
Abstract
Nonalcoholic fatty liver disease (NAFLD) is a metabolic liver disease with a complex underlying mechanism that has not been completely understood. Thus, effective and safe drugs for this disease are not yet available. Artemisia annua L. is a medicinal plant with potent antimicrobial and antioxidant activities. In this study, we prepared a water extract of A. annua (WEAA) and examined its potential for NAFLD treatment. First, we pretreated HepG2 cells (human hepatocarcinoma cell line) with WEAA and then treated the cells with oleic acid or tert-butylhydroperoxide to examine the effect of WEAA on the lipid accumulation and the cytotoxicity, respectively. WEAA not only inhibited lipid accumulation within HepG2 cells but also protected cells from oxidative stress-mediated damage through the activation of antioxidant enzymes (such as activation of superoxide dismutase and production of glutathione) and its own scavenging activity. Next, to confirm protective effect of the WEAA in in vivo, mice were intragastrically administered with WEAA, extract of Silybum marianum or water once a day, and simultaneously provided with high-fat diet to induce fatty liver and hepatic steatosis. Oral administration of WEAA ameliorated weight gain and hepatic lipid accumulation in high-fat diet-fed mice. Moreover, the plasma levels of triglyceride, aspartate aminotransferase, and alanine aminotransferase were reduced in the WEAA-treated group. Our findings indicated that WEAA may be a potential intervention for preventing or treating hepatic lipid accumulation and liver damage.
INTRODUCTION
Nonalcoholic fatty liver disease (NAFLD) is a hepatic symptom of metabolic syndromes. 1 The incidence of NAFLD is increasing in proportion to the prevalence of obesity, diabetes, and hyperlipidemia; moreover, it is associated with major adverse cardiovascular events, including cardiovascular diseases, coronary artery diseases, hypertension, and atherosclerosis. 2,3 Thus, NAFLD is being recognized as a major public health problem. However, only a small minority of patients with NAFLD deteriorate into fibrosis, cirrhosis, and hepatocellular carcinoma. 4,5 The underlying mechanism of the development and progression of NAFLD is complex and multifactorial, and thus remains unclear. 1 Its pathogenesis involves excessive free fatty acid (FFA) level in the liver, which leads to not only triglyceride accumulation but also lipotoxicity. 6 The increased toxicity causes mitochondrial dysfunction, reactive oxygen species (ROS) generation, and endoplasmic reticulum stress related to accumulation of unfolded proteins, which promote liver damage. 6,7
As an effective and safe drug for long-term use is not yet available for the treatment of NAFLD, physicians currently recommend losing body weight through dietary and exercise regimens to patients as a primary therapeutic approach. 8,9 Several candidate drugs, including metformin, statins, and fibrates, are currently being developed as anti-NAFLD agents in the clinical trial stage. 10 However, many candidates are already known to exert adverse side effects, such as high risk of infection and osteoporosis. 11 –13 Thus, the development of novel agents with high efficacy and safety is required for the therapy and prevention of NAFLD.
Artemisia annua L., called mugwort, is an annual short-day herb belonging to the Asteraceae family. It is distributed throughout countries in America, Europe, and Asia, including Korea and China. 14,15 A. annua has been traditionally used as a treatment of mild fever, hemostasis, and skin diseases, as well as used as a pesticide. 16 –19 Many studies had reported its antibacterial, antiviral, and antioxidant activities. 20,21 Among its components, artemisinin, a sesquiterpene lactone with an endoperoxide bridge, is well known to inhibit the activity of Plasmodium falciparum and other malaria parasites by inducing alkylation of malaria-specific proteins. 21,22 Recently, its compounds have been shown to have anticancer activity by selectively inducing necrosis in breast cancer cells. 23 –25 Moreover, artemisinic acid, another compound isolated from A. annua, is able to inhibit adipogenic differentiation of human adipose tissue-derived mesenchymal stem cells. 26 Other mugworts have been reported to have beneficial effects in high-fat diet-induced obesity. 27,28
In this study, we examined the effects of a water extract of A. annua L. (WEAA) on various parameters related to fatty liver in HepG2 cells and in high-fat diet-fed mice. Our results showed that WEAA ameliorated hepatic lipid accumulation and hepatotoxicity in vitro, as well as weight gain and liver damage in vivo.
MATERIALS AND METHODS
Preparation of plant extracts
A. annua L. was purchased from commercial markets in Korea and was extracted with water in an extractor at 85°C for 4 h. The extracts were filtered with 10 μm cartilage filter and the filtrate was concentrated to 25% of solid contents under 60°C reduced pressure. The concentrate was powdered with the addition of 30% dextrin using a spray dryer. An extract of Silybum marianum (ESM) 29 with a silymarin content of 50% and approved by the Ministry of Food and Drug Safety was purchased from a commercial market and used as a positive control in the experiments.
Cell culture
The human hepatocarcinoma cell line HepG2 was purchased from American Type Culture Collection (USA). HepG2 cells were grown in Dulbecco's modified Eagle's medium supplemented with 10% fetal bovine serum and 1% (v/v) penicillin-streptomycin solution. All cell culture products as mentioned above were from GE Healthcare Life Science (USA). Cell culture was sustained at 37°C in a 5% CO2-humidified incubator.
