
Abstract
3,3′-diindolylmethane (DIM) has an anticancer activity, but the role DIM plays on malignant melanoma cells and its specific mechanism is unclear. We studied the biological effects of DIM on malignant melanoma cells and the related mechanism and the results showed that DIM significantly suppressed cell proliferation and induced apoptosis in malignant melanoma cells. In addition, the expression levels of phosphatase and tensin homolog deleted on chromosome ten (PTEN), Bax, Bid, cleaved caspase-3, and cleaved caspase-9 were increased after DIM treatment. In A2058 PTENmut cells, DIM-mediated inhibition of proliferation and DIM-induced apoptosis were attenuated. Additionally, the overexpression and knockdown of PTEN could regulate such effects of DIM in malignant melanoma cells. Furthermore, DIM exerted growth-inhibiting and apoptosis-inducing effects in vivo. This study demonstrated that DIM has antitumor effect in human malignant melanoma cells through the mitochondrial apoptotic pathway activated by PTEN/Akt signaling.
Introduction
Malignant melanomas develop through malignant transformation of melanocytes, which are cells that produce melanin pigment. 1 Although the majority of cases occur cutaneously, malignant melanomas can be found as primary melanomas in other organs and tissues. However, melanoma localized to the skin, is the most aggressive and fatal form of melanoma. 2 As is the case with other solid tumors, staging is an important determinant of survival. The 5-year survival rates of early stage localized melanoma can reach 90%, 3,4 whereas those of invasive and metastasized melanomas still have a poor prognosis despite the marked advances of targeted therapies and immunotherapies. 5,6 Resistance of invasive and metastasized melanoma to conventional chemotherapy treatments is common, limiting the curative effects of these treatments. Therefore, for the treatment of malignant melanomas, natural anticancer compounds with high efficacy and low toxicity are urgently needed.
While indole-3-carbinol (I3C) has been proven to exert anticancer effects, 7 3,3′-diindolylmethane (DIM) a derivative of I3C, exerts antitumor activity in a variety of tumors 8 –13 by regulating several biological processes. Additionally, it has been reported that DIM can activate insulin signaling and promote glucose uptake. 14 Furthermore, in experimental colitis, DIM exerted anti-inflammatory effects by significantly suppressing the expression of several cancer-associated cytokines, such as vascular endothelial growth factor. 15 Meanwhile, DIM shows biological effects through modulating multiple gene signaling pathways, such as Wnt and IL6/STAT3. 16 –18
This study aimed to examine the anticancer effects of DIM in malignant melanoma cells and the related mechanisms.
Materials and Methods
Cells and reagents
Human malignant melanoma cell lines (A375, A2058, Mel501, and SKMel28) were purchased from Shanghai Cell Collection. These cells were maintained in Dulbecco's Modified Eagle Medium (DMEM), which is supplemented with fetal bovine serum (10%), streptomycin (100 μg/mL), and penicillin (100 U/mL) at 37°C in a CO2 atmosphere. DIM was obtained from Sigma-Aldrich (St. Louis, MO, USA).
Plasmid
Human phosphatase and tensin homolog deleted on chromosome ten (PTEN) was cloned from HEK 293T cells using reverse transcription/PCR. The expression of plasmid-efficient short interfering RNA (siRNA) sequences targeting PTEN (NM_000314.7) synthesized by Shanghai GeneChem Co. Ltd. (China) were constructed into the modified pcDNA3-Flag-Dest-B vector for transfection. After transfection of plasmids into HEK 293T cells and puromycin selection for 4 days, the survival clones were cultured in the DMEM with puromycin (1 μg/mL). Western blot was used to confirm gene disruption.
Proliferation assay
A total of 5 × 103 cells per well were seeded, then fresh medium supplemented with DIM (0, 50, 100, 200 μM) at different times were added after replacing the supernatant. Then, cells were incubated with Cell Counting Kit-8 (CCK-8) solution (Beyotime, Shanghai, China) for another 2 h. The microplate reader (Victor3 1420 Multilable Counter; PerkinElmer, USA) was used to determine the absorbance at 450 nm.
