
Abstract
Emerging evidence has shown that flavonoids extracted from Artemisia have beneficial effects on metabolic disorders. However, whether and how jaceosidin ameliorates insulin resistance and diabetic nephropathy in type 2 diabetes mellitus is largely unknown. For 8 weeks, db/db diabetic mice were fed with or without jaceosidin. Oral jaceosidin supplementation reduced fasting blood glucose levels and insulin resistance through the upregulation of insulin receptor downstream pathways in the liver and skeletal muscles. While jaceosidin did not noticeably alter kidney filtration function, this dietary intervention contributed to attenuating the accumulation of advanced glycation end products in diabetic kidneys. The levels of VEGF-a (vascular endothelial growth factor-a) proteins in the diabetic kidneys were markedly diminished by jaceosidin treatments, which increased the expression and activity of Cu (copper) and Zn-SOD (zinc-superoxide dismutase). Therefore, it is suggested that jaceosidin supplementation elicits antidiabetic effects and treats diabetic nephropathy by augmenting insulin signaling, suppressing fibrosis, and enhancing antioxidant activity.
Introduction
Type 2 diabetes mellitus (T2DM) is a complicated and multifactorial endocrine disorder that results from metabolic dysregulation of carbohydrates, fats, and proteins. 1 This pathophysiological condition induces chronic hyperglycemia that leads to high morbidity and mortality rates. The prevalence of T2DM in developing and industrialized countries has been rapidly increasing, with 425 million individuals affected worldwide. 2 This number is expected to exceed 629 million by 2045. 2 Hence, treatments for this global health crisis are urgently sought.
T2DM is primarily associated with obesity and characterized by impaired glucose tolerance and high insulin resistance. 3 –5 Insulin resistance is attributed to the excessive accumulation of reactive oxidative species (ROS) that play a critical role in suppressing insulin responses in various tissues and organs, including the liver, fat, and skeletal muscles. 6,7 A critical metabolic function of insulin is the stimulation of glucose uptake into adipocytes and muscle tissue. Insulin modulates glucose metabolism by activating the insulin receptor substrate-1 (IRS-1)–phosphoinositide 3-kinase (PI3K)–protein kinase B (Akt) signaling pathways. In turn, glucose transporters (GLUT) translocate from intracellular membranes to the plasma membranes of muscle tissue and adipocytes prestimulated by insulin. Along with a reduction in glucose utilization and insulin resistance in T2DM, prolonged hyperglycemia results in life-threatening complications, such as neuropathy, retinopathy, and cardiovascular disorders. 8,9 In particular, diabetic nephropathy is the most frequent cause of end-stage renal disease and is characterized by the thickening of glomerular and tubular basement membranes, tubulointerstitial fibrosis, and elevations in serum creatinine and urea nitrogen. 10,11 While the number of key diabetic complications has markedly diminished, acute and chronic kidney disorders from T2DM have seldom been ameliorated. Therefore, special attention must be given to explore therapies for preventing and treating diabetes-associated renal disorders.
Oral antihyperglycemic synthetic drugs, synthetic drugs have several important limitations, such as high costs, and various side effects including abnormal liver function and heart failure. 12,13 Therefore, nontoxic natural agents with high efficacy and fewer adverse effects must be identified to help patients maintain the appropriate glycemic levels and ultimately conquer T2DM. Botanicals and their derivatives that originate from plants, algae, and lichens have been traditionally exploited for medicinal purposes. 14,15 There is compelling evidence to indicate that oral administration of plant-based flavonoids prevents or ameliorates hypertension, diabetes, and acute/chronic kidney disease, thereby exerting reno-protective and pharmacological activities against a diverse range of pathophysiological diseases. 11 The perennial herb Artemisia is used in traditional Korean medicines for coughs, dyspepsia, colic asthma, and rheumatism. 16 –18 Artemisia leaf extracts that are rich in several flavonoids, including jaceosidin, eupalitin, and eupafolin, 19 are reported to suppress human endometriotic cancer cells and enhance cellular regulation of glucose uptake in cultured L6 muscle cells. 20,21 Furthermore, the functional elements of Artemisia princeps are thought to have anticancer or anti-inflammatory properties. 22
Among flavonoids extracted from the Artemisia species, jaceosidin is one of the most pharmacologically active flavones and has diverse biological actions, including antioxidant activity. 23 It is well accepted that the oral administration of jaceosidin favorably reduced insulin resistance that results from prolonged hyperglycemic oxidative stress. 24,25 However, the intracellular signaling mechanisms by which jaceosidin intake ameliorates insulin resistance in T2DM have yet to be defined. In addition, the focus has to be placed on the therapeutic effects of jaceosidin on diabetic nephropathy to prevent adverse effects on the kidney and reduce mortality in patients with renal disorders. To address these questions, this study investigated the potential mechanisms underlying the antidiabetic effects of jaceosidin on impaired IRS-1–PI3K–Akt signaling pathways and compromised kidney dysfunction in T2DM.
