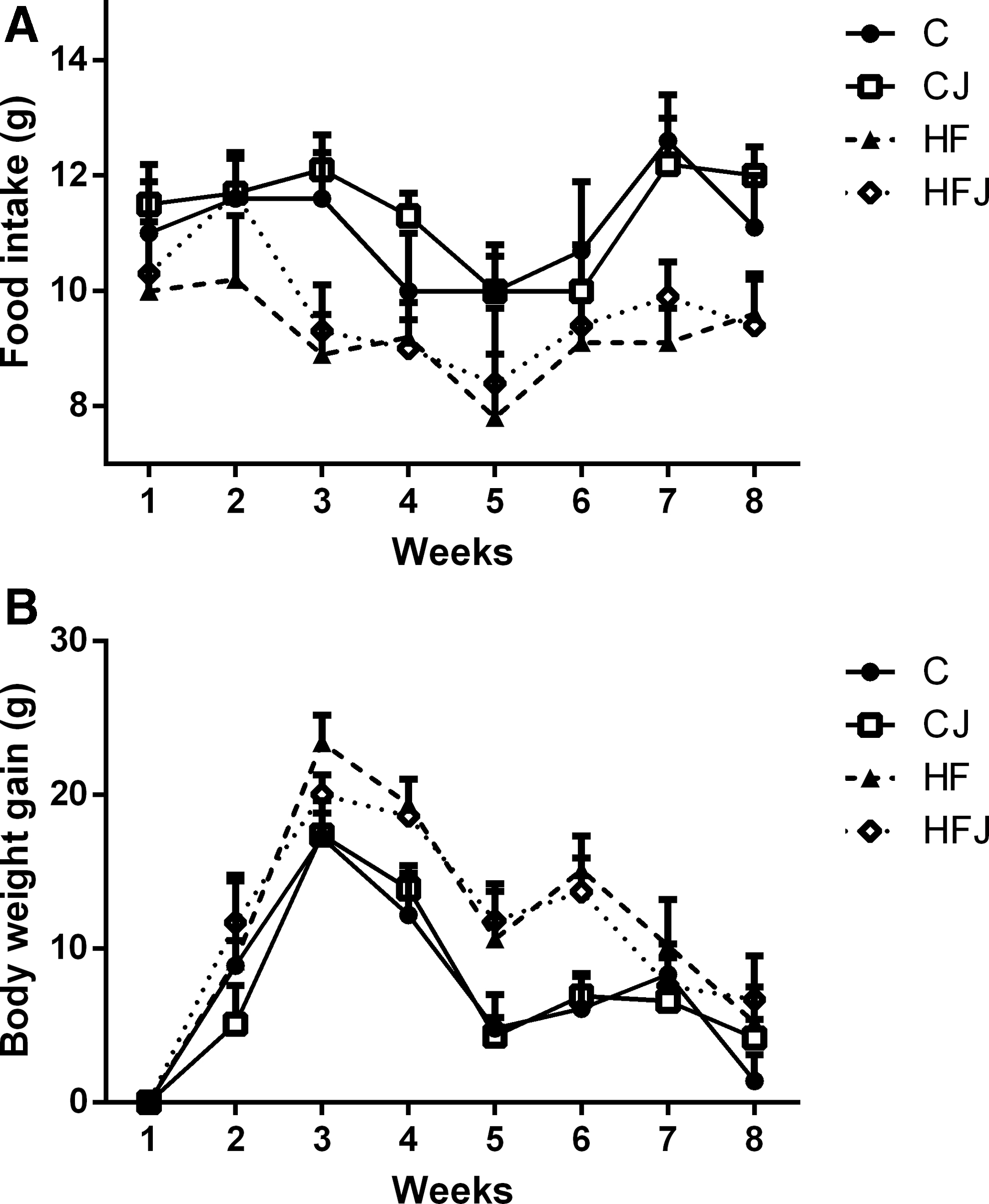
Analysis of food intake
Effect of Jaboticaba Peel Intake on Body and Liver Weights, Food Intake, and Fecal Excretion
Data are presented as mean ± SEM (one-way ANOVA). Different superscript letters within a row indicate statistical significance (P < .05).
Food efficiency = body weight gain/food intake.
Liver relative weight (%) = (liver weight/final body weight) × 100.
ANOVA, analysis of variance; C, control; CJ, control plus 4% JPP; HFJ, high-fat plus 4% JPP.