The appropriate number of cells (1 × 103 cells/well) were seeded, and next day cells were cultured with DIM (0, 50, 100, and 200 μM) after the supernatant was replaced. The supernatant was refreshed every 3 days. After 5 days, these cells were fixed and then stained. Then the counts of established cell colonies (>100) were scored.
Hoechst 33258 staining
Cells were cultured in the medium supplemented with or without DIM for 24 h. After being fixed and washed, Hoechst 33258 was used to stain the cells. A fluorescent microscope (BX51; Olympus, Tokyo, Japan) was used to count the apoptotic cells.
Annexin V-phycoerythrin/7-aminoactinomycin D staining
Seeded cells were washed and centrifuged. Then, according to the manufacturer's recommendations, cells were stained with the annexin V-phycoerythrin/7-aminoactinomycin D (Annexin V-PE/7-AAD) (Beyotime) and analyzed immediately by flow cytometry (BD FACS Aria IIIⅢ, USA). For each experiment, we repeated the total procedure at least three times.
Immunofluorescence assay
A375 and A2058 cells (1 × 105 cells) on 20 mm sterile circular microscope coverslips were treated with DIM (100 μM) for 24 h. After being washed and then being fixed (for 15 min), permeated with Triton X-100 (0.4%, 10 min) and then blocked with bovine serum albumin (5%, 30 min), the cells were incubated with anti-PTEN antibody (overnight, 4°C) and secondary antibody (1 h, room temperature). 4,6-diamino-2-phenyl indole (2.5 μg/mL) was used to stain cell nuclei. For each experiment, we repeated the total procedure at least three times.
Mitochondrial membrane potential
The JC-1 Kit (Beyotime) was used to check the mitochondrial membrane potential (△ψm). After washing, the seeded cells were maintained (37°C, 20 min) in JC-1 solution. Next, cold buffer solution was used to wash cells before they were examined. Each experiment was repeated at least three times.
Western blot analysis
RIPA buffer was used to lyse cells and the Cell Mitochondria Isolation Kit (Beyotime) was used to separate the mitochondrial proteins. After the protein concentration was determined, the denatured protein mixture was subjected to sodium dodecylsulphate polyacrylamide gel electrophoresis, then transferred to nitrocellulose membranes, and blocked with tris buffered saline with tween 20 buffer (containing 5% nonfat milk). Then, the membranes were incubated with the primary antibodies (overnight, 4°C) and secondary antibody (1 h, room temperature). Then West-Q Pico Dura ECL Solution (Gendeport, Barker, TX, USA) was used to detect the antigen–antibody complexes. For each experiment, we repeated the total procedure at least three times.
Xenograft tumor model
BALB/c nude mice (5 weeks old, male) were purchased from Animal Center of Wuhan University. Malignant melanoma cells (A375 PTENWT) were subcutaneously inoculated. These mice were randomly divided into empty vector with nontargeting siRNA group, empty vector with nontargeting siRNA with DIM group, PTEN-siRNA group, and PTEN-siRNA with DIM group. They were intraperitoneally injected with DIM three times per week and weighed as well as monitored the size of each tumor every 3 days. Finally, tumors were separated and weighed. Then Hematoxylin and Eosin (H&E) stain and terminal deoxynucleotidyl transferase-mediated dUTP-biotin nick end labeling (TUNEL) assays were used for analyses. All mouse experiments were approved by the Animal Care and Use Committee of Wuhan University (2019-CK01), and carried out according to the University Guidelines for the Care and Use of Laboratory Animals.
H&E staining and TUNEL assay
After being fixed, dehydrated and embedded, the tumor sections were stained with H&E. The TUNEL kit was used to detect apoptosis. Positive cells were identified, counted, and analyzed by light microscopy in six random fields per slides.
Statistical analyses
Data are shown as the mean ± standard deviation. Differences between groups were compared using a two-way analysis of variance followed by a post hoc Tukey's test. Differences among three or more groups were analyzed by one-way analysis of variance with Bonferroni's correction. We chose SPSS version 17.0 for all statistical analyses.
Results
DIM had proliferation-inhibitory and apoptosis-inducing effects in malignant melanoma cells
The CCK-8 results indicated that DIM suppressed the proliferation of A375, Mel501, and SKMel28 cells, but not that of A2058 cells (Fig. 1A). The colony number in DIM-treated groups was decreased in A375, Mel501, and SKMel28 cells but not A2058 cells (Fig. 1B). The above findings suggest that DIM has an inhibitory effect on A375 cell proliferation.
