
Abstract
Consumption of high-fructose diets early in life increases the risk of developing metabolic disorders, including nonalcoholic fatty liver disease (NAFLD). Zingerone, an alkaloid isolated from Zingiber officinale, has been demonstrated to reverse obesity and fatty liver in adult male rats. We investigated the potential preventive effects of neonatally administered zingerone on the development of fructose-induced NAFLD in male and female rats. Four-day-old male (n = 35) and female (n = 44) rat pups were randomized and gavaged with: 10 mL/kg body weight (bwt) of distilled water (C), 10 mL/kg bwt of 20% fructose solution (Fr), 10 mL/kg bwt of 20% fructose solution +40 mg/kg bwt of zingerone (ZFr), and 40 mg/kg bwt of zingerone (Z) daily for 14 days. After weaning, all groups continued on unlimited standard rat feed; however, groups C and Z had plain drinking water, whereas groups Fr and ZFr had unlimited 20% fructose solution to drink for 10 weeks. Rats on the high-fructose diet (Fr) compared with the negative controls (C) had significantly increased hepatic lipid content (in %, males: P = .0002; females: P < .0001, analysis of variance [ANOVA]) and hepatic steatosis score (in %, males: P = .0018; females: P < .0022, Kruskal–Wallis ANOVA). Zingerone prevented (P < .05) the fructose-induced increase in hepatic steatosis in both sexes. The plasma alanine aminotransferase activity, levels of uric acid, TBARS (thiobarbituric acid reactive substances), IL-6 (interleukin-6), and TNF-α (tumor necrosis factor alpha) were not different (P > .05, ANOVA) across the different treatment groups in both sexes. No difference (P > .05, ANOVA) was observed between the two sexes for treatment, sex and interaction effects with regard to hepatic lipid content, and measured blood parameters. The use of zingerone neonatally should be further investigated as a strategic prophylactic intervention for the prevention of long-term high-fructose diet-induced NAFLD.
Introduction
The average global prevalence of nonalcoholic fatty liver disease (NAFLD) is 25%. 1 This prevalence increases with increasing age and male sex. 2 NAFLD is a clinical condition characterized by fatty infiltration of hepatocytes in the absence of significant alcohol consumption. 3 Histologically, NAFLD initially presents as simple steatosis, which in 10–15% of patients, progresses to nonalcoholic steatohepatitis (NASH). 3 NASH is characterized by inflammation and can further progress to fibrosis and eventually cirrhosis of the liver leading to hepatocellular carcinoma. 4 NAFLD is asymptomatic in most patients but associated with obesity and risk factors of metabolic syndrome. 5 It is the most common cause of chronic liver disease globally. 5 Despite limited data on the global economic burden of managing NAFLD, the annual medical cost of managing NAFLD is ∼62 billion dollars in the United States, 6 and the cost of liver transplants is expected to rise with increasing prevalence of the condition. 7
Despite NAFLD being associated with the components of metabolic syndrome (obesity, dyslipidemia, or insulin resistance), 8 recent evidence shows that NAFLD can develop independently in the absence of the aforementioned metabolic disorders. 9 Nutrition, environment, gut microbiota, and genetics/epigenetics are among the factors implicated in the pathogenesis of NAFLD. 4
Environmental and nutritional influences during periods of developmental plasticity, such as the neonatal period, can result in epigenetic changes, which can alter phenotype physiology and susceptibility to the development of metabolic diseases, including NAFLD, later in life. 10 Excessive intake of fructose during the perinatal period may serve to sensitize the liver to subsequent nutritional, metabolic, and environmental events, which trigger the development of NAFLD, hepatomegaly, and increase in general body mass later in life. 4,11,12 Additionally, high dietary fructose consumption is a major cause of NAFLD due to the unregulated nature of fructose metabolism. 13
Prolonged excessive hepatic fructose metabolism causes adenosine triphosphate (ATP) depletion, which stimulates uric acid synthesis leading to mitochondrial dysfunction and oxidative stress. 13 These events stimulate the production of reactive oxygen species that eventually lead to inflammation characterized by elevated proinflammatory cytokines, apoptosis, and fibrosis. 4
Metabolism of fructose also affects the expression of genes and receptors involved in the different steps of hepatic lipid accumulation and hepatic inflammation. 14 These genes and receptors include those associated with carbohydrate response element-binding protein (ChREBP), sterol regulatory element binding protein 1 (SREBP-1), acetyl-CoA carboxylase 1, carnitine palmitoyltransferase 1, tumor necrosis factor alpha (TNF-α), and peroxisome proliferator-activated receptor alpha (PPARα). 14 Dysregulation of these genes and receptors can lead to the development of NAFLD secondary to hepatic lipid accumulation and inflammation. 4
The inflammatory, apoptotic, and lipid peroxidation processes associated with NAFLD can generate TNF-α, interleukin-6 (IL-6), and thiobarbituric acid reactive substances (TBARS), 3 which damage hepatocytes leading to elevation of key liver enzymes in circulation, particularly alanine aminotransferase (ALT), although this elevation does not correlate with the extent of steatosis, inflammation, or fibrosis. 5 NAFLD can be managed effectively in its early stages of pathological development.
