
Abstract
Sword bean has been known as a traditional medicinal plant to treat cancer, sinus infection, and suppurative disease. It also possesses hypertension-relieving, antioxidation, and antibacterial effects. However, studies on the efficacy of sword bean are limited to mature beans. Few studies have focused on immature sword bean pod (ISBP). Therefore, this study aimed to investigate the anti-inflammatory effect of ISBP in RAW264.7 cells stimulated with lipopolysaccharide (LPS). After LPS-induced RAW264.7 cells were treated with ISBP at concentrations (0.5, 1, 2, and 5 mg/mL), levels of nitrite oxide (NO) and prostaglandin E2 (PGE2) production, protein, and mRNA levels of inducible nitric oxide synthase (iNOS) and cyclooxygenase-2 (COX-2), inflammatory cytokine secretion level, and nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) activity were determined. Under inflammatory conditions induced by LPS, ISBP reduced levels of inflammatory mediators NO and PGE2 by 60% and 23%, respectively. It also decreased protein and mRNA expression levels of iNOS and COX-2 known to synthesize inflammatory mediators. Inflammatory cytokines, interleukin (IL)-6, and IL-1β, levels were decreased, while interferon gamma level was increased by ISBP based on enzyme-linked immunosorbent assay (ELISA) and real time-polymerase chain reaction results. Finally, ISBP showed the ability to inhibit NF-κB activity. In conclusion, ISBP can alleviate inflammation by controlling inflammation-related substances, and may have efficacy as a healthful functional food and natural anti-inflammatory drug.
Introduction
Inflammation is one of the host defense mechanisms against exogenous pathogens and physical or chemical stimuli. 1 When inflammation occurs, inflammatory cytokines that regulate the immune response are secreted and nitrite oxide (NO) and prostaglandin E2 (PGE2) are produced. 2,3 Interleukin (IL)-1β is involved in the inflammatory response by recruiting neutrophils into infected areas to express IL-6. 4 Interferon gamma (IFN-γ) can reduce autoimmune disease and allergic reactions. 5,6 NO is synthesized by inducible nitric oxide synthase (iNOS) induced by lipopolysaccharide (LPS). Its excessive secretion can lead to oxidative stress and inflammation. 7 PGE2 synthesized by cyclooxygenase (COX), which changes arachidonic acid to PG, is involved in tumor growth promotion, apoptosis suppression, and inflammatory response. 8 There are two isoforms of COX, COX-1 and COX-2. COX-2 is involved in the synthesis of PGE2. It is induced by growth factors and cytokines. Their expression levels are increased in cancer cells and inflammatory tissues. 9 iNOS and COX-2 are genes regulated by mitogen-activated protein kinase (MAPK) and nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB). 10 In general, NF-κB is in an inactive state and bound to nuclear factor of kappa light polypeptide gene enhancer in B cells inhibitor (IκB). It is activated when NF-κB is transferred into the nucleus due to phosphorylation of IκB. It is also activated by inflammatory cytokines, external stimuli, and oxidants. 11,12 Transcription levels of inflammatory mediators (iNOS, COX-2) mentioned above are increased when NF-κB is activated. In particular, NF-κB can activate the prostaglandin pathway and cause inflammation. 13 Anti-inflammatory drugs based on these inflammatory regulatory reactions have many complications and side effects such as gastric ulcers and osteoporosis. 14 Therefore, anti-inflammatory agents using natural food materials are needed.
