
Abstract
Cardiac fibrosis is one of the hallmarks of a diabetic cardiomyopathy. When activated, cardiac fibroblasts (CFs) increase the production of extracellular matrix proteins. Transforming growth factor (TGF)-β1 is known to mediate cardiac fibrosis through the SMAD pathway. High glucose (HG = 25 mM) cell culture media can activate CFs using TGF-β1. There is a need to identify effective antifibrotic agents. Studies in animals indicate that treatment with (−)-epicatechin (Epi) appears capable of reducing myocardial fibrosis. Epi binds to G-protein coupled estrogen receptor (GPER) and activates downstream pathways. We evaluated the potential of Epi to mitigate the development of a profibrotic phenotype in HG stimulated CFs. CF primary cultures were isolated from young male rats and were exposed for up to 48 h HG media and treated with vehicle or 1 μM Epi. Relevant profibrotic end points were measured by the use of various biochemical assays. HG exposure of CFs increased TGF-β1 protein levels by ∼15%, fibronectin ∼25%, urea levels ∼60%, proline incorporation ∼70%, and total collagen ∼15%. Epi treatment was able to significantly block HG induced increases in TGF-β1, fibronectin, urea, proline, and total collagen protein levels. GPER levels were reduced by HG and restored in CFs treated with Epi an effect associated with the activation (i.e., phosphorylation) of c-Src. Epi treatment also reverted SMAD levels. Altogether, results demonstrate that CFs cultured in HG acquire a profibrotic phenotype, which is blocked by Epi an effect, likely mediated at least, in part, by GPER effects on the SMAD/TGF-β1 pathway.
Introduction
Tissue fibrosis is frequently the end result of chronic inflammatory reactions induced by a variety of stimuli leading to the excess deposition of connective tissue that progressively remodels and replaces the normal cellular architecture. 1,2 Fibrosis is an important component of a pathologically remodeled heart and contributes to the loss of chamber function. 3 With type 2 diabetes mellitus (DM), extensive tissue remodeling and fibrosis can occur in the absence of atherosclerosis leading to the development of diabetic cardiomyopathy. 4
The cardiac fibroblast (CF) is the cell type that produces the majority of the myocardial extracellular matrix (ECM) proteins, including fibronectin and fibrillar collagen types I and III. 5 In the setting of stimuli such as mechanical overload or following a myocardial infarction, CFs undergo a phenotypic modification giving rise to a profibrotic cell type, which is also termed a myofibroblast. 6 A major central orchestrator of fibroblast activation and, consequently, of profibrotic activity is transforming growth factor (TGF)-β1. 6 This cytokine mediates cardiac fibrosis associated with processes such as hypertension and DM through the modulation in CFs of the SMAD signaling pathway. 1,7 High glucose (HG) blood levels as seen with DM is recognized as a promoter of cardiac fibrosis. When isolated, CFs exposed to HG cell culture media (i.e., simulating DM conditions) can acquire a profibrotic phenotype and increase ECM protein synthesis. 3,8
So far, most of the pharmacological strategies tested to mitigate tissue fibrosis have yielded poor outcomes in clinical trials 9 ; thus, the urgent need to identify potential candidates to treat tissue fibrosis. (−)-Epicatechin (Epi) is the most abundant flavonol found in cacao, and its regular consumption is associated with reduced cardiovascular disease risk through its vasodilatory actions and improved cellular bioenergetics. 10 In animal models of myocardial injury, we have reported on the apparent capacity of Epi to reduce tissue injury and/or fibrosis. 11 –13 In endothelial and skeletal muscle cells, Epi can bind the G-protein coupled estrogen receptor (GPER) and activate downstream signaling pathways 14,15 leading to effects such as enhanced nitric oxide production. However, nothing is known about a possible role of GPER in CFs and potential of Epi to reduce HG stimulated ECM production.
Therefore, the purpose of this study was to examine the potential of Epi to mitigate the development of a profibrotic phenotype in HG stimulated CFs through GPER modulation of the TGF-β1 signaling pathway.