Oil red O staining and measurement of lipid accumulation
HepG2 cells were seeded at a density of 4 × 105 cells/mL in a six-well plate. At 24 h of incubation, the cells were pretreated with 0.0625–1.0 mg/mL of WEAA for 3 h. After that, the WEAA-containing media were washed out and the cells were further incubated with 1.5 mM oleic acid (Sigma, USA) for 24 h to induce lipid accumulation. Oleic acid was prepared as 10 × stock solution (15 mM) by dissolving oleic acid powder (Sigma) into prewarmed culture medium by vigorous vortexing. Before use, the stock solution was diluted with culture medium to make the final concentration of 1.5 mM. To measure accumulated lipid within the cells, cells were stained with Oil red O solution (Sigma) according to the manufacturer's protocol. In brief, lipid-accumulated cells were washed with phosphate-buffered saline (PBS), and fixed with 4% paraformaldehyde for 30 min. The cells were subsequently washed with water and stained with Oil red O solution in a mixture of isopropanol and water (3:2) for 20 min, followed by washing with water and 60% isopropanol for three times. Finally, the stained cells were eluted with 1 mL of 100% isopropanol, and 200 μL of the eluates was used for measurement of optical density at a wavelength of 492 nm.
Viability assay
HepG2 cells were seeded at a density of 1 × 105 cells/mL in a 96-well plate. After 24 h of incubation, the cells were treated with various concentrations (0.0156–1.0 mg/mL) of WEAA for 3 h, and then exposed to 0.7 mM of tert-butylhydroperoxide (t-BHP; Sigma) for an additional 3 h to induce oxidative stress. The media were replaced with fresh media supplemented with 0.5 mg/mL thiazolyl blue tetrazolium bromide (MTT; AppliChem, Germany) for 4 h in the dark at 37°C. To dissolve the insoluble formazan, dimethyl sulfoxide was added to each well after the media was suctioned. The absorbance was measured at 540 nm using an ELISA reader (SpectraMax M2/M2e Microplate Readers; Molecular Devices, USA).
Measurement of aspartate aminotransferase and superoxide dismutase activities and glutathione level
HepG2 cells were plated at a density of 1 × 105 cells/mL in a 100-mm dish. After 24 h of incubation, the cells were pretreated with 0.0156–1.0 mg/mL of WEAA in Dulbecco's modified Eagle's medium without phenol red (GE Healthcare Life Science) for 3 h, and then the cells were treated with t-BHP at a final concentration of 0.7 mM for another 3 h. The cells were harvested and then lysized with 300 μL lysis buffer for measurement of superoxide dismutase (SOD) activity and glutathione (GSH) level, and the media were collected for measurement of aspartate aminotransferase (AST) activity. Among each cell lysate, 20 μL volume was used for measurement of SOD activity and the concentration of protein was quantified by Bradford assay for normalization of GSH level. SOD and AST activity as well as GSH level were determined by using a commercial SOD assay kit (Dojindo, USA), Aspartate Transaminase Assay kit (BioAssay Systems, USA), and Glutathione Assay Kit (Sigma), respectively. All experimental procedures were conducted according to the manufacturers' protocols. The absorbance of each well was measured using a microplate reader. The inhibition rate of SOD activity was calculated as per the equation given below:
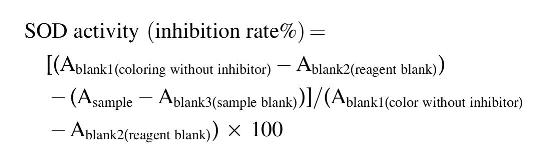
GSH level and AST activity were calculated according to the manufacture's equation.
Measurement of ROS
HepG2 cells were collected in a conical tube and centrifuged to obtain cell pellets (2000 g, 10 min, 4°C). The cells were washed with PBS once and then resuspended in a diluted 2′,7′-dichlorofluorescin diacetate solution at a concentration of 1 × 106 cells/mL for 30 min at 37°C in the dark. The stained cells were washed and seeded at a density of 1 × 105 cells/well in a dark and clear-bottomed 96-well plate. The cells were subsequently treated with 0.0156–1.0 mg/mL of WEAA for 3 h and then exposed to 0.7 mM of t-BHP for another 3 h. The fluorescence of each well was measured at excitation and emission wavelengths of 485 and 535 nm, respectively, using a microplate reader. ROS was detected by using a commercial ROS Detection Kit (Abcam, USA). Relative ROS levels were expressed as fold of control after background subtraction.
2′,2′-Diphenyl-1-picrylhydrazyl-scavenging assay
2′,2′-Diphenyl-1-picrylhydrazyl (DPPH)-scavenging assay was performed as previously described. 30 WEAA were dissolved in 50% ethanol to be final concentration of 0.0078–1.0 mg/mL. Each dissolved WEAA solution was mixed with 0.1 mM DPPH (Sigma) solution, and then kept for 30 min in the dark at room temperature. The optical density was determined at 517 nm.
Real-time quantitative polymerase chain reaction
Total RNA from the HepG2 cells was isolated with NucleoZOL (Macherey-Nage, Germany) according to the manufacture's protocols. cDNA was synthesized using 0.5 μg of total RNA with a ReverTra Ace qPCR RT Master Mix Kit (Toyobo, Japan). Synthesized cDNA was amplified with the target gene primers and THUNDERBIRD SYBR qPCR Mix (Toyobo) by utilizing QuantStudio™ 3 Real-Time PCR System (Applied Biosystems, USA). The amplification cycles were as follows: denaturation at 95°C for 5 min, amplification repeated 40 cycles at 95°C for 5 sec and 60°C for 30 sec, and melting curve analysis at 95°C for 15 sec, 60°C for 1 min, and 95°C for 1 sec. Relative gene expression data were normalized with Gapdh gene expression. Primers are listed in Table 1.