Materials and Methods
Jaceosidin purification
Jaceosidin (4′,5,7-trihydroxy-3′,6-dimethoxyflavone) was isolated from the methanol extracts of the aerial parts of Artemisia argyi, as previously described. 26 Jaceosidin was purified through recrystallization using acetone–hexane. 1 H NMR spectra were recorded at 500 MHz using a HRMAS-FT-NMR.
Experimental animals and diets
Five-week-old male mice (C57BL/6J-db/db) were purchased from Central Laboratory Animal, Inc. (Seoul, Korea). After a 2-week acclimation, the db/db mice were randomly divided into 4 groups (n = 10 per groups) and fed the respective experimental diets for 8 weeks. The diets were prepared based on AIN-93G with or without jaceosidin (Central Lab Animal, Seoul, Korea). Jaceosidin was supplemented at either 0.005% or 0.02%, based on previous studies, 27 and rosiglitazone (Rosi) was used as a positive control. All mice were housed under specific pathogen-free laboratory conditions (25°C, 50% relative humidity) on a 12-h light–dark cycle, with free access to food and water. Food consumption and body weight were measured twice a week for calculation of the food efficiency ratios (FER). Overnight fasting glucose concentrations were monitored weekly using a glucometer (Accu-Check Perfoma; Roche, Seoul, Korea). All animal experiments were approved by the Medical Ethics Committee of Chung-Ang University (#13-0036).
Organ harvest and urine/blood sample collections
Upon completion of jaceosidin supplementation for 8 weeks, a 24-h urine collection was enforced. Mice were euthanized after an overnight fast. All organs and tissues were excised and frozen immediately for further analysis and blood samples were collected.
Western blot analysis
The liver, skeletal muscle, and kidney tissues were homogenized in a RIPA lysis buffer containing protease inhibitors and phosphatase inhibitors (Sigma–Aldrich, St. Louis, MO, USA). Cell debris was removed by centrifugation. Protein concentrations were measured using a Bradford assay kit (Bio-Rad, Hercules, CA, USA). The protein equivalents of samples were separated by 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to polyvinylidene fluoride or nitrocellulose membranes. Membranes were blocked with 5% skimmed milk in a Tris-buffered saline containing 0.05% Tween-20 for 1 h at room temperature, and then probed with antibodies against β-actin, GLUT4, phosphorylated-IRS, p-PI-3K, fibronectin, vascular endothelial growth factor-a (VEGF-a) (all from Cell Signaling, Danvers, MA, USA), p-Akt, p-p38, catalase, copper, zinc-superoxide dismutase (Cu, Zn-SOD), and manganese (Mn)-SOD (all from Santa Cruz Biotechnology, Dallas, TX, USA). The membranes were exposed to horseradish peroxidase-conjugated secondary antibodies (Pierce, Waltham, MA, USA). Immunoreactive bands were detected by enhanced chemiluminescence solution (Animal Genetics, Seoul, South Korea) and exposed to X-ray film.
Blood urea nitrogen, albumin/creatinine ratio, and advanced glycation end products measurement
Blood urea nitrogen (BUN) and albumin/creatinine ratios were measured enzymatically (Cayman Chemicals, Ann Arbor, MI, USA). The levels of advanced glycation end products (AGEs) in the kidneys were measured using an ELISA kit (Cell Biolabs, Inc., San Diego, CA, USA). All samples were assayed in triplicate and analyzed using a microplate spectrophotometer (SpectraMax 340; Molecular Devices, Sunnyvale, CA, USA).