Despite no specifically approved pharmacological drug treatment for NAFLD, 1 regular exercise and good dietary habits are the first-line approaches in the early management of NAFLD. 15 The use of synthetic pharmacological agents, such as fenofibrate used in the management of specific risk factors of metabolic syndrome, is limited as these agents are usually expensive, require chronic use, inaccessible in some communities and monotherapeutic. 16,17 There is need for innovative prophylactic interventions against NAFLD using locally available resources.
Globally, communities are increasingly opting for plant-derived medications for their health care needs. 16 The medicinal potential of natural agents for protection against adverse neonatal metabolic programming of NAFLD needs to be explored. Previous studies have used fructose-fed rats during the immediate postnatal period to program for long-term metabolic health outcomes using phytochemicals. Neonatal administration of ursolic acid, 18 oleanolic acid, 19 and S-allyl cysteine 20 prevented the development of hepatic lipid accumulation in rats fed high-fructose diets. In the current study, we investigated whether zingerone could be used for that purpose.
Zingerone (vanillyl acetone) is an alkaloid found in the rhizomes of the plant ginger (Zingiber officinale). Studies have documented the hepatoprotective and antiobesity properties of zingerone in adult rat and mouse experimental models of metabolic disorders. 21 –24 The mechanisms by which zingerone exerts its hepatoprotective effects include its ability to upregulate lipolysis, scavenge free radicals, and downregulate inflammatory cytokines in the liver. 21,24
Given the heavy burden that management of NAFLD places on health care systems globally and the absence of specific treatments for NAFLD, there is need for strategic prophylactic interventions. The biological properties of zingerone made it a suitable candidate phytochemical in the current study for administration during the neonatal period to explore its potential to protect against long-term development of NAFLD secondary to high-fructose diets.
Materials and Methods
Experimental setting and procedures
The study was approved by the Animal Research Ethics Committee (AREC) of the University of the Witwatersrand (Clearance No: 2017/010/71/B) and conducted at the Central Animal Service (CAS) of the University of the Witwatersrand, Johannesburg, South Africa, using internationally accepted principles for laboratory animal use and care (South African National Standard [SANS 10386:2008] and Animals Protection Act, 1962: Act No. 71). The Animal Research Reporting of In Vivo Experiments (ARRIVE) guidelines were adopted while compiling the article.
The rats used in this study were obtained from the CAS, University of the Witwatersrand. Seven nulliparous dams of Sprague Dawley rats (Rattus norvegicus), each with a litter consisting of 8–12 four-day-old suckling male and female pups weighing 9.7 ± 1.1 g, were used in this study. The dams with their respective litters were housed in Perspex cages containing wood shavings as bedding. The room temperature was maintained at 25°C ± 1°C and a 12-h light and dark cycle with lights off from 7 pm to 7 am. The dams were supplied with standard rat chow (LabChef Rodent Breeder [Pty] Ltd., Stellenbosch, South Africa) and ad libitum water access throughout the suckling period. The pups were allowed to freely nurse from their respective dams until weaning on postnatal day 21. The dams were returned to stock after weaning of their pups. The weaned rats were then housed individually in Perspex cages having the same environmental conditions described earlier.