Polymerase Chain Reaction Primers Used In This Study
Animals and experimental groups
All animal procedures were conducted following the protocols approved by the Animal Research Ethics Committee of KPC Co., Ltd. (Approval No. P173009; Gyeonggi-do, Korea). Male C57BL/6 mice (weighing 18–23 g, aged 6 weeks) were purchased from OrientBio Co., Ltd. (Gyeonggi-do, Korea) and maintained in a temperature (21°C ± 2°C)- and humidity (50% ± 20%)-controlled room with a 12-h light and dark cycle. Purchased mice were reallocated into experimental groups to make average weights similar and provided with acclimation period for at least 1 week. The animals had access to food and water ad libitum and were fed with normal diet (Fat 5 kcal%; Purina feed® Solid feed for laboratory animals; Cargill Agri PURINA®, Gyeonggi-do, Korea) or high-fat diet (Fat 45 or 60 kcal%; Research Diet, Inc., USA) for 8 weeks starting when the mice were 7 weeks of age. The treatment of each group was as follows: normal diet and water, high-fat diet and water, high-fat diet and ESM (100 mg/kg), high-fat diet and WEAA (50 mg/kg), and high-fat diet and WEAA (100 mg/kg). Each group was administered treatment materials or water orally once a day for 8 weeks. Body weight and food intake were measured weekly. The order of treatment and assessment was not considered. After food deprivation for 16 h, the mice were anesthetized with 3% isoflurane and blood samples were collected for biochemical analysis. Then, they were euthanized through CO2 inhalation (40%, for 5–10 min) and their livers were obtained for histological investigation.
Histological analysis
Mouse liver tissues were fixed in 10% neutral buffered formalin solution and prepared into paraffin blocks, which were cut into 4-μm sections. The sections were stained with hematoxylin-eosin (H&E) and observed under an optical microscope (Olympus MUV10 microscope, equipped with a DC71 camera; Olympus, Japan). Histological scores were calculated following the criteria presented in Table 2.
Criteria For Histopathological Scores of Hepatocellular Steatoses *
* Matsumoto et al. 31
Biological analysis
Collected blood samples in 0.6 mL SST tube (Microtainer; BD, USA) were maintained at room temperature for 30–60 min to induce blood coagulation. Blood samples were centrifuged at 1500 ×g for 10 min to obtain serum. Serum samples were used to measure the levels of triglyceride, cholesterol, AST, and alanine aminotransferase (ALT) using a Blood Biochemistry Analyzer (Accute TBA-40FR; Toshiba Medical System Co., Japan).
Statistical analyses
Statistical analyses were performed using the GraphPad Prism software (GraphPad Software, USA). For comparison to WEAA-untreated positive control in in vitro experiments, linear regression with a 95% confidence interval and unpaired two-tailed Student's t-test were performed. We examined differences in the results between groups of individual animals using the nonparametric Mann–Whitney test. No samples or animals were excluded from the analyses. Differences with P < .05 were considered statistically significant.
RESULTS
Effects of WEAA on intracellular lipid accumulation in oleic acid-exposed HepG2 cells
HepG2 cells treated with FFAs have been used as models reflecting the key features of steatohepatitis in humans. 32,33 Because treatment with up to 1 mg/mL of WEAA for 24 h did not influence the viability of HepG2 cells, WEAA was used at concentrations lower than 1 mg/mL (data not shown). To assess the effect of WEAA on lipid deposition in hepatocytes, HepG2 cells were pretreated with WEAA for 3 h and then incubated with 1.5 mM of oleic acid, a dietary long-chain FFA abundant in the plasma of both healthy subjects and NAFLD patients. 34 Numerous red lipid droplets fused with each other were observed within the oleic acid only-treated HepG2 cells (Fig. 1A). Compared with those in the positive control group, WEAA pretreatment markedly alleviated lipid accumulation and lipid fusion in HepG2 cells, reaching a maximum inhibition at 0.25 mg/mL (Fig. 1B), suggesting that WEAA might be an effective natural material for prevention of lipid accumulation in the liver.

WEAA alleviates FFA-induced lipid accumulation in hepatocytes. HepG2 cells were pretreated with WEAA (62.5, 125, 250, 500, and 1000 μg/mL). After 3 h, the WEAA was washed out and the cells were cultured in the absence or presence of oleic acid (final concentration: 1.5 mM) for an additional 24 h. Lipid droplets deposited in the cells were stained with Oil red O.