Thiobarbituric acid-reactive substances
Lipid peroxidation in kidney was measured by the thiobarbituric acid-reactive substances (TBARS) method. Kidney tissues were homogenized in Tris-buffer. Homogenates were mixed with SDS, acetic acid, and TBA solution and boiled at 95°C for 1 h. The cooled mixture and n-butanol were vortexed and centrifuged. The upper layer was measured at 532 nm and concentrations were determined using a malondialdehyde standard curve.
Antioxidant enzyme activity
SOD and glutathione peroxidase (GPx) activities were assessed using a colorimetric assay kit (Cayman Chemicals). The activities of SOD and GPx were presented as units per milligram of protein.
Statistical analysis
Data obtained from at least three independent experiments are presented as mean ± standard deviation. Statistical analyses were conducted using SPSS Statistics 23.0. One-way analysis of variance (ANOVA) was performed to evaluate statistical differences where there were more than three different treatments. If significant differences were detected by one-way ANOVA, Duncan's multiple range tests were used for post hoc testing. Statistical significance was set at P < .05.
Results
Effects of jaceosidin on body and organ weight changes, and FER
We first determined whether jaceosidin supplementation affects the FER in db/db mice (Table 1). Control and jaceosidin administration groups exhibited a less weight gain compared with Rosi group. The relative body weight gain (%) of mice treated with or without jaceosidin was 28.44% (control), 20.96% (0.005%), and 39.52% (0.02%), whereas that of db/db mice fed with Rosi was 78.72%. Conversely, since the control and jaceosidin groups showed lower total body weight gains, the FER in those groups were markedly lower than in the positive control group.
Effect of Jaceosidin on Changes in Body Weight, Food Intake, Food Efficiency Ratio and Organ Weights in db/db Mice
All data are mean ± SD. Values not sharing the same letter are significantly different (P < .05).
J-5, J-20, and Rosi are defined as 0.005% jaceosidin, 0.02% jaceosidin, and 0.001% rosiglitazone, respectively.
FER: food efficiency ratio (total body weight gain/food intake × 100).
SD, standard deviation.
Hyperglycemic conditions in T2DM enlarge or diminish the size of organs, such as the liver, spleen, and kidney, so this study attempted to examine the hypothesis that jaceosidin administration alters organ weights. The internal organ weights were normalized to body weight. In contrast to the positive control treatment, the jaceosidin dietary intervention did not significantly affect the weights of organs or tissues in db/db mice including the liver, spleen, kidneys, epididymal adipose tissue, and perirenal adipose tissue.
Effects of jaceosidin on impaired fasting blood glucose in diabetic mice
We assessed whether jaceosidin supplementation mediates fasting blood glucose and glucose/insulin tolerance. Basal fasting blood glucose levels (i.e., 0 week) in all of the control and intervention groups were close to or over 200 mg/mL, indicating a diabetic condition (Fig. 1). No differences in the fasting blood glucose levels among the groups were observed at the start of the study. While the diabetic control and jaceosidin (0.005% and 0.02%) groups displayed progressive increases in fasting blood glucose levels during the first 4 weeks, hyperglycemia was substantially ameliorated after 5 weeks of 0.02% jaceosidin supplementation. In addition, 0.005% and 0.02% jaceosidin interventions sustained blood glucose levels after 5 weeks, whereas blood glucose levels continued to increase in the control group.