Sword bean is a perennial vine plant originated endemic to tropical Asia, Africa, and India. It has a bow-shaped fruit. It is also called “Jakdu” or “Dodu.” It blooms from June to July and bears fruit from August to October. 15,16 All parts of sword bean, including its beans, leaves, pods, and roots are edible. It is composed of 35–45% starch and 22–29% protein. It also contains medicinal ingredients such as urease, canavanine, canavalia, and hemagglutinin. It has been used as a medicinal plant for relieving inflammation, cancer, constipation, sinusitis, and pyogenic diseases. 17 Recently, sword beans are known to possess antioxidative action to remove free radicals and protect against oxidative stress. 18 Antioxidant and ACE inhibitory activities of soybean pastes with added sword bean are higher than those of general soybean paste. 19 In addition, it has been reported that its addition to kochujang can increase the antioxidant activity and tyrosinase inhibitory effect more than general kochujang. 20 There are many studies on pharmacological efficacy and functional properties of sword bean. However, there have been no reports of immature sword bean pod (ISBP). Therefore, this study aims to investigate the anti-inflammatory effect of ISBP in RAW264.7 cells stimulated with LPS. This study might provide evidence for using ISBP as an ingredient in anti-inflammatory functional foods and drugs.
Materials and Methods
Materials
Dulbecco's modified Eagle's medium (DMEM) and penicillin-streptomycin antibiotics (P/S) were purchased from Gibco (Gaithersburg, MD). Fetal bovine serum (FBS) and secondary antibody were obtained from GenDEPOT (Barker, TX). LPS, dexamethasone (Dex) and dimethyl sulfoxide were obtained from Sigma-Aldrich (St. Louis, MO). Polyvinylidene difluoride (PVDF) membranes, sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE), and Chemi-doc image detector (ChemiDoc™ XRS+ System) were purchased from Bio-Rad (Hercules, CA). Primary antibodies for iNOS, COX-2, and β-actin were obtained Abcam (Cambridge, MA). NF-κB response element reporter vector was obtained from Promega (Madison, WI).
Sample extract preparation
Fresh ISBP (collected from Hwasun, South Korea) were chopped and boiled in distilled water. The ratio of solid to liquid was 1:3. The extraction was conducted at 100°C for 24 h using a reflux condenser. After extraction, the extract was filtered and lyophilized, and the lyophilized sample was stored at −20°C before analysis. The extraction yield of ISBP was 7.3% of fresh material.
Cell culture
Mouse macrophage cell line RAW264.7 was purchased from Korean Cell Line Bank (Seoul, Korea). RAW264.7 cells were cultured in DMEM containing 10% FBS and 1% P/S at 37°C in a 5% CO2 incubator.
Cell cytotoxicity
Intracellular adenosine triphosphate (ATP) indicate that cell survival rate was measured to evaluate cytotoxicity of ISBP. RAW264.7 cells (5 × 104 cells/well) were seeded onto 24-well plates. Then ISBP were treated at various concentrations (0.5, 1, 2, 5 mg/mL) and Dex (1 μM) as a positive control at 37°C for 24 h in a 5% CO2 incubator. After 24 h, luminescence was measured using Cell Titer-Glo® Luminescent Cell Viability Assay Reagent (Promega).
NO production
NO production by cells treated with ISBP was measured using Griess reagent (Promega). RAW264.7 cells (1 × 105 cells/well) were cultured in 96-well plates for 2 h. They were then treated with LPS (1 μg/mL) and ISBP extracts (0.5, 1, 2, 5 mg/mL) and Dex (1 μM) at 37°C for 24 h with 5% CO2. After 24 h, sulfanilamide and N-1-napthylethylenediamine dihydrochloride solution were sequentially added to the cell supernatant and each solution was reacted at room temperature for 10 min. Then the absorbance of the reaction mixture is measured at 540 nm.
PGE2 measurement
The PGE2 production in the supernatant of cells treated with LPS, ISBP, and Dex for 24 h was measured using PGE2 ELISA Kit (Abcam). Cell supernatants were added to goat anti-mouse IgG-coated 96-well plate. Then 50 μL of PGE2 conjugate and antibody were added and incubated for 2 h at room temperature with shaking at 500 rpm (minishaker, MS-1, IKA®-Werke GmbH & Co. KG, Staufen, Germany). After incubation, plates were washed with wash buffer. After adding 200 μL pNPP substrate, the plate was incubated for 45 min at room temperature. Absorbance was measured at wavelength of 405 nm after adding stop solution.