The study consisted of two main stages (Fig. 1): a preweaning stage (postnatal day 8–21) preceded by a 4-day adaptation period and a postweaning stage (postnatal day 22–91). On postnatal day 8, the pups (n = 79; 35 males, 44 females) from each litter were randomly allocated to four treatment groups: Group 1: received 10 mL/kg body weight (bwt) of distilled water gavaged orally before weaning (daily) and ad libitum access to plain drinking tap water after weaning (9 males, 11 females); Group 2: received 10 mL/kg bwt of a 20% fructose solution gavaged orally before weaning (daily) and ad libitum access to a 20% drinking fructose solution after weaning (9 males, 11 females) to induce metabolic dysfunction; Group 3: in which we investigated whether neonatal zingerone could program for protection against long-term high-fructose consumption, received 10 mL/kg bwt of a 20% fructose solution + zingerone at 40 mg/kg bwt gavaged orally before weaning (daily) and ad libitum access to a 20% drinking fructose solution after weaning (9 males, 12 females); Group 4: received 40 mg/kg bwt of zingerone dissolved in 10 mL/kg bwt of distilled water gavaged orally before weaning (daily) and ad libitum access to plain drinking tap water after weaning (8 males, 10 females) to assess the effects of zingerone alone on metabolic health of the animals.

Flow diagram showing the timeline and treatment groups.
Fructose solution at 20% has been shown to induce metabolic dysfunction in rats, 18 whereas zingerone at 40 mg/kg bwt has been shown to prevent against ethanol-induced hepatotoxicity in rats, 25 hence these doses were chosen for the current study.
The fructose solution (Natures Choice, South Africa) was constituted to 20% by adding 20 g of fructose into 100 mL of water, 26 whereas food-grade zingerone (Catalogue No. W312401-1KG) was purchased from Sigma–Aldrich.
The rats were weighed daily during the preweaning stage and then twice a week during the postweaning stage by individually placing the rats in a preweighed cage on an electronic balance (Snowrex Electronic Scale, Clover Scales, Johannesburg).
Procedures at termination
After the 12-week intervention period, the rats were fasted overnight, terminal body mass measured, and then euthanized with an appropriate and acceptable euthanasia in rats, an intraperitoneal injection of sodium pentobarbitone 27 (Centaur Laboratories, Johannesburg, South Africa) at 150 mg/kg bwt. After euthanasia, a ventral midline incision was made through the thorax and abdomen; syringes with attached hypodermic needles were used to withdraw blood samples via intracardiac access. The blood samples were then transferred into heparinized tubes (BD Vacutainer, Plymouth, United Kingdom) and centrifuged (Rotofix 32A; Hettich Zentrifugen, Germany) at 3700 g for 15 min at 20°C. The plasma was harvested and stored at −20°C for determination of ALT, uric acid, and TBARS. The liver was carefully dissected out from the abdominal cavity and weighed using a Precisa 310M balance (Precision Instruments, Johannesburg, South Africa). The right medial lobe of the liver was preserved in 10% phosphate-buffered formalin for histological analysis, while the remaining liver lobes were preserved at −20°C for quantification of lipid content.
Determination of hepatosomatic index
The terminal body mass and the fresh whole liver weight were used to compute the hepatosomatic index
28
:
Determination of hepatic lipid content
The hepatic lipids were determined by solvent extraction using a Soxhlet apparatus (Gebr. Rettberg GmbH, Göttingen, Germany), according to the Official Methods of Analysis of the Association of Official Agricultural Chemists (AOAC) International, 2005, Method 960.39. Briefly, the liver samples were lyophilized in a freeze-dryer (VirTis BenchTop BK-FD12; SP Scientific, New York) and milled into a fine powder. The milled liver samples were then extracted using petroleum ether in a Soxhlet apparatus and the lipid quantified gravimetrically.
Histological assessment of the liver
The right medial lobes of the liver samples that were preserved in 10% phosphate-buffered formalin were processed using an automatic tissue processor (Microm STP 120; Thermo Scientific, MA), embedded in paraffin wax, sectioned at 5 μm, and then stained with hematoxylin and eosin (H&E) for steatosis and inflammation assessment and Masson's trichrome (MT) for assessment of fibrosis. The slides were viewed under a light microscope (40 × ) (Leica Biosystems) mounted to a camera, which uses ZEISS ZEN microscope software for image capture, and scored for steatosis, collagen deposition, and fibrosis.