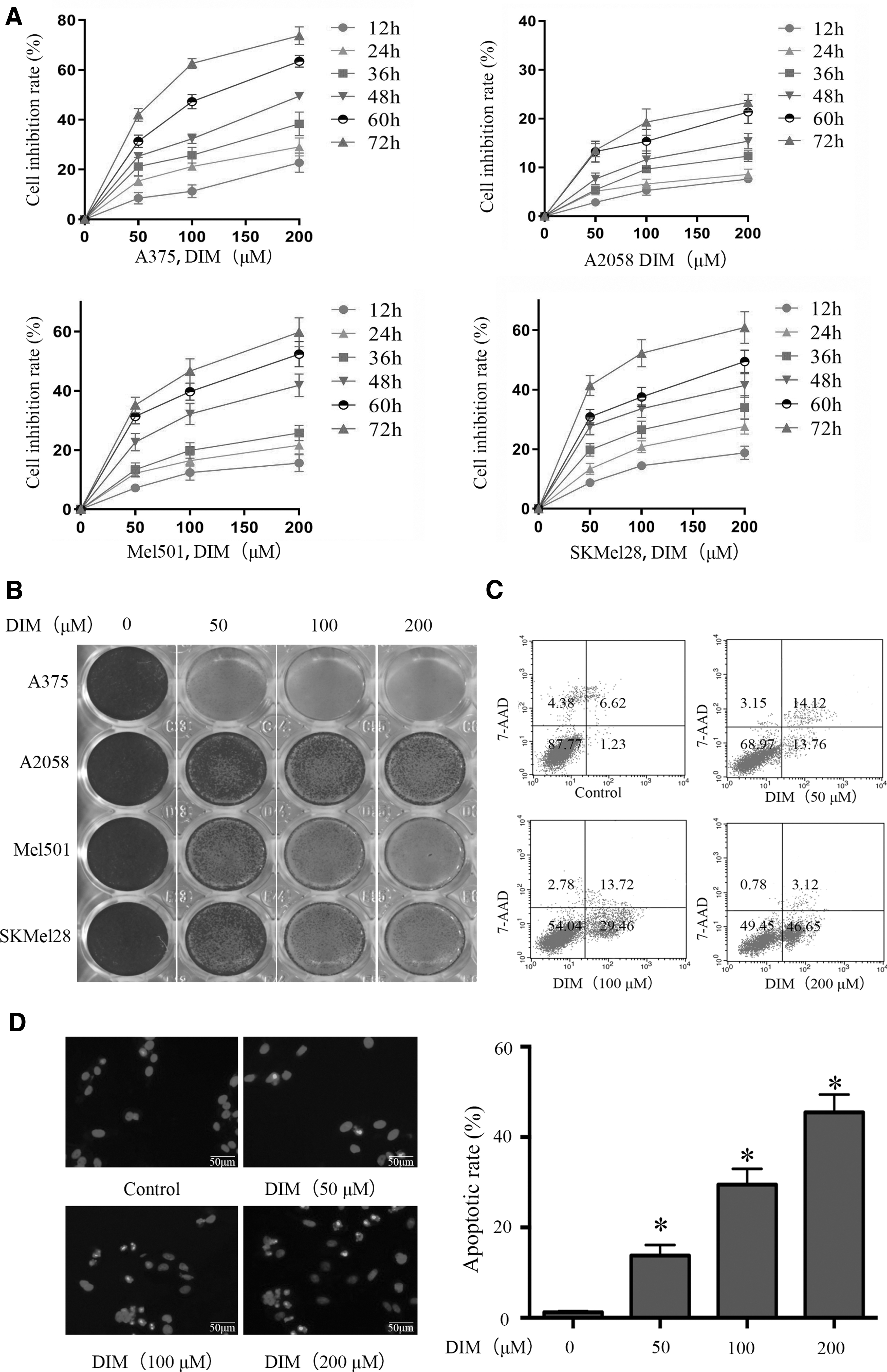
DIM inhibited the proliferation and induced the apoptosis of malignant melanoma cells.