Real-time polymerase chain reaction
Expression levels of inflammatory genes (iNOS, COX-2, IL-6, IL1-β, and IFN-γ) were measured to confirm the anti-inflammatory effect of ISBP. RAW264.7 cells were treated with ISBP and Dex for 24 h after stimulation with LPS (1 μg/mL) for 1 h. After cultured cells were washed with PBS, total RNA was extracted using RNeasy Mini Kit (Qiagen, Valencia, CA) and cDNA was synthesized using M-mLV Reverse Transcriptase (Promega). Then inflammation-related genes expression was evaluated by real-time polymerase chain reaction (PCR) using synthesized cDNA as template and 2 × SYBR Green Master Mix on a Rotor-Gene Q real-time thermal cycler (Qiagen). Gene expression levels were normalized against GAPDH and expressed as fold-change value relative to a negative control group (cells not treated by LPS or ISBP).
Western blot analysis
After treatment with LPS, ISBP, and Dex for 24 h, cells were lysed with lysis buffer (0.5% NP-40, 0.5% Triton X-100, 0.1% sodium deoxycholate, 50 mM Tris-HCl, 150 mM sodium chloride, 1 mM EDTA). Lysed cells were left on ice for 40 min. The supernatant obtained after centrifugation (12,000 g, 4°C, 20 min) was quantified using the Bradford method. Proteins were separated by 4–20% SDS-PAGE, transferred to PVDF membrane, blocked with skim milk for 1 h, and then incubated with primary antibody (iNOS, COX-2, β-actin) at 4°C overnight. After incubation, cells were washed with 1 × TBST and incubated with secondary antibody for 1 h. Band was developed using ECL kit (Pierce, Rockford, IL) and band image was captured with a Chemi-doc image detector.
Measurement of inflammatory cytokine
Levels of inflammatory cytokines (IL-6, IL-1β, and IFN-γ) in supernatants of cells after treatment with LPS, ISBP, and Dex were measured using commercially available ELISA kits (R&D Systems, Minneapolis, MN) following the manufacturer's instructions. In brief, cell supernatant was added into 96-well plates coated with specific cytokine monoclonal antibody and incubated at room temperature for 2 h. After incubation, the conjugate was added and reacted for 2 h and the substrate for 30 min. Absorbance of inflammatory cytokine was then measured at wavelength of 450 nm.
Luciferase assay for NF-κB activity
RAW264.7 cells (5 × 104 cells/well) were seeded onto 24-well plates and incubated for 24 h. NF-κB response element reporter (0.1 μg/well) and TransFast Transfection Reagents (Thermo Fisher Scientific, Waltham, MA) were added to transfection medium. At 16 h after transfection, the medium was replaced with fresh medium and further incubated at 37°C for 48 h. Transfected cells were treated with ISBP and Dex for 2 h and then stimulated with LPS for 1 h before measuring NF-κB activity in cells using Dual-Glo Luciferase Assay (Promega) using luminometer (Berthold Technologies GmbH, Vienna, Austria).