Effects of WEAA on hepatotoxicity and oxidative stress in t-BHP-treated HepG2 cells
In addition to intracellular lipid accumulation in hepatocytes, cellular injury caused by oxidative stress is a primary factor in the development of hepatopathy, such as hepatitis. 35 To assess the hepatoprotective activity of WEAA, HepG2 cells were treated with t-BHP, an organic hydroperoxidant that causes cell damage induced by oxidative stress. 36 To determine the appropriate concentration of t-BHP for our experiments, cells were treated with various concentrations of t-BHP for 3 h. t-BHP reduced cell viability in a dose-dependent manner, displaying an LC50 value of 0.7836 mM (data not shown). Therefore, 0.7 mM t-BHP was used in the subsequent experiments. As expected, exposure to 0.7 mM of t-BHP lowered the viability of HepG2 cells by ∼55.6%, whereas WEAA pretreatment protected cells from the cytotoxicity of t-BHP in a dose-dependent manner (Fig. 2A). The t-BHP-induced cytotoxicity was completely restored by WEAA at concentrations higher than 0.25 mg/mL, and the effect reached a plateau at 0.5 mg/mL (Fig. 2A). AST, an enzyme associated with liver parenchymal cells, is released in response to hepatocellular injury; therefore, AST level has been used as a biochemical marker in the diagnosis of hepatic diseases. 37 Parallel with the decrease in cellular viability, oxidative stress caused by t-BHP resulted in AST leakage from HepG2 cells to the media, with a value of 23.4 U/L (Fig. 2B). Consistent with the results of cellular viability, WEAA pretreatment decreased the leakage of AST in a dose-dependent manner (Fig. 2B). These results indicated that WEAA exhibited a hepatoprotective effect against oxidative stress-induced cytotoxicity.

WEAA has a cytoprotective effect against oxidative damage in hepatocytes. HepG2 cells were pretreated in the presence or absence of WEAA (15.6, 31.3, 62.5, 125, 250, 500, and 1000 μg/mL) for 3 h, and then exposed to t-BHP (final concentration: 0.7 mM) for an additional 3 h.
Next, we investigated the antioxidant activities of WEAA in t-BHP-treated HepG2 cells. As t-BHP is metabolized into free radical intermediates in cells, t-BHP treatment has been known to induce ROS production and to reduce several antioxidant indices, such as GSH level and SOD activity. As shown in Figure 3A, t-BHP treatment increased ROS production 8.3-fold. In contrast, WEAA pretreatment dramatically decreased the level, indicating a potent antioxidant activity (Fig. 3A). Treatment with t-BHP reduced the level of GSH and the activity of SOD by 18.4 μM and 11.8%, respectively, whereas untreated HepG2 cells showed a GSH level of 42.78 μM and SOD activity of 38.92% (Fig. 3B, C). WEAA restored the cellular level of GSH in a dose-dependent manner; moreover, at concentrations higher than 0.25 mg/mL, it enhanced GSH level to a higher amount of that in nontreated cells (Fig. 3B). Furthermore, pretreatment with WEAA clearly increased the enzyme activity of SOD in a dose-dependent manner (Fig. 3C). The restoration was dramatic at WEAA concentrations up to 0.0625 mg/mL, but gentle at higher concentrations, although the restoration did not reach the level in naive cells (Fig. 3C). In addition, we measured the mRNA expression of glutamate cysteine ligase (GCL), which is the first and rate-limiting enzyme of the cellular GSH synthesis. 38 The Gcl mRNA expressions were significantly increased by WEAA treatment at 0.5 and 1 mg/mL (Supplementary Fig. S1). The result could explain, in part, the increase of GSH content by WEAA treatment. To test whether WEAA directly acts as an antioxidant, the extract was reacted with DPPH solution composed of free radical molecules. WEAA showed strong free radical-scavenging activity by itself, reaching a plateau at a concentration of 0.25 mg/mL (Fig. 3D), which implied that it could alleviate ROS generation and relieve oxidative stress-induced cellular injury. These results suggested that WEAA may protect hepatocytes from oxidative stress by activating cellular antioxidant enzymes and by acting as a free radical scavenger.