The Annexin V-PE/7-AAD staining indicated that DIM induced the apoptosis of A375, Mel501, and SKMel28 cells but not that of A2058 cells (Fig. 1C). The apoptotic rates in the DIM groups were increased in A375, Mel501, and SKMel28 cells but not in A2058 cells (P < .05, Fig. 1D). The western blot results, shown in Figure 2, revealed that the expressions levels of the critical proteins of mitochondria-mediated apoptosis were increased, which indicated that the mitochondrial pathway may contribute to the DIM-induced apoptosis. Because A2058 cells are PTENmut cells, whereas A375, Mel501, and SKMel28 are PTENWT cells, we speculated that PTEN signaling may impact the antitumor effects of DIM in malignant melanoma cells through the mitochondrial apoptotic pathway.

DIM induced apoptosis of malignant melanoma cells through the mitochondrial pathway.
PTEN/Akt signaling may contribute to the antitumor effects of DIM in malignant melanoma cells
Given the major interest in identifying how DIM exerts proliferation-inhibitory and apoptosis-induced effects, we studied the expression of proteins in cancer-related pathways. As shown in Figure 3A and C, we found that DIM could increase both the protein and messenger RNA levels of PTEN and subsequently inhibit the phosphorylation of Akt (at Thr308), as well as the downstream signaling which may contribute to DIM-mediated proliferation-inhibitory and apoptosis-inducing effects in A375, Mel501, and SKMel28 cells but not in A2058 cells. In addition, the expression of PTEN and phosphorylated PTEN was highest in A375 cells, whereas no PTEN expression was observed in A2058 cells (Fig. 3B), which is consistent with previous reports. 19,20 Furthermore, immunofluorescence assay revealed that the fluorescence intensity of PTEN was markedly higher in the DIM group (Fig. 3D), which suggested that DIM influenced PTEN expression.