The H&E images were used to score micro- and macrovesicular steatoses as described by Kleiner et al. 29 The grading was performed based on the total area affected per camera field, according to the following criteria: Grade 0 steatosis: <5% fat droplets; Grade 1 steatosis: 5–33% fat droplets; Grade 2 steatosis: 34–66% fat droplets; and Grade 3 steatosis: >66% per camera field at 40 × magnification. Inflammation was scored by counting the foci of inflammatory cells in the liver parenchyma per camera field and graded as follows: Grade 0 = no foci of inflammation; Grade 1 = less than 2 foci; Grade 2 = 2–4 foci; Grade 3 = greater than 4 foci.
Fibrosis was assessed from photomicrographs of the MT-stained sections at 40 × magnification.
Determination of ALT, uric acid, TBARS, and inflammatory markers in plasma
Plasma ALT activity and uric acid concentration were determined using a calibrated VetTest analyzer (IDEXX VetTest® Clinical Chemistry Analyzer; IDEXX Laboratories, Inc.). TBARS, IL-6, and TNF-α were determined using rat-specific Enzyme-Linked Immunosorbent Assay kits (Elabscience Biotechnology Co., Ltd., Houston, TX), respectively.
Statistical analysis
Data were analyzed using GraphPad Prism 8 software (GraphPad Software, Inc., San Diego). Parametric data (hepatosomatic index, ALT, TBARS, IL-6, TNF-α, and hepatic lipid content) were analyzed by one-way analysis of variance (ANOVA) and expressed as mean ± standard deviation, followed by Tukey post hoc tests for comparison of the means. The Kruskal–Wallis test (nonparametric one-way ANOVA) was used to analyze multiple-group data for steatosis grades, inflammation, and fibrosis and expressed as median ± range, followed by Dunn's post hoc test to compare the medians. Sex variations were analyzed using two-way ANOVA. Statistical significance was considered when P < .05.
Results
Hepatosomatic index and hepatic lipid accumulation
The mass of the liver relative to body weight was not different (males: P = .6217; females: P = .0657) across the different treatment groups in both sexes (Table 1). No sex difference was observed between male and female rats regarding percent liver weight (sex effect: P = .9999, treatment effect: P = .4540, and interaction effect: P = .3500).
Effects of Neonatally Administered Zingerone on the Hepatosomatic Index and Hepatic Lipid Accumulation of High-Fructose Diet-Fed Rats
Data are expressed as mean ± standard deviation.
Row means with different letters (ab) significantly different at P = .0002 (males) and P < .0001 (females).
C, 10 mL/kg bwt of distilled water before weaning and free access to plain tap water after weaning (n = 20; 9 males, 11 females); Fr, 10 mL/kg bwt of a 20% fructose solution before weaning and free access to a 20% fructose solution after weaning (n = 20; 9 males, 11 females); ZFr, 10 mL/kg bwt of a 20% fructose solution combined with zingerone at 40 mg/kg bwt before weaning and free access to a 20% fructose solution after weaning (n = 21; 9 males, 12 females); Z, zingerone only at 40 mg/kg bwt in 10 mL/kg bwt of distilled water before weaning and free access to plain tap water after weaning (n = 18; 8 males, 10 females).
bwt, body weight.
Long-term fructose consumption significantly increased the hepatic lipid accumulation in both male (P = .0002) and female (P < .0001) rats compared with all other groups (Table 1). Thus, the neonatally administered zingerone prevented the fructose-induced hepatic lipid in both sexes, such that the zingerone-treated (with or without fructose) group had a similar (males: P = .7632; females: P = .5103) hepatic lipid content compared with negative controls. There was no sex difference observed (sex effect: P = .3774, treatment effect: P < .0001, and interaction effect: P = .8416) with regard to percent hepatic lipids accumulation.