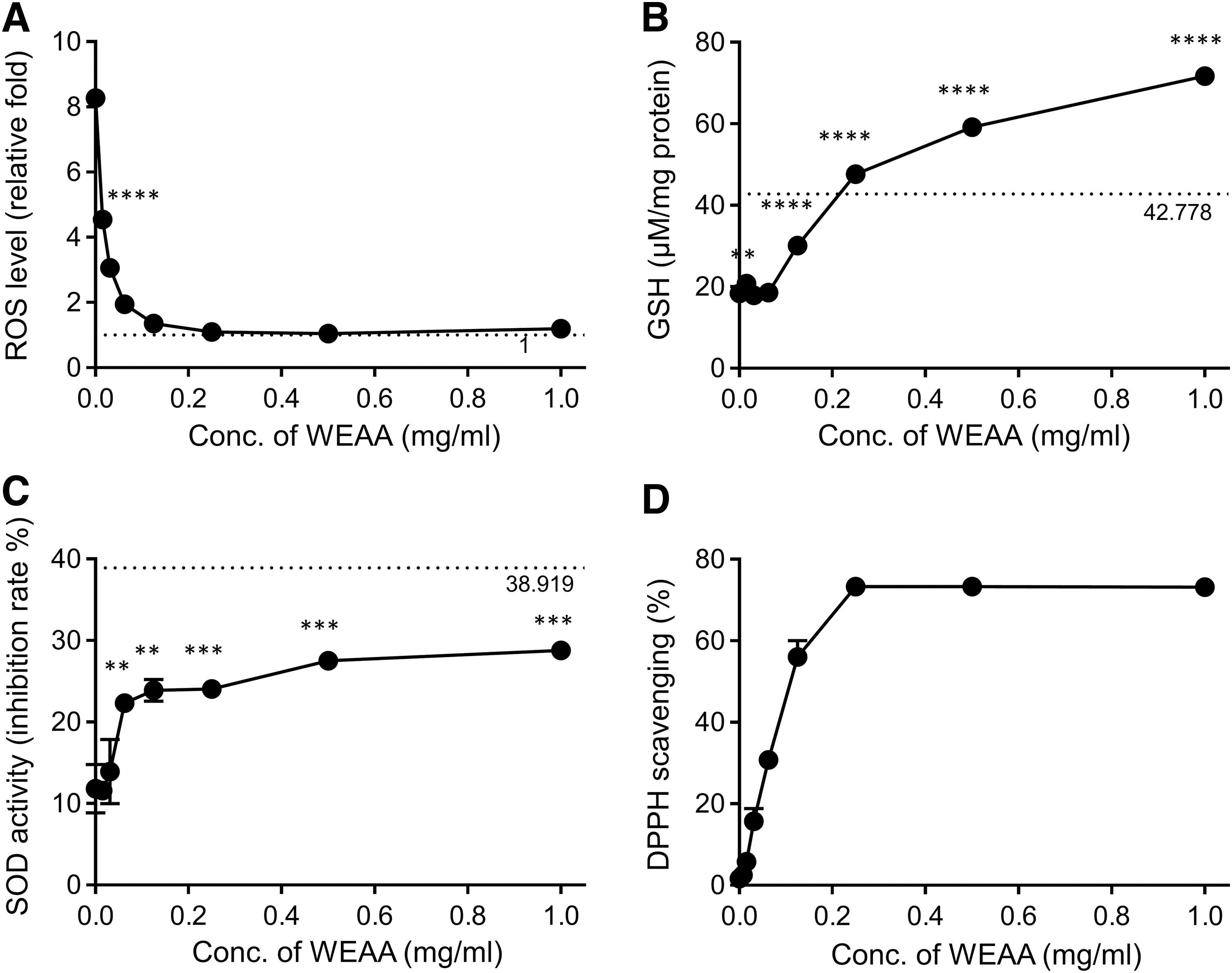
WEAA exhibits antioxidant activities.
Effects of WEAA on body weight in high-fat diet-induced fatty liver mouse models
Generally, high-fat diet-fed animals have been used to study manifestations of NAFLD. 39 C57BL/6 mice treated with 60% high-fat diet and water for 55 days had markedly increased body weight by ∼1.5-fold (at day 55), compared with that of the normal chow-fed group (Fig. 4). When mice were intragastrically administrated with WEAA (50 or 100 mg/kg), the weight gain was significantly suppressed by 19.1% and 10.4% (at day 55), respectively (Fig. 4). About 14% reduction in body weight was also observed in ESM-fed group used as a comparative positive control. Moreover, to assess the effect of WEAA in mild weight gain, we established fatty liver mice induced by the administration of 45% high-fat diet, instead of 60% high-fat diet. The intake of 45% high-fat diet increased the body weight 1.17-fold at day 49 in comparison with the normal chow diet group (Supplementary Fig. S2A). The weight gain was dramatically suppressed by WEAA (100 mg/kg) administration, almost similar to the level observed for the normal chow-intake group (Supplementary Fig. S2A). These data indicated that WEAA has the potential to alleviate weight gain caused by high-fat diet intake.

WEAA reduces weight gain in high (60%)-fat diet-induced fatty liver mouse models. Except for those in the normal diet group (n = 5), mice were provided 60% high-fat chow (n = 10 per group) during the experimental period. From day 0, all mice were intragastrically administrated an ESM (100 mg/kg), WEAA (50 or 100 mg/kg), or water once every day. The treatment of each group is as follows: normal diet, black triangle; high-fat diet and water, white triangle; high-fat diet and ESM, white square; high-fat diet and WEAA (50 mg/kg), black circle; high-fat diet and WEAA (100 mg/kg), white circle. The data represent mean ± SEM. Mouse body weight in each experimental group was compared to that in the high-fat diet control water group using Mann–Whitney test, with pairs defined by time point. The data shown are representative of two independent experiments. *P < .05, **P < .01, ***P < .001, and ****P < .0001. ESM, extract of Silybum marianum; SEM, standard error of the mean.
Effects of WEAA on lipid accumulation in the liver tissue of high-fat diet-induced fatty liver mouse models
To evaluate structural changes in fatty liver, liver sections were stained with H&E. Severe structural degenerations, such as marked fatty deposition and inflammatory cell infiltration, were observed in liver specimens from control mice consuming high-fat diet (Fig. 5), indicating that fatty liver was established successfully. WEAA administrations improved steatohepatitis and cell infiltration within and between hepatocytes (Fig. 5). In both WEAA-treated groups (50 and 100 mg/kg), the reductions of fat accumulation and cell infiltration were more dramatic than those in ESM-fed animals, although high-dose treatment with WEAA (100 mg/kg) showed lower efficacy in reducing body weight than ESM and low-dose WEAA (50 mg/kg), as shown in Figure 4. In fact, there were no differences in effect on steatohepatitis between 50 and 100 mg/kg WEAA. In addition, oral administration of WEAA tended to inhibit steatosis in mild fatty liver induced by 45% high-fat diet, although the difference was not statistically significant (Supplementary Fig. S2B). These data showed that WEAA exhibited more effective inhibitory activity on lipid accumulation and immune cell infiltration than that of ESM.