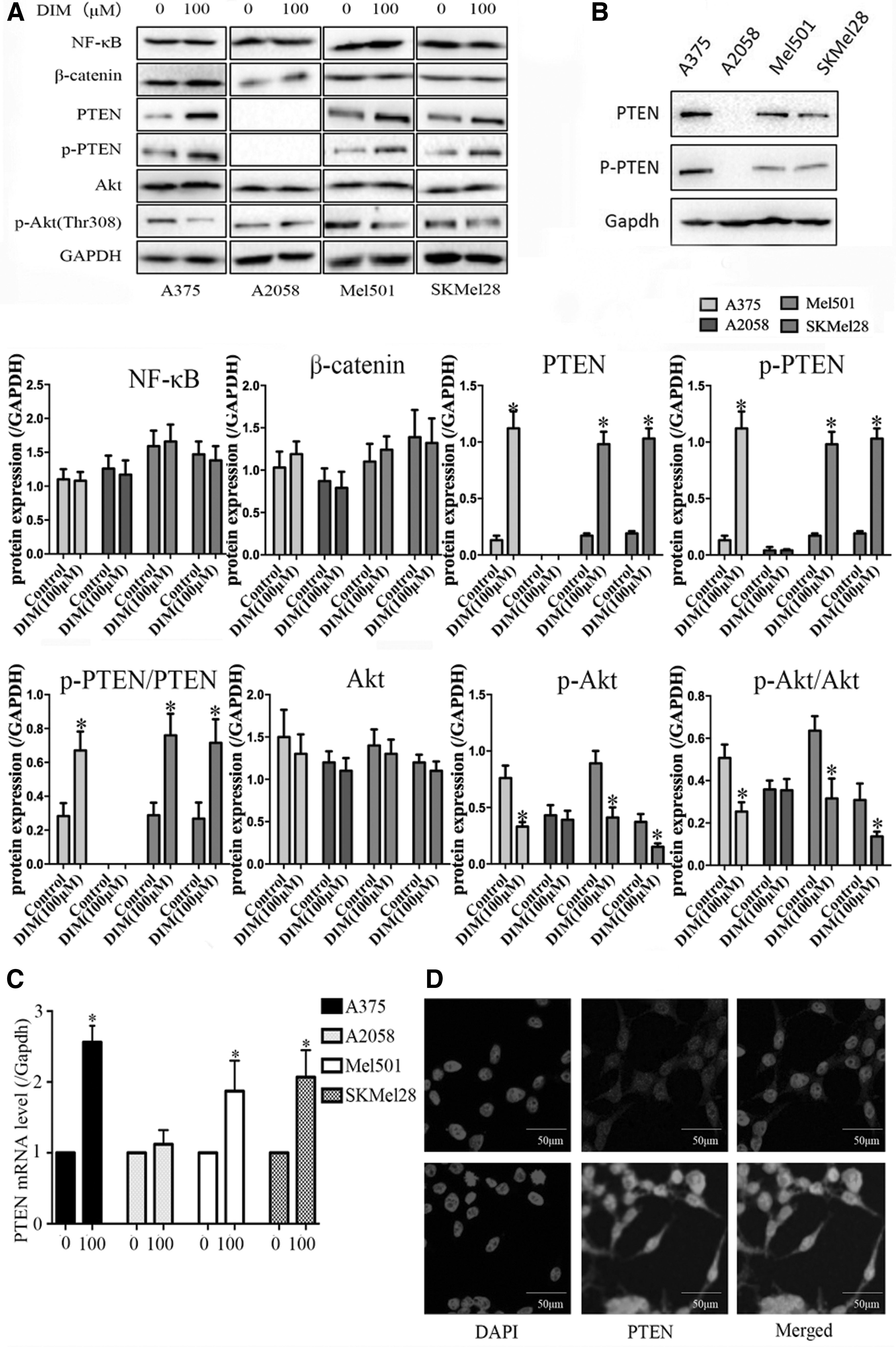
DIM increased the expression of PTEN and the subsequent inactivation of Akt signaling.
These findings led us to propose that PTEN/Akt signaling contributes to the antitumor effects of DIM in human malignant melanoma cells.
PTEN regulated the antitumor effects of DIM in malignant melanoma cells
To further explore the roles of PTEN and the inactivation of Akt signaling in DIM-mediated proliferation-inhibitory and apoptosis-induced effects, PTEN-siRNA and Flag-PTEN plasmids were constructed to interfere with the expression of PTEN and phosphorylated Akt. Western blot results showed that the PTEN-siRNA and Flag-PTEN plasmids influenced the expression of PTEN (P < .05; Fig. 4A, B). In addition, the CCK-8 results suggested that knockdown of PTEN in A375 PTENWT cells could significantly attenuate the inhibitory effects of DIM on cell proliferation (P < .05, Fig. 4C, D). The colony number in PTEN-siRNA-treated cells in the DIM group was much greater than that in nontargeting siRNA-treated cells in the DIM group (Fig. 4E, F). Furthermore, the apoptotic rate was markedly higher in PTEN-siRNA-treatment A375 cells in the DIM group than that in nontargeting siRNA-treatment A375 cells in the DIM group (Fig. 4G–I).

The overexpression and knockdown of PTEN regulated the effects of DIM on the proliferation and apoptosis in malignant melanoma cells.
PTEN/Akt signaling-mediated activation of mitochondrial pathway contributed to DIM-induced apoptosis in malignant melanoma cells
DIM could increase the △ψm in A375 PTENWT cells, and the knockdown of PTEN could suppress the DIM-induced increase in △ψm (Fig. 5A). However, there was no obvious difference in A2058 PTENmut cells when treated with DIM (Fig. 5A), but the △ψm was increased with overexpressed PTEN using Flag-PTEN.