2.11. High-performance liquid chromatography analysis of free phenolic acid
Free phenolic acid was analyzed as described by Krygier et al., 21 with slight modifications. In brief, 5 g of lyophilized ISBP was extracted with 100 mL of 70% methanol/acetone (1:1, v/v) three times. The supernatant was collected after centrifuging at 9000 g for 10 min at 4°C followed by evaporation at 45°C under vacuum to 90 mL. The concentrate was extracted with 90 mL of hexane after adjusted to pH 2.0 with 5N HCl. The water phase was further extracted three times with diethyl ether/ethyl acetate (DE/EA, 1:1, v/v). Combined DE/EA extracts were dehydrated with anhydrous sodium sulfate and then evaporated under nitrogen gas. Dried samples were reconstituted in 80% methanol and analyzed by high-performance liquid chromatography (HPLC). HPLC system was constructed with Shiseido Nanospace SI-2 series containing two pumps (3201), a degasser (3202), an auto sampler (3023), and a UV-visible detector (3002). Then 10 μL of analyte was injected into an analytical column (Shiseido, Capcellpak C18 UG120 5 μm, 4.6 × 250 mm). Free phenolic acids were detected by monitoring the eluent at 270 nm. Mobile phase was consisted of 0.1% formic acid in distilled water (mobile phase A) and HPLC grade methanol (mobile phase B). Solvent flow rate was maintained at 0.8 mL per minute. The following gradient conditions were used: 15% B for 0–2 min, 20% B for 3–25 min, 40% B for 26–36 min, 45% B for 37–56 min, 100% B for 57–60 min, and 15% B for 61–65 min.
Statistical analysis
In this study, all data are expressed as mean ± standard error of mean. All statistical analyses were performed using SPSS version 12.0.0 (IBM Co., Armonk, NY). Significance between mean values of experimental groups was analyzed by analysis of variance. Significance between LPS treatment group and the no-LPS treatment group was determined using Student's t-test. Significant difference was considered at P < .05.
Results
Evaluation of cytotoxicity of ISBP treatment