Fecal excretion was 57.41% and 35.71% higher in the CJ and HFJ groups, respectively, when compared with the C and HF groups. Furthermore, HF animals increased fecal lipid content excretion and the JPP supplementation group significantly intensified it (Table 3).
Effect of JPP treatment on serum lipid profiles
Our results indicate that after 8 weeks, rats fed the HF diet presented alterations in serum lipid profiles since a significant increase in Total-c (3.6-fold) and non-HDL-c (9.07-fold) levels and decrease in HDL-c (1.6-fold) levels were observed compared with rats fed the control diet. JPP intake significantly lowered Total-c and non-HDL-c levels in the HFJ rats (P < .0001) (Fig. 3).

Jaboticaba peel intake ameliorated the serum lipid profile in rats fed the HF diet. Statistical analysis was performed with one-way ANOVA. Different superscript letters indicate statistical significance (P < .05). Values are presented as means ± SEMs (n = 8). C, control; CJ, control plus 4% JPP; HDL-c, high-density lipoprotein cholesterol; non-HDL-c, non-HDL cholesterol; TG, triglycerides; Total-c, total cholesterol.
Effect of JPP treatment on liver biomarkers associated with NAFLD
Rats fed the HF diet exhibited increases in liver fat content and in the activity of serum enzymes, ALT and AST (Fig. 4C, E, and F, respectively), in addition to presenting hepatomegaly (Table 3). Furthermore, histological analysis showed an increase in hepatic lipid deposition in the HF diet rats, characterized by macro- and microvesicular steatosis. Then, the macrovesicular steatosis grade in livers in the HF group was higher compared with the C group (Fig. 4C and E). These results indicated a worsening of liver function in the HF group and demonstrated that rats fed the HF diet developed NAFLD.
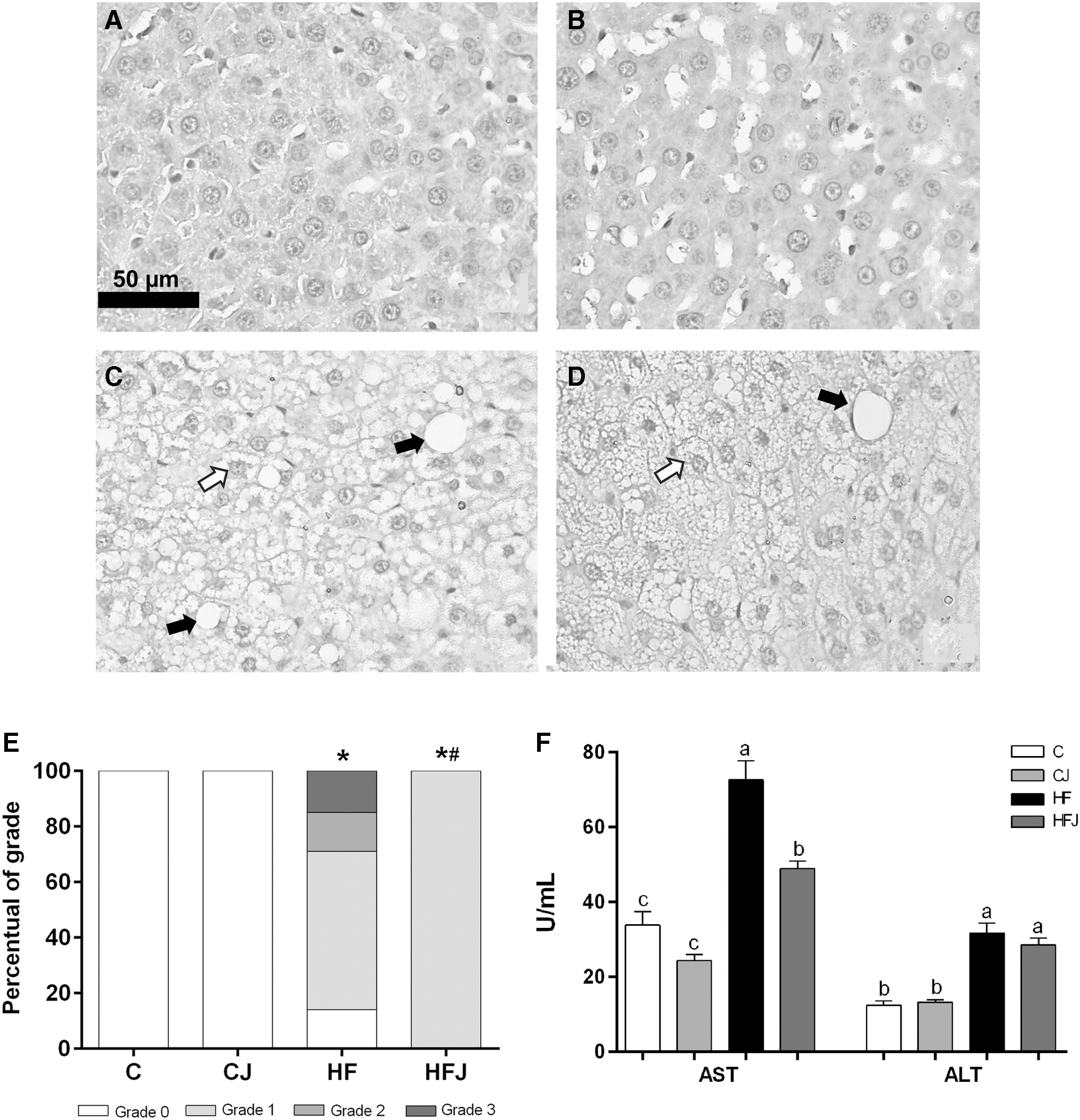
Jaboticaba peel supplementation attenuated hepatic steatosis and improved liver function. Representative hematoxylin and eosin-stained histological sections of livers from rats fed a control or HF diet treated with JPP. Hepatocyte abnormalities were not observed in the C group