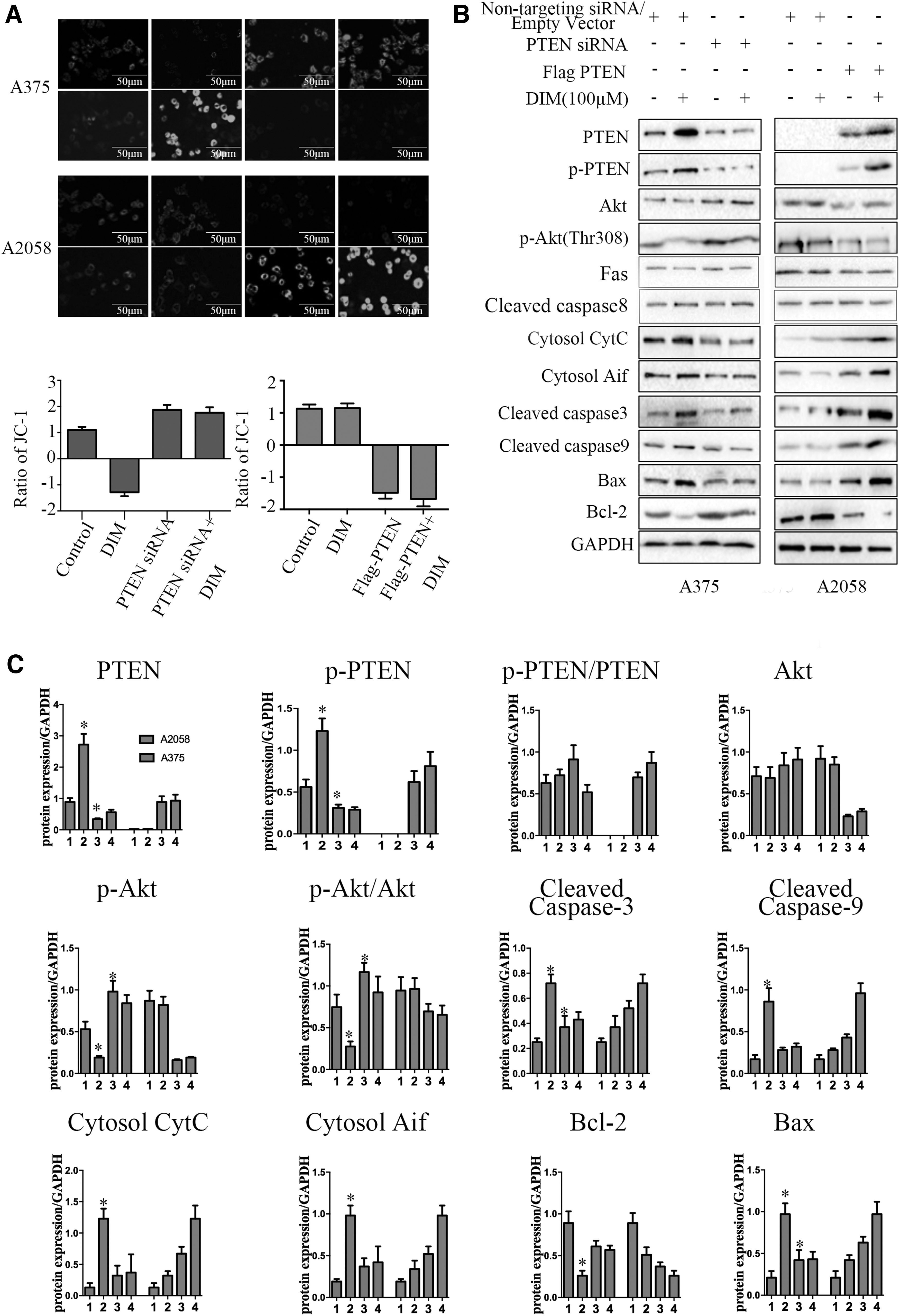
△ψm and Western blot showed the overexpression or knockdown of PTEN regulated the effects of DIM on the apoptosis in A375 PTENWT or A2058 PTENmut cells.
As shown in Figure 5B, in A375 PTENWT cells, DIM increased the expressions of cytosol AIF and Cyt C, cleaved caspase-3 and -9, Bax, and Bid, and decreased the expression of Bcl-2 as well as the antiapoptotic/proapoptotic protein ratio, then subsequently induced mitochondria-mediated apoptosis. However, the knockdown of PTEN could attenuate DIM-mediated changes in the expression of mitochondrial pathway proteins. On the contrary, in A2058 PTENmut cells, DIM could not cause any obvious change in the expression levels of mitochondrial pathway proteins unless the overexpression of PTEN using Flag-PTEN.
Collectively, these results support that DIM exerted proliferation-inhibitory and apoptosis-inducing effects in human malignant melanoma cells through mitochondrial pathway activated by PTEN/Akt signaling.