Materials and Methods
Materials
Three-month-old Sprague-Dawley male rats were obtained from Envigo, Inc. All procedures were approved by University of California, San Diego's Institutional Animal Care and Use Committee (Protocol No. S19042) and conform to published NIH guidelines for animal research. Collagenase was obtained from Worthington Biomedical Corporation (Lakewood, NJ), Epi and pancreatin were from Sigma-Aldrich, Inc., and cell culture media, trypsin, fetal bovine serum (FBS), and antibiotics were from Gibco BRL (Thermo Fisher Scientific). Antibodies used were as follows: p-SMAD 2/3 (No. PA5-36028, 1:1500 working dilution; Invitrogen), SMADS 2/3 (Catalog No. 3102, 1:1000 working dilution; Cell Signaling), SMAD 7 (MADH7, Catalog No. 190987, 1:2000 working dilution; Abcam), p-c-SRC (No. 2105, 1:1000 working dilution; Cell Signaling), c-SRC (No. sc-18, 1:500 working dilution; Santa Cruz Biotechnology), and GPER (No. PA5-87717, 1:1000 working dilution; Invitrogen) and to normalize for loading differences ribosomal subunit 6 protein (S6RP, No. 23175, 1:1000 working dilution; Cell signaling). All other reagents were of cell culture and/or molecular biology grade.
Cell culture
Primary adult rat CF cultures were isolated from hearts as previously described.
8
Briefly, rats were sacrificed by CO2 asphyxia, and hearts were quickly removed under sterile conditions. Ventricular tissue was isolated and digested using a Langendorff system and an enzymatic solution of collagenase (100 U/mL) and pancreatin (0.6 mg/mL). Isolated CFs were pooled together, centrifuged, and resuspended in growth media containing Dulbecco's modified Eagle's media (DMEM) (EDTA, pH 7.4), 5.5 mM
Treatment
CFs were grown to ∼80% confluency in growth media. CFs were rendered quiescent by serum starving for 24 h in phenol red free DMEM supplemented with 1% FBS and 1% PSF. CFs were then treated for 48 h with DMEM 0.5% FBS, 1% PSF using either NG or HG (25 mM
Total protein extraction
Cells were washed thrice with cold buffer (4 mL/plate) and lysed in 80 μL of ice-cold lysis buffer with protease and phosphatase inhibitor cocktails. Homogenates were sonicated for 15 min at 4°C and centrifuged at 13,000 g for 15 min to remove cell debris. The total protein concentration was measured in the supernatant using the colorimetric Bradford micro method (Bio-Rad, Inc.) with readings at 595 nm wavelength using a BioQuant 800 spectrophotometer (BioTek, Inc.)
TGF-β1 and fibronectin ELISA
TGF-β1 and fibronectin were quantified in cell lysates using competitive ELISA Kits following supplier instructions for TGF-β1 (Catalog No. 88-50390-22; Thermo Fisher) and fibronectin (Catalog No. EK0350; ScienCell).
Hydroxyproline quantification
A Hydroxyproline Assay Kit (Catalog No. MAK008; Thermo Fisher, Inc.) was used to determine the total amount of collagens synthetized following manufacturer's instructions. A total of 30 μg of cell lysates were hydrolyzed and stained to determine the hydroxyproline content of the samples with values recorded using a microplate reader at a wavelength of 550 nm. Total collagen protein levels were estimated according to the method reported by Cissell et al. 17
Western blots
To examine relative protein levels of SMAD2/3, p-SMAD2/3, SMAD7, c-Src, p-c-Src, GPER, and S6RP, immunoblotting was performed. Cells were treated as described above. After treatment, cells were lysed and total protein extracted. A total of 30 μg of protein was loaded onto a 4–15% gel and electrotransferred onto a membrane followed by either 1–3 h incubation at room temperature or overnight incubation at 4°C with primary antibodies. Primary antibodies were diluted in buffer plus 5% bovine serum albumin or 2% nonfat milk. Membranes were washed (three times for 5 min) in buffer and incubated for 1 h at room temperature in the presence of conjugated secondary antibodies diluted 1:5000 in a blocking solution. Membranes were again washed thrice in buffer, and immunoblots were developed. Band intensities were digitally quantified and normalized against S6RP.