JPP treatment reduced 40.93% of serum AST (Fig. 4F) compared with the HF diet group. In addition, JPP attenuated the severity of hepatic steatosis, as evidenced by the presence of only the discrete grade of macrovesicular steatosis in the HFJ group (Fig. 4D, E).
Moreover, the analysis of microvesicular steatosis showed that HF and HFJ groups presented 100% of grade 3, and the analysis of liver inflammation revealed no difference between all experimental groups regarding the number of inflammatory cells (data not shown).
Effect of JPP treatment on the expression of genes related to cholesterol metabolism
The mRNA expression levels of LDL-R and apolipoprotein B100 (ApoB100) were lower in the HF group than in the C group, and JPP intake cotreated with the HF diet reduced ApoB100 expression when compared with the HF group (2.65-fold, P < .0001) (Fig. 5A). As also shown in Figure 5A, expression levels of SREBP-2 and HMGCR exhibited similar profiles. These results showed that both mRNA levels tended to decrease in the HFJ group compared with the HF group (P > .05). However, SREBP-2 and HMGCR mRNA expression levels were significantly lower in the HFJ group compared with the C group.
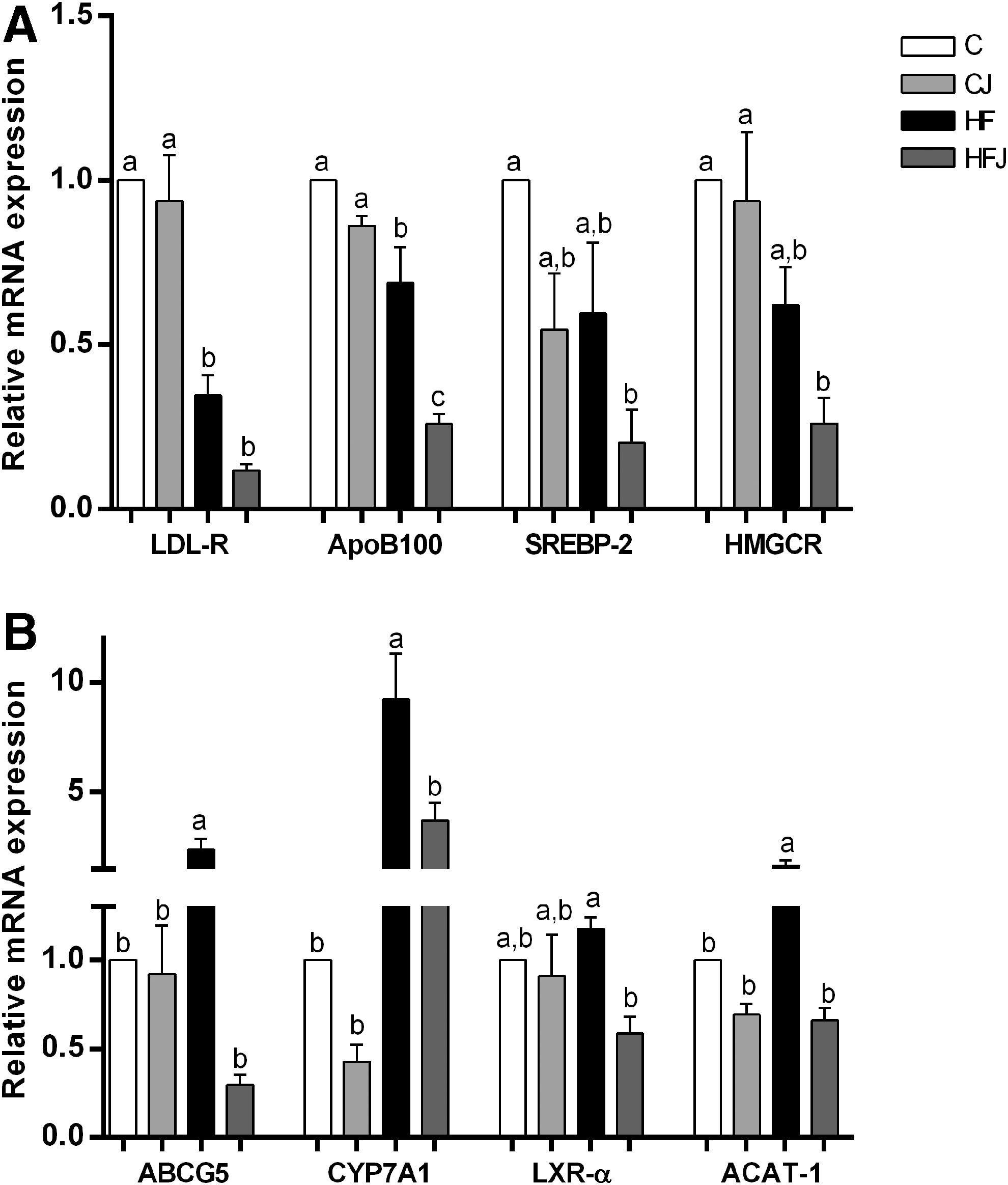
Jaboticaba peel intake modulated the expression of genes involved in lipid metabolism. Relative expression of mRNA of genes involved in cholesterol endocytosis and biosynthesis
We also determined the hepatic expression of ABCG5, CYP7A1, and LXR-α (Fig. 5B). The results showed a significant increase in ABCG5 and CYP7A1 gene expression in the HF group compared with the C group. JPP treatment significantly reduced expression of both mRNAs in the HFJ group to levels similar to controls, including the LXR-α gene.
In addition, mRNA expression of acyl-CoA cholesterol acyltransferase 1 (ACAT-1) was increased in the HF group, and JPP intake reduced ACAT-1 expression when compared with the HF group (2.51-fold, P < .0001) (Fig. 5B).
Discussion