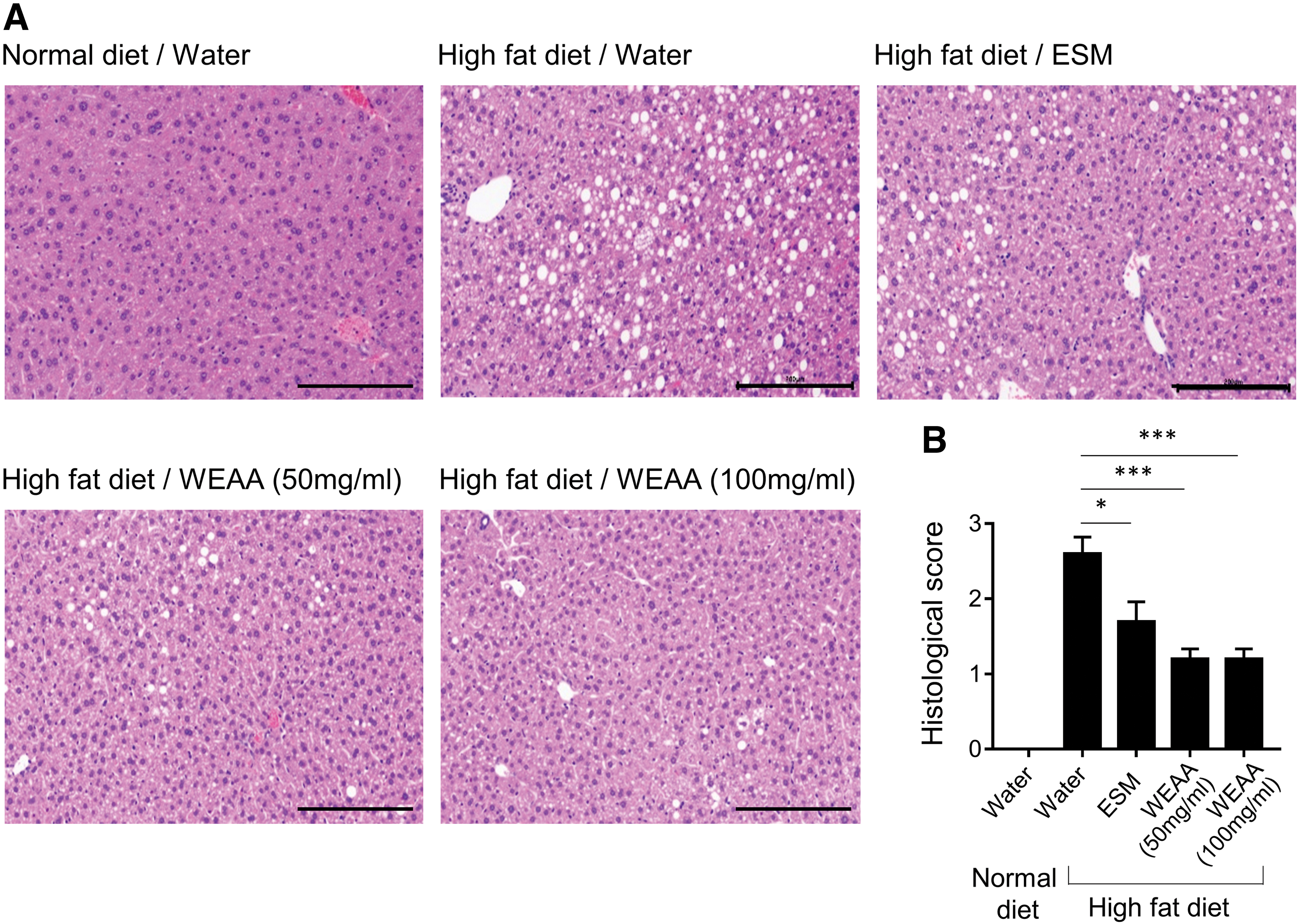
WEAA ameliorates lipid deposition in high-fat diet-stimulated liver tissues.
Effects of WEAA on biochemical parameters in the peripheral blood of high-fat diet-induced fatty liver mouse models
To investigate biochemical parameters in plasma, all mice were starved 1 day before sacrifice, and then their plasma was collected on day 56 and analyzed. High-fat diet intake tended to slightly increase the level of plasma triglycerides, despite no statistical difference, and significantly increase the amount of total cholesterol (Fig. 6A, B). Compared to those in control mice that consumed high-fat diet, plasma triglyceride concentrations were significantly decreased in the WEAA-fed groups (Fig. 6A); however, the increased level of total cholesterols was not altered by WEAA administration (Fig. 6B). In contrast, ESM administration did not have any effect on both plasma triglyceride and total cholesterol levels (Fig. 6A, B).