Proline incorporation
[ 3 H] proline incorporation was used as an indicator of total collagen synthesis. CFs were plated in 24-well tissue culture dishes and grown to 80–85% confluence. Cells were incubated as described above. During the last 36 h of treatment, CFs were pulsed with [ 3 H] proline (μCi/mL). To end the experiment, each well was washed twice with cold buffer solution followed by the addition of cold 10% trichloroacetic acid (TCA; 500 μL/well) to lyse the cells and precipitate cellular proteins. Wells were washed thrice with TCA. NaOH (250 μL, 1 N) was added to each well to solubilize proteins. Samples were neutralized with 250 μL of 1 N HCl for 30 min and radioactivity counted after the addition of scintillation fluid.
Urea quantification
The urea concentration was calculated as an indirect measure of synthetized proline. Urea was determined as an end product of arginase activity. After treatment, cells were homogenized in 100 μL of solution A (sucrose 2 M, EDTA 0.01 M, HEPES 0.5 M; pH 7.4) and samples centrifuged for 10 min (12,000 g) at 4°C. The supernatant was collected, and protein concentration on it was determined using the Bradford method. To evaluate arginase activity, 100 μg of protein was added to 25 mM Tris-HCl pH 7.4 and 5 mM MnCl2 (100 μL final volume) and activated at 55°C for 10 min. The enzymatic reaction was started by the addition of 100 μL of 0.5 M
Wound healing assay
To calculate migration rate, passage 2 cell cultures were seeded in six well plates. CFs were grown to full confluency in growth media. A starvation and treatment period were rendered as described above. Four hours after treatment started, using a 10 μL pipette tip a scratch was made to create a gap between cells. Pictures of the wound closure were taken over a 36-h time period at three different points in each well. The decellularized area was calculated using ImageJ software.
Cell number
To assess for changes in cell proliferation, cell counting was performed over a 36 h period after initially plating at ∼60% confluency. For this purpose, 10,000 cells were seeded in 24-well plates. A starvation and treatment period were rendered as described above. At 0, 12, and 36 h after treatment, two wells per group were trypsinized and cells counted using Countess™ II FL automated cell counter (Catalog No. AMQAF1000; Thermo Fisher Scientific) following the device instructions.
Statistical analysis
All data are shown as mean ± standard error of the mean. Statistical analysis used ANOVA (Dunn's post-test). Analysis was done using Prism software (GraphPad, CA, USA). Results were deemed significant when P ≤ .05.
RESULTS
The quantification of total protein concentrations demonstrated no significant differences between the NG, HG, or HG-Epi groups (Fig. 1A). As shown in Figure 1B and C, HG demonstrated a trend toward stimulating increases in TGF-β1 by ∼16% while significantly increasing those of fibronectin by ∼25%. Pretreatment with 1 μM Epi was able to significantly block HG induced increases in TGF-β1 (Fig. 1B) with reductions of ∼23% and of fibronectin (Fig. 1C) of ∼30%. Indicators of collagen synthesis and deposition are noted in Figure 2 and demonstrate a ∼60% increase of urea levels (Fig. 2A), ∼70% in [ 3 H] hydroxyproline incorporation (Fig. 2B), and ∼15% in total collagen content (Fig. 2C) in HG groups. Treatment with Epi was able to significantly reduce HG-induced increases in urea levels by ∼50% and in [ 3 H] hydroxyproline incorporation and total collagen where their levels became comparable to those of NG.