In this study, we investigated effects of the jaboticaba peel, a Brazilian fruit source of polyphenols and dietary fiber, on NAFLD induced by an HF diet in rats. Supplementation of the HF diet with 4% M. cauliflora peel was able to prevent metabolic complications involved in NAFLD since JPP intake was associated with improvements in serum lipid profiles and liver function, reduction of hepatic macrovesicular steatosis severity, increased fecal lipid excretion, and modulated mRNA expression of genes involved in cholesterol metabolism.
The jaboticaba peel dose used in this study was selected considering an average daily intake of ∼20 g by a healthy individual who consumes 2000 kcal/day. This dose has already been shown to be beneficial to health, such as improvement of lipid metabolism in obesity experimental models, 12,13 and it has already demonstrated benefits in humans. 15
Evidence indicates a relationship between diet and chronic diseases, and functional foods have been highlighted as sources of bioactive compounds that may exert health benefits. The jaboticaba peel has polyphenols as the most abundant group, with flavonoids and anthocyanins being predominant, which have been identified as potential antioxidants. 28,29
The tentatively identified phenolic compounds (by HPLC-MS/MS analysis) confirmed that JPP has phenolic acids, flavonoids, and anthocyanins. This result is supported by other studies that focused on the phytochemical characterization of jaboticaba extracts 15,30,31 and provide evidence of their stability during the drying process. Corroborating with our result, a study found that JPP was obtained by convective drying, the same process used in the present study, presenting a high content of anthocyanins and ellagic acid, as well as other bioactive substances. 32 Considering that the jaboticaba fruit is highly perishable and the peel is generally discarded, this result also supports the idea that JPP, obtained through a simple and low-cost technology compared with other widely applied techniques (freeze-drying and spray-drying), can be a good, cheap, and safe strategy for use of the jaboticaba peel.