Effects of DIM on xenograft tumor growth
Next, we studied whether DIM could exert antitumor effects in vivo. As shown in Figure 6, DIM inhibited tumor growth (P < .05). The weight of tumors in PTEN-siRNA-treated A375 cells in the DIM group was greater than that in nontargeting siRNA-treated A375 cells in the DIM group (P < .05). The results from H&E staining and TUNEL assay showed that DIM induced obvious cell apoptosis in the tumor mass (Fig. 6B) (P < .05).
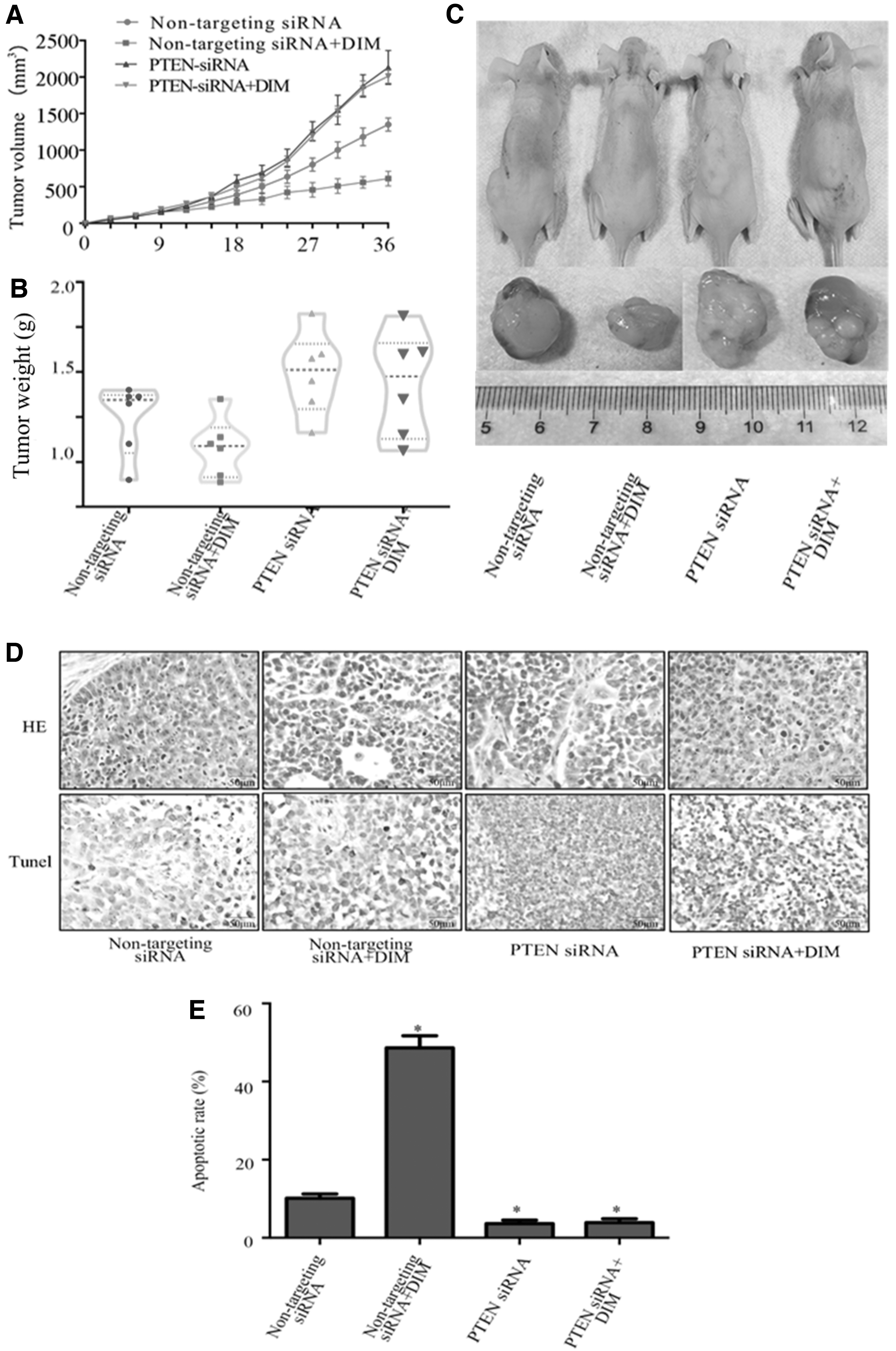
In vivo antitumor effect of treatment groups in a malignant melanoma xenograft mouse model.