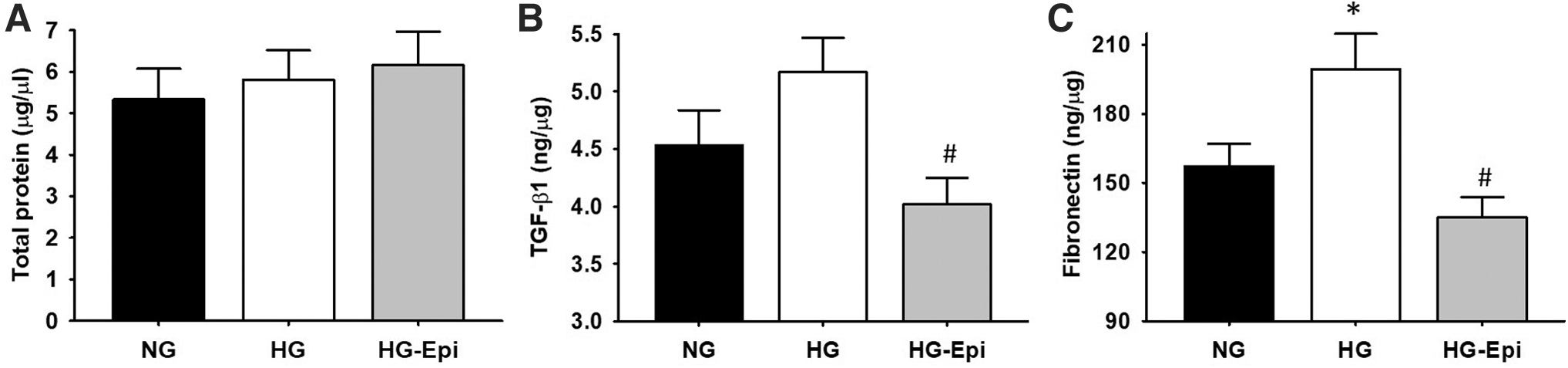
Effects of (−)-Epi treatment (1 μM) on HG media cultured cardiac fibroblasts on total protein content

Effects of (−)-Epi treatment (1 μM) on collagen synthesis and total collagen content in HG cultured cardiac fibroblasts.
We also explored the effects of HG and Epi on CF proliferation. Cell numbers at 36 h post-treatment were comparable in all groups (NG = 4.11 × 105 ± 3.82 × 104, HG = 4.77 × 105 ± 5.66 × 104, and HG-Epi = 4.02 × 105 ± 7.36 × 104); thus, HG did not demonstrate an ability to modify CF proliferation. Results from the assessment of CF migration are reported in Figure 3. As can be observed, no significant differences were detected at any of the times examined with HG on CF migration and Epi did not modify these end points.

Analysis of HG ±1 μM (−)-Epi effects on cardiac fibroblast migration as determined by a wound healing assay.
TGF-β1 mediated modulation of the SMAD pathway occurs through increases in SMAD 2/3 levels and/or activity (i.e., phosphorylation) and inverse effects on SMAD 7 protein levels. Results are reported in Figure 4A–E with representative Western blot images shown in Figure 4A. HG media was able to significantly increase SMAD 2/3 phosphorylation (Fig. 4B) and the total levels of SMAD 2/3 protein (Fig. 4C). Figure 4D reports on the levels of phosphorylated SMAD 2/3 as normalized to the total protein levels yielding an increase of ∼100%. HG also led to the suppression of SMAD 7 protein levels by ∼35%. With Epi, CFs demonstrated a suppression of SMAD 2/3 activity toward those observed in NG cells and full recovery of SMAD 7 levels to become comparable to those of NG. Figure 5A–E reports on the effects that treatment yielded on GPER protein levels and its associated signaling pathway (c-Src) with representative images from Western blots shown in Figure 5A. As shown in Figure 5B, GPER is expressed in CFs, and its protein levels were suppressed by HG media showing a ∼35% decrease, which was partially restored with Epi treatment. Binding of Epi to the GPER has been associated with the activation of c-Src, which is known to interact downstream with the SMAD pathway. 14,18 As shown in Figure 5E, CFs treated with HG-Epi denote a doubling of c-Src phosphorylation (i.e., activation) versus NG or HG levels when values were normalized for total protein levels (Fig. 5C, D), which remained stable with HG or Epi treatment.