Histomorphometry of the liver
Figure 2A and B shows representative photomicrographs of the liver histology from each of the experimental groups (H&E stain) of male and female rats, respectively. There was a notable fatty infiltration (macro- and microvesicular steatosis indicated by solid and dash arrows, respectively) in the sections of the fructose-only-treated group, which was generally absent in the other treatment groups (Fig. 2A [males]; Fig. 2B [females]). This was confirmed by the micro- and macrovesicular steatosis scores, which were greater in rats fed a high-fructose diet only compared with the other treatment groups in both sexes (P < .05, Dunn's multiple comparison; Table 2). This demonstrated that zingerone prevented the development of steatosis induced by high-fructose diet in the study rats. No difference was observed between males and females on the steatosis score for sex (P = .1889), treatment (P = .0826), and interaction (P = .7528) effects. Additionally, there were no observable inflammatory cell aggregates in both sexes.
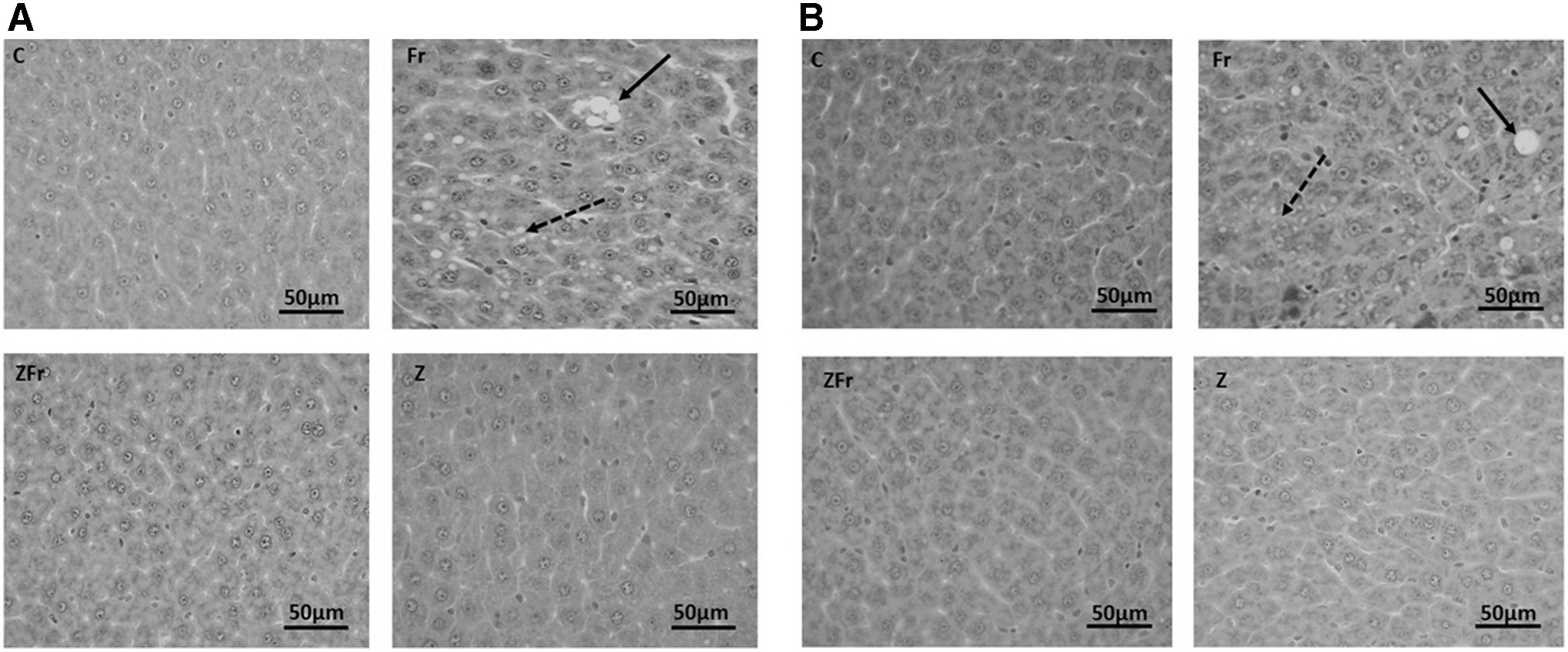
Representative photomicrographs of the male
Effects of Neonatally Administered Zingerone on the Steatosis Scores of High-Fructose Diet-Fed Rats
Data are expressed as median (interquartile range).