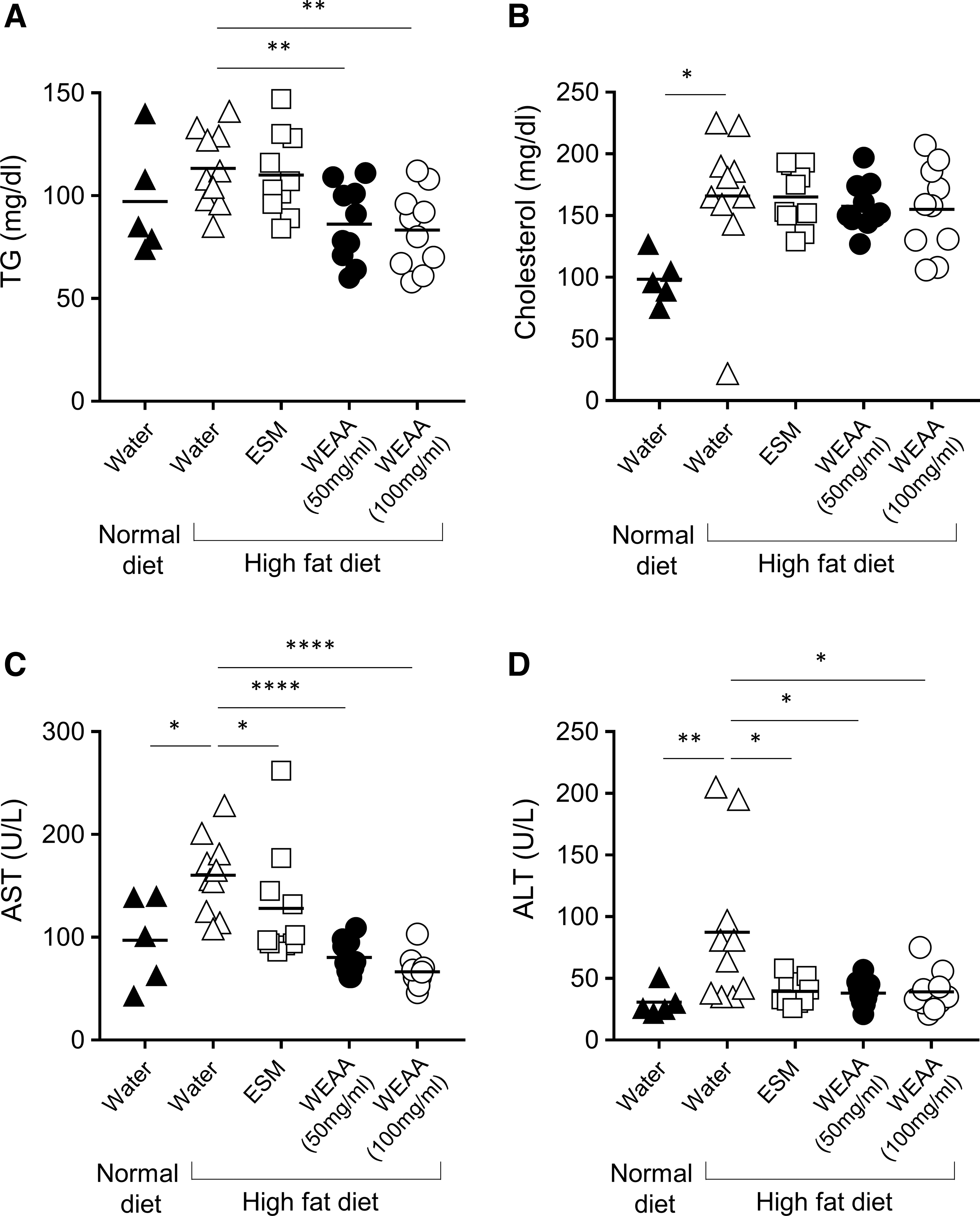
WEAA reduces the levels of plasma TG, AST, and ALT, but not that of cholesterol, in high-fat diet-induced fatty liver mouse model. After the last treatment (day 55), all mice were fasted for 1 day, and then samples were obtained on day 56. The levels of TG, cholesterol, AST, and ALT in the collected plasma were measured by ELISA. Each dot indicates an individual mouse. Horizontal lines show the mean. The data shown are representative of two independent experiments. *P < .05, **P < .01, ****P < .0001; compared to the levels of high-fat diet control group, as analyzed by Mann–Whitney test. ALT, alanine aminotransferase; ELISA, enzyme-linked immunosorbent assay; TG, triglyceride.
Exposure to high-fat diet also increased plasma AST and ALT levels, compared with those in mice fed normal chow diet (Fig. 6C, D and Supplementary Fig. S2C). Similar to its effect on triglyceride level, treatment with WEAA at both concentrations evidently decreased the generation of AST and ALT, restoring the levels almost to those shown in the normal chow diet group (Fig. 6C, D and Supplementary Fig. S2C). These results were consistent with the in vitro data and liver histopathology data (Figs. 2B and 5). Moreover, the inhibitory activity of WEAA on triglyceride, AST, and ALT levels were greater than that of ESM (Fig. 6A, C, D). These results suggested that WEAA ameliorated plasma biochemical parameters that are important factors in NAFLD pathogenesis.
DISCUSSION
In this study, we investigated the hepatoprotective activity of a water extract of A. annua to evaluate the therapeutic/prophylactic potential of the extract for the treatment of NAFLD. For this purpose, oleic acid-treated or t-BHP-treated HepG2 cells and high-fat diet-fed mice were used as in vitro and in vivo experimental systems, respectively. In addition to the general benefits of using a cell line, such as easy maintenance, HepG2 cells possess the typical metabolic and enzymatic activities of primary hepatocytes, thereby reflecting the morphological and biochemical characteristics of FFA-induced lipid accumulation and oxidative stress-induced cytotoxicity. Thus, the cell line has been widely used as a typical in vitro model of hepatocytes to investigate the effects of candidate substances in various metabolic and toxicity studies. Administration of high-fat diet in vivo causes hepatic steatosis and fatty liver changes as well as metabolic abnormalities, such as obesity and insulin resistance, even though hepatic inflammation and fibrosis, the more aggressive features of fatty liver diseases, seldom occur in high-fat diet only-fed animals. Thus, in this study, high-fat diet-fed mice were used as an in vivo model for investigation of the efficacy of a natural material against the initial pathogenesis of NAFLD.