Likewise, the carotenoid content, also present in the jaboticaba peel, may have contributed to the health benefits observed in rats fed the HF diet. 31 Thus, the phenolic compounds that were identified in JPP probably supported the in vitro antioxidant capacity since they have the ability to stabilize free radicals.
Moreover, in our study, the E and NE phenolic content in jaboticaba peel revealed a higher concentration of those fractions. Most studies on phenolic compounds have focused on the aqueous supernatant (E). Nevertheless, significant quantities of phenolic compounds remain in the solid residue after extraction (NE). 33 The nonextractable bound polyphenolics, mostly proanthocyanidins, phenolic acids, and hydrolyzable tannins, are usually more stable during processing and storage. Therefore, they may escape degradation under the upper gastrointestinal digestion conditions and can be absorbed into blood plasma after being released by intestinal microflora fermentation, thus promoting health benefits. 33
The histological hallmark of NAFLD is steatosis, an accumulation of hepatic TG. Diets rich in saturated fats and cholesterol are usually efficient in increasing liver lipid accumulation once they induce changes in lipogenesis and lipolysis, culminating in hepatic steatosis and inflammation of the tissue. 34,35 Indeed, our HF diet caused liver damage in rats, as indicated by the increase in liver weight and the hepatic enzymes, AST and ALT, as well as by the presence of hepatic steatosis, as evidenced by micro- and macrovesicular fat droplets.
Scientific reports have shown that lipoprotein metabolism is also dysregulated in NAFLD. An abnormal plasma lipoprotein concentration reflects disturbances in the homeostasis of TG and cholesterol, indicating a pattern of atherogenic dyslipidemia. 36 –38 Thus, our data supported the idea that a diet rich in saturated fats and cholesterol promotes development of NAFLD and its metabolic complications.
We demonstrated that jaboticaba peel treatment alleviated liver damage because its addition to the HF diet reduced AST activity and levels of Total-c and non-HDL-c in serum. Moreover, the JPP treatment of 6 weeks attenuated the severity of hepatic macrovesicular steatosis, even though it was not efficient to decrease liver weight and/or weight gain. Probably, the JPP experimental time used in the present study was not enough to promote these effects, contrasting with Batista et al. 14 who observed a reduction in the hepatic steatosis area as well as in liver weight and total weight gain in the HF animals that received 4% Myrciaria jaboticaba berry peel during a 10-week trial.
Scientific reports have observed that HF diet intake increases hepatic fat and cholesterol and reduces total body water in rats; however, these changes in blood biochemistry were not reflected by anthropometric variables. It is possible that the duration of the study intervention was not enough to change body composition, but enough to modify the biochemical profile since it changes more readily, or that a diet with different fat proportions could be necessary to modify body composition. 39 Likewise, it is important to emphasize that an alteration in steatosis severity may already have minimized the risk of liver damage progression. 40
Several studies have demonstrated beneficial effects of the jaboticaba peel on the serum lipid profile and the liver in different experimental models using HF diets. 14,16,41,42 Consistent with our results, Lage et al. 43 showed that rats fed a hypercholesterolemic diet supplemented with 3% jaboticaba skin flour reduced serum AST and ALT activities and also decreased liver macrovesicular steatosis after 28 days of treatment.
Besides that, Batista et al. 13 observed that rats that received the HF diet with 1% and 4% of freeze-dried jaboticaba peel presented a reduction of serum saturated fatty acids, in addition to increasing antioxidant defenses. Indeed, the low levels of circulating saturated lipids as well as cholesterol might act as a protection factor against NAFLD. Additionally, a recent study by Batista et al. 14 also showed that the addition of 4% M. jaboticaba berry peel to the HF (31% lard) diet reduced dyslipidemia and hepatic steatosis in male Swiss mice. However, the molecular mechanisms by which the jaboticaba peel exerts these effects have not been completely clarified.