Row means with different letters (ab) significantly different at P < .05.
C, 10 mL/kg bwt of distilled water before weaning and free access to plain tap water after weaning (n = 20; 9 males, 11 females); Fr, 10 mL/kg bwt of a 20% fructose solution before weaning and free access to a 20% fructose solution after weaning (n = 20; 9 males, 11 females); ZFr, 10 mL/kg bwt of a 20% fructose solution combined with zingerone at 40 mg/kg bwt before weaning and free access to a 20% fructose solution after weaning (n = 21; 9 males, 12 females); Z, zingerone only at 40 mg/kg bwt in 10 mL/kg bwt of distilled water before weaning and free access to plain tap water after weaning (n = 18; 8 males, 10 females).
For the MT histological sections (MT stain), there was no observable fibrosis across the different treatment groups in both sexes (Fig. 3A [males]; Fig. 3B [females]).

Representative photomicrographs of the male
Plasma ALT activity and concentrations of uric acid, TBARS, and inflammatory markers
There were no differences in the activity of ALT (males: P = .0633; females: P = .2832), concentrations of uric acid (males: P = .5159; females: P = .7666), TBARS (males: P = .9889; females: P = .2282), IL-6 (males: P = .8979; females: P = .9867), and TNF-α (males: P = .3966; females: P = .4457) across all the treatment groups (Table 3). Males had higher ALT activity than females (sex effect: P < .0001, treatment effect: P = .0114, and interaction effect: P = .0626); otherwise no sex difference (P > .05) was observed in the levels of uric acid, TBARS, IL-6, and TNF-α for sex, treatment, and interaction effects.
Effects of Neonatally Administered Zingerone on the Concentrations of Alanine Aminotransferase, Uric Acid, Thiobarbituric Acid Reactive Substances, and Levels of Inflammatory Markers of High-Fructose Diet-Fed Rats
Data are expressed as mean ± standard deviation. P > .05.
Male rats had higher ALT activity than their female counterparts at P < .0001.
C, 10 mL/kg bwt of distilled water before weaning and free access to plain tap water after weaning (n = 20; 9 males, 11 females); Fr, 10 mL/kg bwt of a 20% fructose solution before weaning and free access to a 20% fructose solution after weaning (n = 20; 9 males, 11 females); ZFr, 10 mL/kg bwt of a 20% fructose solution combined with zingerone at 40 mg/kg bwt before weaning and free access to a 20% fructose solution after weaning (n = 21; 9 males, 12 females); Z, zingerone only at 40 mg/kg bwt in 10 mL/kg bwt of distilled water before weaning and free access to plain tap water after weaning (n = 18; 8 males, 10 females).
ALT, alanine aminotransferase; IL-6, interleukin-6; TBARS, thiobarbituric acid reactive substances; TNF-α, tumor necrosis factor alpha.
Discussion
The present study investigated the potential protective effects of neonatal administration of oral zingerone on long-term dietary fructose-induced fatty liver in Sprague Dawley rats. Hepatic lipid accumulation and fatty infiltration of hepatocytes were observed when the male and female rats were terminated in early adulthood, following long-term administration of the 20% fructose solution. Zingerone administered orally to the neonatal rats protected them against high-fructose diet-induced increased hepatic lipid accretion and steatosis (both sexes). Thus, neonatal administration of oral zingerone was effective in attenuating the development of long-term fructose-induced NAFLD in both male and female rats. A sexually dimorphic response was observed only in plasma ALT activity where male rats had more activity of the enzyme than their female counterparts.