Lipid accumulation in hepatocytes is a representative feature of NAFLD. Although the elevated flux of FFAs into the liver is currently regarded as benign, evidence has indicated that hepatic lipid accumulation generally precedes progression to nonalcoholic steatohepatitis, cirrhosis, and hepatocellular carcinoma. In other words, hepatic deposition of lipid droplets precedes toxic substance production, lipotoxicity, and liver damage, which lead to progression to more aggressive fatty liver diseases. 6,40 In this study, WEAA treatment ameliorated lipid accumulation in HepG2 cells and in the liver of high-fat diet-fed mice, suggesting that WEAA may be potential as a therapeutic/prophylactic substance for the treatment of the initial pathogenesis of various fatty liver diseases, including NAFLD.
Hepatocytes contain aminotransferases, such as AST and ALT, in their cytoplasm. Excessive oxidative stress leads to damage of cell integrity and leakage of cellular molecules, such as proteins and DNA, including liver-specific enzymes. Thus, increased levels of aminotransferases in a culture medium or in blood are considered as markers of hepatotoxicity and hepatic damage. 41 In this study, t-BHP, which was metabolized into free radical intermediates within cells, was used to facilitate lipid peroxidation, triggering the generation of oxidative stress and finally the damage of hepatocytes. Our data showed that WEAA could inhibit t-BHP-mediated AST release and cytotoxicity in HepG2 cells and alleviate the plasma levels of AST and ALT increased by high-fat diet intake, indicating that WEAA exhibited a hepatoprotective effect against oxidative stress-mediated cell damage.
To understand the mechanism underlying the mitigating effect of WEAA against hepatotoxicity, we investigated the effect of WEAA as an enzymatic defense mechanism and its own activity as an antioxidant. In this study, we measured SOD activity and GSH level as a typical antioxidant enzyme and an antioxidant enzymatic product, respectively. SOD plays a key antioxidant role of scavenging the radical intermediates generated by incomplete oxidation. GSH plays a pivotal role in protecting cellular molecules from endogenous and exogenous oxidants, such as superoxide anion, hydroxyl radical, nitric oxide, and carbon radicals, and it is thus involved in the process of detoxification. 38 WEAA ameliorated the SOD activity and GSH level suppressed by an organic peroxide, suggesting that WEAA could scavenge the excessive free radicals produced by leakage of incompletely reduced oxygen generated through electron transport chain in the mitochondria. Consistently, the increased ROS level was also dramatically normalized by WEAA pretreatment. Moreover, WEAA had the ability to directly quench stable or free radical molecules. Taken together, the hepatoprotective efficacy of WEAA was attributable to its ability to increase antioxidant enzyme activity and to its own free radical scavenging activity.
Although artemisinin is the most famous component in A. annua, it was not detected by HPLC analysis in the WEAA in this study, which was composed of A. annua grown in Korea. Instead, the following six major compounds were determined: five dicaffeoylquinic acid derivatives (1,3-di-O-caffeoylquinic acid, 3,4-di-O-caffeoylquinic acid, 3,5-di-caffeoylquinic acid, 1,5-di-O-caffeoylquinic acid, and 4,5-di-O-caffeoylquinic acid) and one coumarin derivative (scopoletin).
42
Another research group identified five compounds from A. annua grown in Cameroon, namely artemisinin, scopoletin, chrysosplenetin, eupatin, and 3-O-β-
In our current study, we used an extract prepared from S. marianum as a positive control. For centuries, milk thistle, the popular name of S. marianum, has been used for the treatment of liver diseases. 55 Moreover, many studies have reported that the main active component and by-product of S. marianum extract possess strong antioxidant and anti-inflammatory activities, thereby improving hepatic steatosis and oxidative stress in various in vitro and in vivo systems. 56 Our in vivo results showed that WEAA had higher efficacy than ESM at the same concentration in terms of improvement of body weight, hepatic lipid accumulation, and triglyceride, AST, and ALT plasma levels. Moreover, the results of macroscopic observation (data not shown) and measurement of plasma parameters (Fig. 6) showed no particular side effect of WEAA, although its exact toxicity should still be elucidated. Thus, our results indicated that WEAA may be an attractive natural product for the treatment and prevention of liver diseases.
Taken together, WEAA activated antioxidant activities, which in turn regulated the proteins involved in fatty acid and cholesterol synthesis, resulting in hepatoprotective effect against oxidative damage. Moreover, WEAA treatment directly ameliorated lipid accumulation and whole-body weight gain in mice. Our findings suggested that WEAA could be a candidate treatment with high efficacy to improve hepatic lipid accumulation and liver damage. Furthermore, its compounds or fractions could be used as sources for more effective drug development for the treatment of NAFLD.
Footnotes
ACKNOWLEDGMENTS
We thank Ki-Moon Park in SungKyunKwan University and Hee-Jin Yang in SungKyun BioTech. Co., Ltd. for their advice and support to the project.
AUTHOR DISCLOSURE STATEMENT
No competing financial interests exist.
FUNDING INFORMATION
This work was supported by Korea Institute of Planning and Evaluation for Technology in Food, Agriculture, Forestry and Fisheries (IPET) through High Value-added Food Technology Development Program, funded by Ministry of Agriculture, Food and Rural Affairs (MAFRA) number 2017100491. The funding body has no role in the design of the study and collection, analysis, and interpretation of data and in writing the article.
SUPPLEMENTARY MATERIAL
Supplementary Figure S1
Supplementary Figure S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.