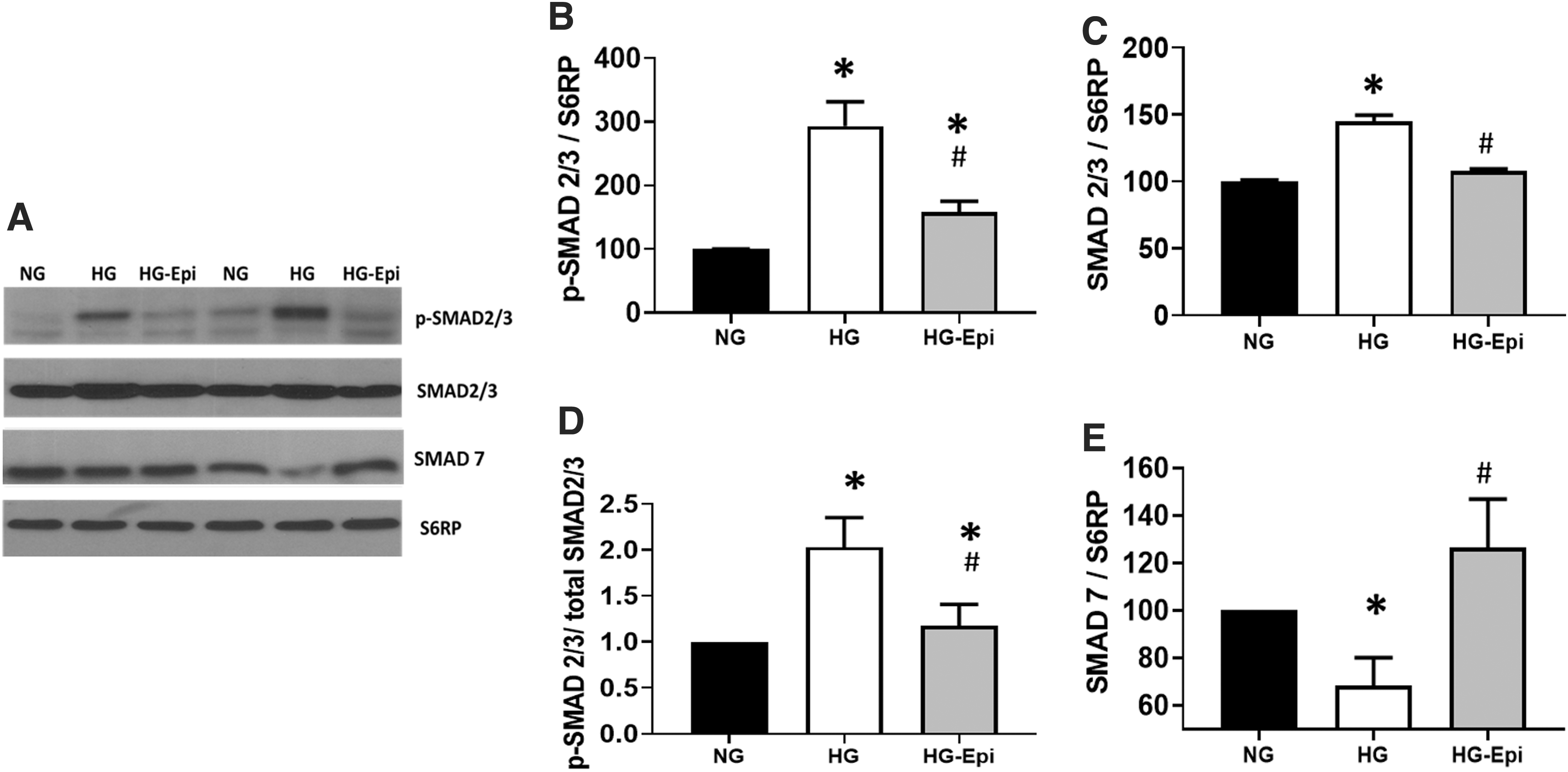
Effects of HG ±1 μM (−)-Epi treatment on the SMAD pathway in cardiac fibroblasts. Images of relative protein levels assessed by Western blot

Effects of HG ±1 μM (−)-Epi treatment on GPER expression and c-SRC activation in cardiac fibroblasts. Images of relative protein levels assessed by Western blot
Discussion
Various classes of antifibrotic candidate drugs have been evaluated with limited success in the setting of adverse cardiac remodeling and fibrosis and include those that target the renin angiotensin aldosterone system, β-blockers, cross-link breaker compounds, modulators of inflammation, and agents such as relaxin. 9 Drugs tested both in the preclinical and clinical setting include angiotensin converting enzyme inhibitors, AT1 receptor blockers such as losartan, and aldosterone blockers such as spironolactone or eplerenone. 9 While initial results from small clinical studies appeared promising, subsequent larger trials failed to validate them as effective antifibrotic agents. Currently, only two compounds, pirfenidone and nintedanib, are approved for the treatment of organ fibrosis specifically of idiopathic pulmonary fibrosis. 19 Unfortunately, the side effects of both compounds are such that their long-term use is highly problematic. Thus, the need to identify novel candidate compounds in particular those that may be well tolerated over the long term and carry an acceptable safety profile.