The mass of the liver relative to body mass is known as the hepatosomatic index, 30 and it can be affected by nutritional status 31 and hepatic pathology including fatty liver diseases. 30 An increase in hepatosomatic index can result from a severe form of fatty liver disease including NASH, 30 whereas a decrease in hepatosomatic index can result from undernutrition. 31 In the current study, we observed an increase in intrahepatic lipid accumulation in the high-fructose-fed male and female rats, without affecting the hepatosomatic index. This finding is in agreement with Mamikutty et al. 12 who reported increased hepatic lipid deposition without an increase in liver weight in adult male Wistar rats that consumed 20% fructose solution for 8 weeks. The hepatosomatic index that was not affected could be that the amount of lipid vacuoles in the hepatocytes of the study rats had not reached the threshold for inducing changes in the liver weight. 12 This finding indicated that the observed steatosis was not severe enough to cause NASH that is usually associated with increase in hepatosomatic index. 30
However, the neonatally administered zingerone conferred protection against the long-term dietary fructose-induced hepatic lipids accretion in both sexes. A previous study also reported the hepatoprotective effect of zingerone in adult male rats, in which zingerone was administered orally at 100 mg/kg bwt for 8 weeks, and it prevented the intrahepatic lipid accumulation. 24 We have shown that by strategically targeting the neonatal period, a similar protective effect can be conferred. This beneficial effect of zingerone could have been due to its ability to enhance fatty acid oxidation via increase in hepatic PPAR activities, thereby attenuating hepatic lipogenesis. 24,25 In addition, zingerone could have prevented the fructose-induced hepatic lipid deposition (observed in this study) by attenuating hepatic de novo lipogenesis, which has been shown to be one of the deleterious effect of fructose on the liver. 13 Although excessive hepatic lipid content is of diagnostic importance for hepatic steatosis, the gold standard for diagnosis of NAFLD is histological assessment of hepatocytes. 32
Histologically, NAFLD primarily begins with fatty infiltration of hepatocytes when there are >5% steatotic liver cells in the histological sections of the liver. 5 In the present study, we observed the presence of both micro- and macrovesicular steatoses in both male and female rats that received the high-fructose diet only compared with the other groups. The observed fructose-induced steatosis in both sexes was prevented by neonatal administration of zingerone in this study, meaning that zingerone had programmed protection against the hepatic steatosis secondary to high-fructose diet consumption. The findings of this study are supported by those of a previous study in older rats, which reported that zingerone administered for 8 weeks (at 100 mg/kg bwt) prevented the fructose-induced steatosis in adolescent male Wistar rats. 24 It is thus likely that zingerone protected against accumulation of lipids in the liver by downregulating hepatic lipogenic genes and receptors including ChREBP, SREBP-1, and PPARα 24 since fructose has been reported to activate hepatic expression of these genes and receptors leading to increase hepatic lipid synthesis and storage. 14 However, the current study is limited in that we did not determine how these genes were expressed. Therefore, future studies on zingerone need to explore the molecular mechanisms involved in attenuating hepatic lipid accretion.
Additionally, zingerone has been reported to attenuate fructose-induced hepatic steatosis via decrease in hepatic pathways of fatty acids uptake, synthesis, and secretion. 24 This effect of zingerone could have been responsible for the prevention of hepatic steatosis observed in this study.
Despite the presence of hepatic steatosis in the study rats, no evidence of inflammation or fibrosis was observed in both male and female rats in this study. Thus indicating that the rats in the present study did not develop advanced fatty liver disease, which is typified by steatohepatitis and parenchymal tissue damage. 24 This has contrasted the findings of Narayanan and Jesudoss 24 who observed severe steatosis with associated inflammation and fibrosis following ad libitum consumption of 20% fructose solution by adolescent male Wistar rats for 8 weeks. The absence of inflammation and fibrosis in the histological samples of the study rats of both sexes could be explained based on the progressive nature of NAFLD, which begins with simple steatosis before inflammation or fibrosis manifest. 14 Additionally, the age of rats at commencement of the study could also have had an impact as younger rats are less susceptible to developing fructose-induced metabolic disorders than older ones. 33 It is also noteworthy that only a small percentage (10–15%) of patients with steatosis develop inflammation and fibrosis (NASH) later in life. 3 Fatty liver could be associated with hepatocellular damage and elevated levels of the hepatic function marker, ALT. 34