Discussion
It has been reported that the mutation rate of PTEN is comparatively high both in melanoma cell lines and in primary melanomas. While the expression of PTEN is ectopic, tumorigenicity and metastasis could be reduced in PTEN-deficient melanoma cells. PTEN dephosphorylates phosphatidylinositol-3,4,5-trisphosphate and then inactivate Akt pathway, 21 which is involved in various cell processes. 22 –24 Collective studies demonstrated that DIM exerted antitumor activity in several types of cancers through PTEN/Akt signaling. 25 DIM enhanced the toxic effects of LY294002 on colon cancer cells through inhibiting PI3K/Akt pathway. Also, the proteins involved in the Akt pathway, including PTEN, p-PTEN, and pT308-Akt, were suppressed and the expression of RASSF1 gene, Mst1, as well as LATS1 were increased. 26 Although lots of evidence supports that DIM and Akt pathway are closely linked in the occurrence and development of several human cancers, 27,28 whether DIM could possess antitumor effects in malignant melanoma cells and whether the PTEN/Akt pathway is involved is unclear.
In this study, the antitumor effects of DIM in malignant melanoma cells and the related mechanisms were examined.
First, we found that DIM exerted proliferation-inhibitory and apoptosis-induced effects in malignant melanoma A375 cells, but did not have these effects in A2058 cells. Given the major interest in identifying how DIM exerts its effects in A375 malignant melanoma cells, we examined the expression levels of proteins in cancer-related pathways. We found that DIM could increase the expressions of PTEN and Akt (Thr308) and inactivate Akt signaling. It has been reported that A375 cells are PTENWT cells, whereas A2058 cells are PTENmut cells. Furthermore, DIM has been shown to have anticancer properties in hepatocellular carcinoma cells through upregulating PTEN and inhibiting phospho-FAK (Tyr397). 29 We found no difference between control and DIM treatment in A2058 PTENmut cells. Therefore, we speculated that DIM exerted proliferation-inhibitory and apoptosis-induced effects in human malignant melanoma cells through PTEN/Akt signaling.
To investigate whether the PTEN and Akt signaling pathways contributed to the proliferation-inhibiting and apoptosis-inducting effects of DIM, PTEN-siRNA, and Flag-PTEN plasmids were constructed to interfere with the expression of PTEN and phosphorylated PTEN. We found that knockdown of PTEN in A375 PTENWT cells significantly attenuated the inhibitory effect of DIM on proliferation and DIM-induced apoptosis. Furthermore, overexpression of PTEN in A2058 PTENmut cells could augment DIM-mediated inhibition of proliferation and DIM-induced apoptosis.
Bcl-2 and Bid can influence the structure and permeability of mitochondrial permeability transition pore through their expression levels and subsequently promote mitochondria-mediated cell death. 30,31 To clarify whether the mitochondrial pathway is involved in the changed expression of PTEN and the apoptosis of malignant melanoma cells induced by DIM, we investigated the levels of mitochondrial pathway proteins. We found that DIM increased the expression levels of mitochondrial pathway proteins. The DIM-mediated changes in △ψm and the expressions of mitochondrial proteins indicated that DIM induced apoptosis in human malignant melanoma cells by activating the mitochondrial pathway through PTEN/Akt signaling.
Collectively, these results support the view that DIM exerted proliferation-inhibitory and apoptosis-inducing effects in human malignant melanoma cells by activating the mitochondrial pathway by promoting the expression of PTEN and subsequently inactivating Akt signaling. Because PTEN can inhibit Akt signaling by inhibiting the phosphorylation of Akt, the activation of PTEN and subsequent inactivation of Akt signaling may result in decreased expression of Bcl-2, and changes in the structure and permeability of mPTP. Such changes release Cyt C and AIF to form the apoptosome, which activates mitochondrial apoptosis (Fig. 7).

The roles of PTEN/Akt-induced activation of mitochondrial pathway in DIM-induced apoptosis in malignant melanoma cells.
In conclusion, this study demonstrated that DIM can increase PTEN expression and subsequently inactivate Akt signaling as well as activate the mitochondrial pathway in malignant melanoma cells, which contributed to DIM-induced apoptosis.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this study.