Natural products are recognized as a reliable source of therapeutic compounds as ∼46% of the drugs approved by the FDA between 1981 and 2014 are derived from the original source or their derivatives. 19 As such, natural products continue to be an attractive source of molecules with potential antifibrotic action. A series of recent reviews comprehensively address the topic and summarize those natural products that have been tested in preclinical models of organ fibrosis. 19 –21 The review by Zhang et al., focuses on products that have been evaluated for the possible treatment of cardiac fibrosis using either cell culture models and/or in vivo (rodent) models. 21 Forty-one studies published since 2009 are included in a table and are presented by flavonoid/other subcategories, including flavanols (e.g., quercetin), flavones (e.g., genistein), flavanones (e.g., hesperetin), nonflavonoids (e.g., resveratrol), and others (e.g., curcumin). A summary of the main actions observed is also included where suppressive effects over TGF-β-1 and related signaling pathways become prominent. On the basis of the evidence provided it becomes readily apparent that natural products have the potential to mitigate the development of cardiac fibrosis. However, none of the review articles report on the implementation of clinical trials where such actions may be tested.
Epidemiological reports indicate that the consumption of modest amounts of cocoa products rich in Epi (such as dark chocolate) has been associated with the reduced incidence of cardiovascular diseases such as hypertension, myocardial infarction, and heart failure. 22 Steinhaus et al., reported on the association between chocolate consumption and heart failure in a large (31,917) population of men in Sweden. 23 In this large prospective cohort study, there was a positive relationship between moderate chocolate consumption and a lower rate of heart failure hospitalization or death. These observations were confirmed in a population study in women (38,823 in the Swedish Mammography Cohort) and physicians (20,278 in the Physicians Health Study). 24,25 Thus, there is strong suggestive epidemiological evidence supporting the cardiovascular protective effects of cocoa. Currently, a double blind, randomized, placebo-controlled trial (COSMOS) is taking place in ∼22,000 subjects to be treated for 4 years with a high flavanol cocoa extract examining the effect of treatment on heart disease, stroke, and cancer incidence. 26
In various animal models of organ damage and/or adverse remodeling, we reported on the capacity of Epi to ameliorate tissue injury and preserve organ structure. 11 –13 Using rat models on infarction, we reported on the cardioprotective effects of Epi. In a mouse model of severe muscular dystrophy (δ-sarcoglycan null mice), Epi treatment was able to reduce skeletal and cardiac muscle fibrosis. 27 Antifibrotic actions of Epi were also detected in the left ventricle of an aged, female rat model of pre-heart failure with preserved ejection fraction. 28 Interestingly, in a recent pilot clinical trial providing 100 mg/day of Epi for 8 weeks to Becker muscular dystrophy patients, we reported on improvements in multiple indicators of heart (heart rate, maximal oxygen consumption, lactate levels, tissue saturation index) and skeletal muscle health (regulators of muscle differentiation and growth). 29 However, it is not clear if reductions detected in muscle fibrosis follow reduced levels of tissue injury and/or direct antifibrotic actions.