We did not observe any significant differences in the blood levels of ALT in rats in the different groups of both sexes. The lack of impact of treatments on the ALT is supported by our histological findings that did not show evidence of inflammation, which is invariably associated with elevated ALT. 12 This lack of elevation in ALT, secondary to a high-fructose diet, was also reported by a previous study after ad libitum consumption of 20% fructose solution by adolescent male Wistar rats for 16 weeks. 11 Studies have shown that 25% of histologically confirmed NAFLD may exist without elevation of ALT. 34 Thus, this enzyme is mainly used for the assessment of hepatic biochemical function and hepatic damage, but not to predict NALFD. 11
Despite ALT activity not being affected by the interventions in both sexes, we observed that the activity of ALT was higher in male rats than in female rats. It is not clear whether male sex hormones are responsible for the sex difference in the ALT activity. 35 However, since ALT is also produced by muscle cells, 35 and male rats usually have higher muscle mass than females, 36 we could speculate that the higher ALT activity observed in males could be due to their greater muscle mass when compared with females. Fructose-induced fatty liver is associated with elevated blood levels of uric acid 14 due to uncontrolled phosphorylation of fructose that converts ATP to adenosine monophosphate, which is eventually broken down to uric acid. 13
We observed that the plasma levels of uric acid were not different across the different treatments of both male and female rats in this study, despite the presence of steatosis in the hepatocytes of fructose-treated groups. In rodents, uric acid is converted to allantoin by the enzyme uricase, thereby decreasing circulating uric acid levels. 37 Humans normally do not have the enzyme uricase and are thus more prone to develop hyperuricemia than rats when exposed to triggering factors such as high-fructose diets. 37 Fructose-induced hyperuricemia is usually associated with severe forms of NAFLD than simple steatosis. 14 Additionally, Tapia et al. 38 demonstrated that severe metabolic derangement could be enhanced in rats when uricase is inhibited with oxonic acid. Thus, the findings of current study showing insignificant differences in uric acid concentrations could be explained by the effect of uricase in rats and the moderate NAFLD observed. Moreover, it has also been shown that fructose-induced fatty liver can develop with or without hyperuricemia. 39 Fructose-induced fatty liver can lead to oxidative stress secondary to lipid peroxidation 13 and subsequent hepatic inflammation. 40
Although zingerone has been reported to prevent hepatic damage by inhibiting the generation of free radicals and downregulating inflammatory cytokines of hepatic inflammation, 23 –25 the treatments had no effect on the levels of TBARS, IL-6, and TNF-α in male and female rats in the present study. Fakhoury-Sayegh et al. 11 also reported similar findings when 20% fructose solution was consumed ad libitum by adolescent male Wistar rats for 16 weeks. These findings suggested that the interventions did not cause an overt proinflammatory state, a finding that can be reinforced by histological features of the rats' hepatocytes as there were no evident inflammatory foci, further confirming the presence of a nonsevere form of the disease that had not progressed to NASH.
In conclusion, neonatal administration of zingerone orally to Sprague Dawley rats prevented the long-term fructose-induced fatty liver development in both sexes. A common mantra among health care professionals is that “prevention is better than cure.” Targeting this critical period of developmental plasticity, with a naturally derived phytochemical may provide an alternative prophylaxis for NAFLD and hence reduce the burden of this condition on health care facilities globally. Future studies will be directed toward understanding the mechanisms of action of zingerone that resulted in these long-term protective effectives and the possible transgenerational benefits of the interventions. Additionally, the interaction of fructose with micronutrients such as copper will also be considered in future studies since fructose has been shown to alter the utilization of copper. 41
Footnotes
Authors' Contributions
The study was designed and conducted by N.M., B.W.L., and K.H.E. N.M. produced the first draft of the article, and all the authors contributed in writing and revising the article. The authors have also consented to the article submission.
Acknowledgments
Staff of the Central Animal Service Unit, Faculty of Health Sciences, University of the Witwatersrand are hereby acknowledged for the care of the animals. Miss Monica Gomes and Miss Hasiena Ali are also thanked for their technical assistance with the histology.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
We acknowledge the Medical Faculty Research Endowment Fund, Faculty of Health Sciences Research Committee and School of Physiology of the University of the Witwatersrand (Grant No. 001 254 8521101 5121105 000000 0000000000 5254); the National Research Foundation (NRF) Thuthuka Fund (Grant No. TTK170415227205) that provided funds for the research. Federal University Birnin Kebbi (Nigeria) and Tertiary Education Trust Fund of Nigeria are hereby acknowledged for providing funds for support of the PhD candidate.