ATP is an indicator of metabolic activity and survival rate of cells. It is proportional to the number of surviving cells. 22 As a result of measuring ATP levels in RAW264.7 cells treated with ISBP, cell viability was revealed to be 80% after treatment with all concentrations of ISBP. Therefore, subsequent experiments were performed with ISBP under 5 mg/mL (Fig. 1).
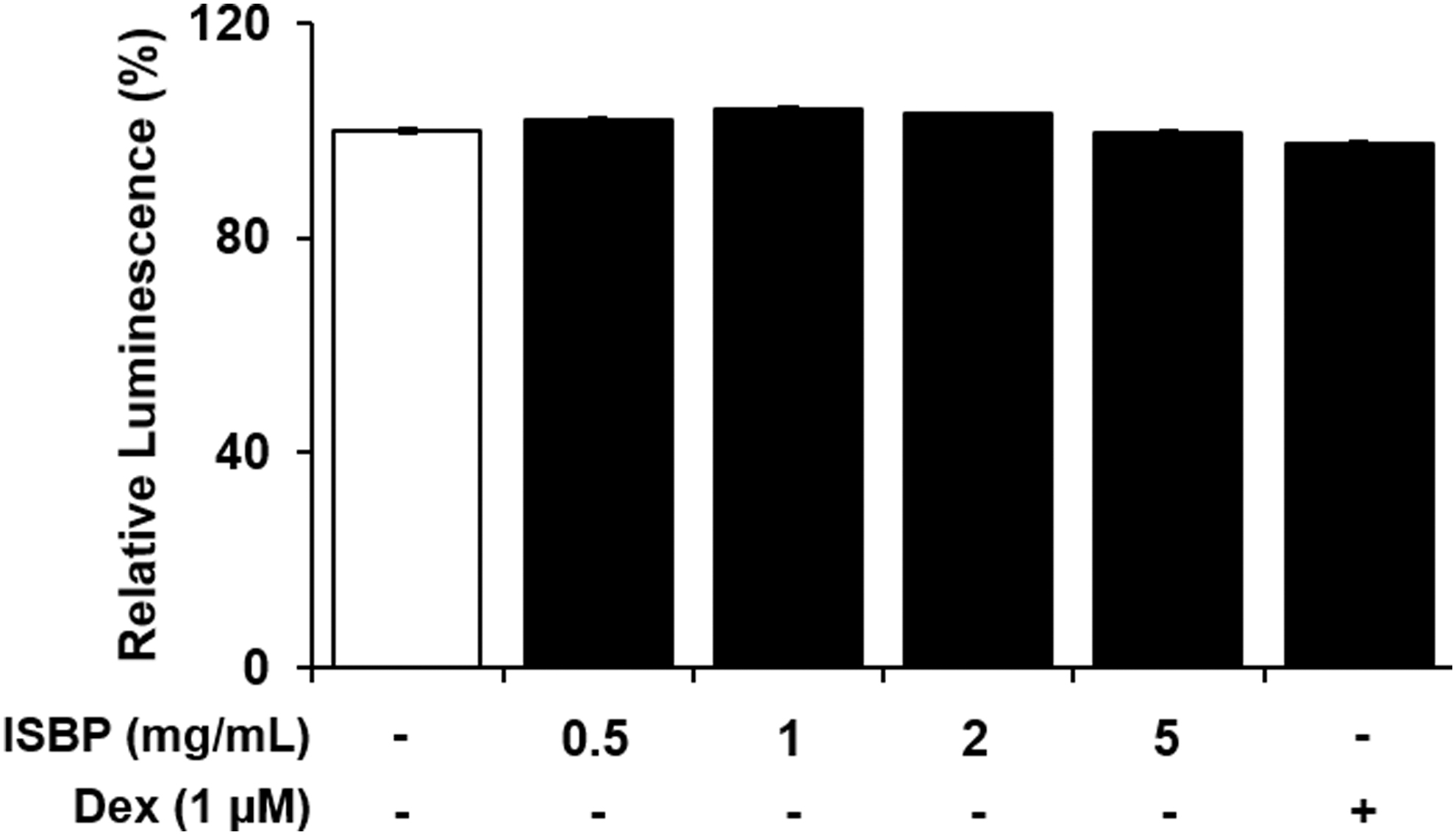
Effect of ISBP extract and Dex on the cytotoxicity of RAW264.7 cells. ISBP was treated at various concentrations for 24 h. Data are expressed as mean ± SEM. ISBP, immature sword bean pod. Dex, dexamethasone; SEM, standard error of mean.
Inhibitory effect of ISBP on NO and PGE2 production
Next, we investigated the inhibitory effect of NO and PGE2 on ISBP in LPS-induced cells. NO production was significantly increased by LPS treatment. NO production was dose-dependently decreased by ISBP treatment at 24, 19, 14, and 10 μM, respectively (Fig. 2A). In addition, the amount of PGE2 produced after LPS treatment was significantly decreased by ISBP (Fig. 2B). Especially, we observed higher inhibitory NO production than Dex in high concentrations (2 and 5 mg/mL) of ISBP extract. Therefore, ISBP is thought to be effective in the treatment and prevention of chronic inflammatory diseases.

Effect of ISBP extract and Dex on
Effect of ISBP on iNOS and COX-2 expression
Since ISBP reduced inflammatory mediators such as NO and PGE2, the effect of mRNA and protein expression on iNOS and COX-2 known to synthesize inflammatory mediators was then investigated (Fig. 3). The iNOS and COX-2 proteins expression were increased in the LPS-stimulated cell compared to the untreated LPS group. However, the ISBP-treated group had significantly lower iNOS and COX-2 expression. The quantification results of protein expression of iNOS. COX-2 mRNA expression also showed the same tendency (Fig. 3A). iNOS was reduced by 61% and COX-2 was reduced by 60% at the cell treated with ISBP (5 mg/mL) compared to that in LPS-treated control (Fig. 3B). Taken together, these results suggest that the inhibitory effect on NO and PGE2 by ISBP extract was mediated by the inhibition of iNOS and COX-2 expression.

Effect of ISBP extract and Dex on LPS-induced iNOS and COX-2 production in RAW264.7 cells.