Estrogens are well known to confer cardiovascular disease protection to premenopausal women. Intriguingly, estrogens also appear to confer to premenopausal women, a high degree of protection toward the development of organ fibrosis. 30 The antifibrotic effects of estrogens appear to occur in most organs, including kidney, lung, liver, and heart. It is known that estrogen effects are mediated through three families of receptors, which include the ERα, ERβ, and GPER. 31 GPER is a membrane-bound receptor that binds estradiol with high affinity but signals differently from ERα and ERβ. 31 Noteworthy, 9-month-old female cardiomyocyte-specific GPER null mice display increased heart weight and cardiac collagen deposition among other detrimental changes. 32 The expression of GPER in cultured rat CFs was recently reported, and use of the GPER agonist G1 leads to the inhibition of CF cell proliferation. 33 Furthermore, in studies using G1 treatment of mouse and rat models of cardiac remodeling and fibrosis induced by aging, ovariectomy, infarction, pulmonary hypertension, and heart failure, a common outcome was reduced myocardial fibrosis. 31 Thus, the presence and/or activation of GPER is associated with lessened degrees of myocardial fibrosis.
We recently reported that Epi can bind and activate GPER in endothelial and skeletal muscle cells leading to effects such as increased production of nitric oxide and stimulation of mitochondrial biogenesis. 14,34 Using in silico studies, we reported on the favorable binding between Epi and GPER in endothelial cells. 14 The binding was comparable to that of the GPER agonist, G1. We also demonstrated that in endothelial cells, the activation of downstream pathways was dependent on the activation of the GPER/c-Src/ERK1/2 axis mimicking those effects reported with G1. 14 It has been reported that in oncogenic cell lines ERK1/2 activation counters the effects of TGF-β activity as it can decrease TGF-β receptor levels, increase the level of SMAD corepression, and attenuate SMAD accumulation in the nucleus. 18 We previously reported that CFs acquire a profibrotic phenotype when exposed to HG media 3 and that these effects are associated with TGF-β1 pathway activation through increases in stimulatory SMAD 2/3 and reductions in inhibitory SMAD 7 protein levels. 8 As noted above, there is extensive precedent for the participation of TGF-β1 in stimulating the development of cardiac fibrosis and that its suppression can lead to its mitigation. Such effects have been reported in studies using natural products such as quercetin, resveratrol, and curcumin. 21 These results are in line with our findings where Epi treatment was able to reduce HG-induced production of ECM proteins through the inhibition of the TGF-β1-SMAD pathway.