One possible mechanism that was proposed for reduction in serum cholesterol levels is the effect of viscous soluble fiber such as pectin, a soluble fiber found in jaboticaba peel, on cholesterol metabolism. 14,44 Viscous fibers can reduce transit time through the small intestine, promoting a laxative effect and an increase in fecal weight, which impairs the uptake of dietary cholesterol and prevents the reabsorption of bile acids. Indeed, the addition of 4% JPP increases fecal weight in rats fed both diets (standard and HF), as well as an HF diet supplemented with JPP presented higher fecal lipid excretion, indicating a possible beneficial effect of viscous soluble fibers on intestinal lipid absorption, as found by Batista et al. 14
The alterations in cholesterol homeostasis in the pathogenesis of NAFLD caused by foods containing bioactive compounds and dietary fiber play a key role in prevention of NAFLD progression to NASH (nonalcoholic steatohepatitis).
In the present study, we measured mRNA expression of genes involved in cholesterol synthesis (SREBP-2 and HMGCR), but jaboticaba peel intake did not change gene expression. Interestingly, jaboticaba peel treatment downregulated ApoB100, which encodes an apolipoprotein found in lipoproteins (e.g., VLDL and LDL) that bind the LDL-R, promoting cholesterol uptake. It is well known that uptake of LDL-c through LDL-R leads to inhibition of HMGCR by reducing SREBP-2 activation, 45 thus functional foods are usually LDL-R activators, removing LDL-c from plasma. 46 In our study, a significant reduction in LDL-R gene expression was observed in both HF groups compared with the control group, indicating a possible high concentration of intracellular cholesterol.
Another pathway related to cholesterol homeostasis is the conversion of cholesterol to bile. Cholesterol oxidation by CYP7A1, an enzyme that limits the rate of bile acid synthesis, is the main metabolic pathway by which cholesterol is eliminated from the body. It is regulated primarily by a nuclear receptor (LXRα) that responds to elevated levels of intracellular cholesterol, increasing the transcription of genes that control cholesterol efflux and excretion through bile such as CYP7A1 and the transporters, ABCG5 and ABCG8. 47,48 We found that rats fed the HF diet exhibited higher expression of CYP7A1 and ABCG5, whereas JPP treatment restored their expression as well as LXRα expression to the levels observed in the control group. Studies indicate that flavonoids may regulate lipid synthesis through modulation of LXRα, and its inhibition by polyphenols through AMP-activated protein kinase activation has been shown to be a protective factor against NAFLD. 8,49
Moreover, jaboticaba peel intake negatively modulated the mRNA expression of ACAT to levels that were similar to controls, as identified by Park et al. 50 with red pericarp glutinous rice rich in polyphenols in mice fed a high-cholesterol diet. ACAT is an enzyme that catalyzes the synthesis of cholesteryl ester from free cholesterol and long-chain fatty acyl-CoA and it is considered a promising therapeutic target for reducing atherosclerosis and cardiovascular disease. 51
Overall, these results suggested that JPP supplementation yielded benefits for liver health through modulation of the expression of genes involved in cholesterol metabolism.
Conclusions
Thus, we conclude that JPP obtained through the convective drying process was a source of phenolic compounds and dietary fiber, in addition to not being cytotoxic to HepG2 cells. Furthermore, supplementation of the HF diet with JPP for 6 weeks promoted an increase of fecal lipid excretion, decreased levels of Total-c and non-HDL-c in serum, improved liver function, and attenuated the severity of hepatic steatosis. Such benefits were associated with the JPP-mediated downregulation of genes involved in cholesterol metabolism signaling, including ACAT-1, LXR-α, CYP7A1, and ABCG5. These findings indicate that JPP consumption may represent a viable, safe, and promising dietary approach to attenuate hepatic steatosis related to metabolic disorders such as NAFLD.
Footnotes
Acknowledgments
The authors thank Dr. Steve Talcott for his support in HPLC-MS/MS analysis and Sabrina Peixoto and Andrea Urrutia for their technical assistance. The authors thank the Federal University of Ouro Preto and Texas A&M University. The authors also thank Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES/Brazil) for providing N.N.L. the PDSE scholarship/process number 88881.132176/2016-01.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by Fundação de Amparo à Pesquisa do Estado de Minas Gerais—FAPEMIG (grant no. APQ-03322-15), and it was also financed in part by the CAPES/Brazil — Finance Code 001.
Supplementary Material
Supplementary Figure S1
Supplementary Table S1
Supplementary Table S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.