Effect of ISBP on production of inflammatory cytokines
Inflammatory cytokines are rapidly secreted during inflammatory conditions to regulate inflammation and pathogen removal. 7 However, if inflammation persists due to the infiltration of many pathogens, it can damage host tissues and cause sepsis and organ failure. 23 In this experiment, we confirmed that ISBP in inflammatory conditions could relieve inflammatory conditions by regulating inflammatory cytokines. Previous results have shown that ISBP can inhibit inflammatory mediators. Thus, secretion levels of inflammatory cytokines, including IL-6, IL-1β, and IFN-γ, were evaluated using enzyme-linked immunosorbent assay (ELISA) and real time-polymerase chain reaction (RT-PCR) (Fig. 4). LPS stimulation increased IL-6 and IL-1β mRNA expression levels by 23 and 424-folds, respectively. However, IFN-γ mRNA expression was decreased to 0.5-fold. After ISBP treatment, IL-6 and IL-1β productions were decreased, while those of IFN-γ were increased (Fig. 4A). In addition, the degree of inflammatory cytokine secretion after ISBP treatment was similar to mRNA expression level (Fig. 4B). These results suggest that ISBP can alleviate inflammation by regulating the secretion of inflammatory cytokine.

Effect of ISBP extract and Dex on LPS-induced cytokine production in RAW 264.7 cells.
Effect of ISBP on NF-κB activity inhibition
Normally, NF-κB is activated when inflammation is induced by LPS, reactive oxygen species, and cytokines. It is a key factor in controlling the inflammatory response. 24 NF-κB activity can increase expression levels of COX-2, iNOS, IL-6, and IL-1β, leading to inflammation and degenerative diseases. 25 –27 Therefore, Luciferase assay was performed to determine whether ISBP could affect NF-κB activity (Fig. 5A). As a result, NF-κB activity was increased by LPS in NF-κB response element reporter transfected cells. However, NF-κB activity was dose-dependently inhibited by ISBP. Figure 5B and 5C shows the effect of ISBP extract on p65 and IκB-α expression by RT-PCR and western blotting. The phospho p65 and phospho IκB-α were significantly reduced by ISBP extract.

Effect of ISBP extract and Dex on NF-κB activation in RAW264.7 cells.
Total free phenolic acid concentration of ISBP
Results of analysis of total free phenolic acid composition of the ISBP extract are shown in Figure 6. Nine free phenolic acids (ellagic acid, gallic acid, p-hydroxybenzoic acid, methyl gallate, protocatechuic acid, pyrogallol, salicylic acid, sinapic acid, and vanillic acid) in ISBP were identified. Their representative chromatograms are shown in Figure 6. Pyrogallol and sinapic acid, which are widely distributed in plants, were the most abundant detected phenol components in ISBP.

HPLC chromatograms of a standard mixture of phenolic acids (top) and profiles of ISBP extract. Peak 1, gallic acid; 2, protocatechuic acid; 3, methyl gallate, 4, p-hydroxybenzoic acid; 5, vanillic acid; and 6, benzoic acid. HPLC, high-performance liquid chromatography.
Discussion
We evaluated the anti-inflammatory activity of ISBP extract in LPS-induced RAW 264.7 cell. It is known that macrophages involved in inflammatory reactions are activated by several stimuli, including LPS to produce NO, cytokines, and PGE2, that can induce inflammation and promote the migration of immune cells to inflammation sites. 8 When inflammation occurs, the synthesis of inflammatory mediators NO and PGE2 increases as part of the inflammatory response. 1 NO produced from iNOS can induce chronic diseases such as cancer and arteriosclerosis and cause cell damage. PGE2 that can promote tumor growth and suppress apoptosis is produced by COX-2. 10 In this study, ISBP extract inhibited NO and PGE2 productions as well as mRNA and protein expression of iNOS and COX-2. These results are similar to those reported by Kim et al. 23 in an animal model of alcoholic gastritis, showing that sword bean extracts can decrease COX-2 and NF-κB expression.