We and others have also reported on the capacity of various candidate antifibrotic agents to inhibit HG or angiotensin II stimulated CF functions. We previously reported that angiotensin II stimulates CF TGF-β1 production through the AT1 receptor and effect blocked by losartan. 35 In CFs, losartan blocks HG induced increases in CF protein and collagen synthesis. 3 In neonatal rat CFs incubated with HG, relaxin reduces cell proliferation. 36 In a study by Cai et al., epigallocatechin-3-gallate, the most abundant polyphenol found in green tea, significantly reduced collagen synthesis, fibronectin expression, and cell proliferation in rat CFs stimulated with angiotensin II. 37 Chen et al., reported that resveratrol treatment inhibits mouse CF proliferation and fibroblast-to-myofibroblast transition stimulated by angiotensin II through a suppression of the TGF-β/SMAD pathway. 38 In human CFs, curcumin can inhibit HG induced increases in collagen production through the inhibition of the TGF-β1 and SMAD 2/3 pathway. 39
Taking into consideration our results and those noted above by others, Figure 6 summarizes a potential explanatory depiction on how Epi-GPER activation in CFs suppresses a HG-induced profibrotic phenotype. In this scenario, Epi induced GPER activation of c-Src leads to the downstream inhibition of the TGF-β1/SMAD pathway and reductions in ECM synthesis. However, it is worth noting that the involvement of other receptors cannot be excluded as Epi is known to bind to other entities 40 and c-Src can also be activated through other receptors. 41
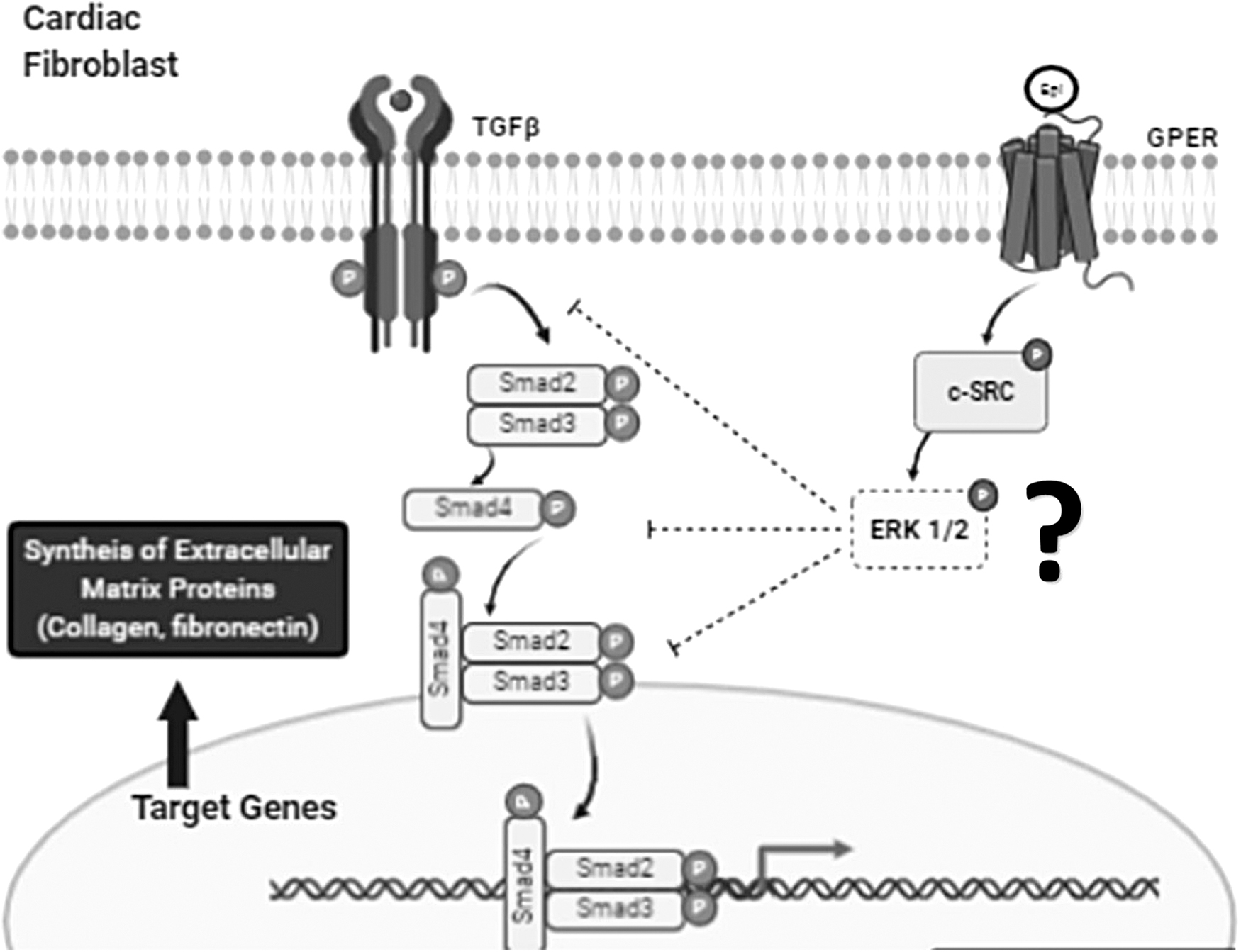
Proposed antifibrotic mechanisms of (−)-Epi. Binding of (−)-Epi to GPER phosphorylates s-Src leading to the downstream activation of pathways such as ERK1/2 that can inhibit extracellular matrix protein production through the suppression of the TGF-β1 pathway (image generated with BioRender.com).
In conclusion, there is in vivo evidence that Epi may exert antifibrotic actions but the potential direct mechanisms modulating CF phenotype are poorly understood. This study provides evidence for the potential role played by CF GPER and the suppression of TGF-β1 action by Epi. As Epi is known to be safe and well tolerated by humans, its antifibrotic potential warrants further validation in clinical trials.
Footnotes
Author Disclosure Statement
F.V. is a cofounder and stockholder (G.C.) of Epirium Bio, Inc.
Funding Information
DoD PR150090, NIH DK98717, AG47326, VA I01BX3230 to F.V.