NF-κB, a major transcriptional regulator of the inflammatory response, plays an important role in the development of the inflammatory process. 28 NF-κB exists in the cytoplasm as a heterodimer consisting of p50 and p65. It remains in its inactive form by creating a complex with inhibitor protein IκBα. When inflammatory stimulation occurs, IκBα is phosphorylated and NF-κB is released. NF-κB then migrates to the nucleus and upregulates inflammation-related genes such as iNOS, COX-2, and pro- and anti-inflammatory cytokines. 29 In the present study, the NF-κB, iNOS, and COX-2 production as inflammatory mediators were reduced by ISBP.. These results are consistent with data obtained in other studies performed on RAW macrophages, in which LPS-induced p65 and IκBα expression levels are decreased by different bioactive compounds. Calixto-Campos et al. 30 have shown that sword bean extract can reduce NF-κB expression in an alcoholic gastritis animal model. ISBP inhibited NF-κB activity and decreased expression levels of inflammatory mediators such as COX-2 and iNOS in the present study, suggesting that ISBP might possess anti-inflammatory activity. In addition, various drugs focusing on inhibition of NF-κB activity for the treatment of human inflammatory diseases have been developed. 31 However, they can cause side effects similar to nonsteroidal anti-inflammatory drugs (NSAIDs), such as gastrointestinal damage, renal syndromes, and respiratory effects, when they are taken for a long time. 32,33 Therefore, ISBP due to its inhibition of NF-κB activity has been developed as an anti-inflammatory natural material. It is expected to be commercialized as a natural anti-inflammatory agent with less side effects compared to NSAIDs.
Obtained results from the phenolic analysis in the present study are in agreement with previous studies reporting the presence of phenolic acids in botanical with anti-inflammatory activity. Han et al. 34 has reported that red sword bean composition contains more phenolic compounds such as methyl gallate, gallic acid, and ellagic acid with anti-inflammatory and antioxidant effects than soybean. The results of our study demonstrate that ISBP can inhibit iNOS and COX-2 production in a dose-dependent manner when LPS-induced RAW264.7 cells are treated with ISBP, as ISBP contains many phenolic compounds with anti-inflammatory properties. According to a previous study, pyrogallol can regulate proinflammatory expression in bronchial epithelial cells. 35 while sinapic acid can dose-dependently regulate TNF-α, IL-1β, NO, and PGE2 in LPS-induced RAW264.7 cells. 36 Huang et al. 37 have reported that p-hydroxybenzoic acid, a derivative of benzoic acid with controversial anti-inflammatory effects, is a main active compound with anti-inflammatory properties in mice. Methyl gallate, derivative of gallic acid, and gallic acid are also phenolic compounds with anti-inflammatory effects, They inhibit COX-2, regulate arachidonic acid metabolizing enzymes, and suppress the production of NO in LPS-induced RAW264.7 cells. 38 –42 Finally, protocatechuic acid can also suppress proinflammatory cytokines and inflammatory mediators in RAW264.7 cells. 43 –45 Lende et al. 44 have also reported that protocatechuic acid possesses analgesic and anti-inflammatory activities in a carrageenan-induced paw edema model. Consequently, ISBP is expected to be developed as an anti-inflammatory new material for health functional foods and natural anti-inflammatory drugs. However, a more precise study is required to identify signaling pathways involved in the anti-inflammatory action of abundant pyrogallol, sinapic acid, and salicylic acid among nine phenolic compounds found in ISBP extract.
In conclusion, ISBP seems to exhibit anti-inflammatory effects by inhibiting the production of NO and PGE2 and the expression of COX-2 and iNOS due to its phenolic compounds sinapic acid, ellagic acid, methyl gallate, and so on. Therefore, ISBP with anti-inflammatory effects might be useful as a natural material of NSAID that can simultaneously act on NO and PGE2 as well as a natural ingredient of health functional foods if the present results can be confirmed in human clinical trials.
Footnotes
Author Disclosure Statement
No competing financial interests exist
Funding Information
This study was carried out with the support of the Research Program for Agricultural Science & Technology Development (project number PJ01456202, National Institute of Agricultural Sciences, Rural Development Administration, Republic of Korea).