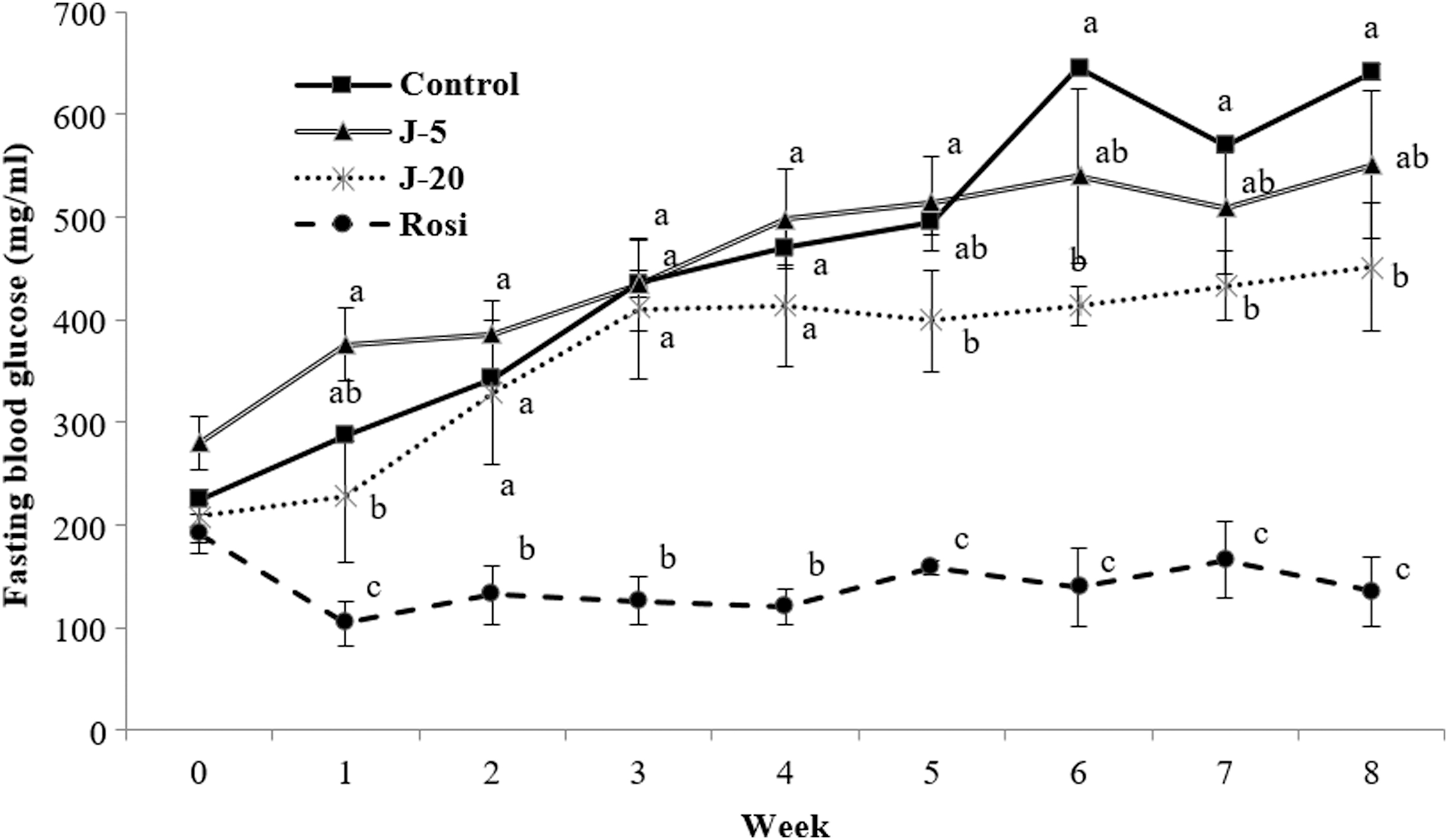
Effects of jaceosidin on fasting blood glucose levels in db/db mice. Five-week-old db/db mice were treated with 0.005% jaceosidin (J-5), 0.02% jaceosidin (J-20), or 0.001% rosiglitazone (Rosi) for 8 weeks. Fasting blood glucose was measured once a week for 8 weeks Statistical analysis was performed using ANOVA. Values not sharing the same letter are significantly different (P < .05). ANOVA, analysis of variance.
Effects of jaceosidin on the expression of insulin signaling-related proteins in the liver and skeletal muscles
The phosphorylation of the downstream signaling proteins involved in glucose utilization was examined in the absence or presence of jaceosidin administration in the liver (Fig. 2) and skeletal muscle (Fig. 3) of db/db mice. Both oral treatments considerably stimulated insulin signaling. Interestingly, jaceosidin intervention exhibited concentration-dependent increases in the phosphorylation of IRS-1, PI3K, Akt, and p38 proteins in the liver, which were similar or higher than the effects of Rosi on the phosphorylation of those proteins (Fig. 2B–E). Next, in the skeletal muscle, the phosphorylation of PI3K observed in the jaceosidin-treated groups was significantly higher than that in the control group. While there was no significant effect of jaceosidin on the post-translational modification of IRS-1, Akt, and p-38 proteins, GLUT4 expression was considerably augmented by jaceosidin 0.02% supplementation in skeletal muscles.
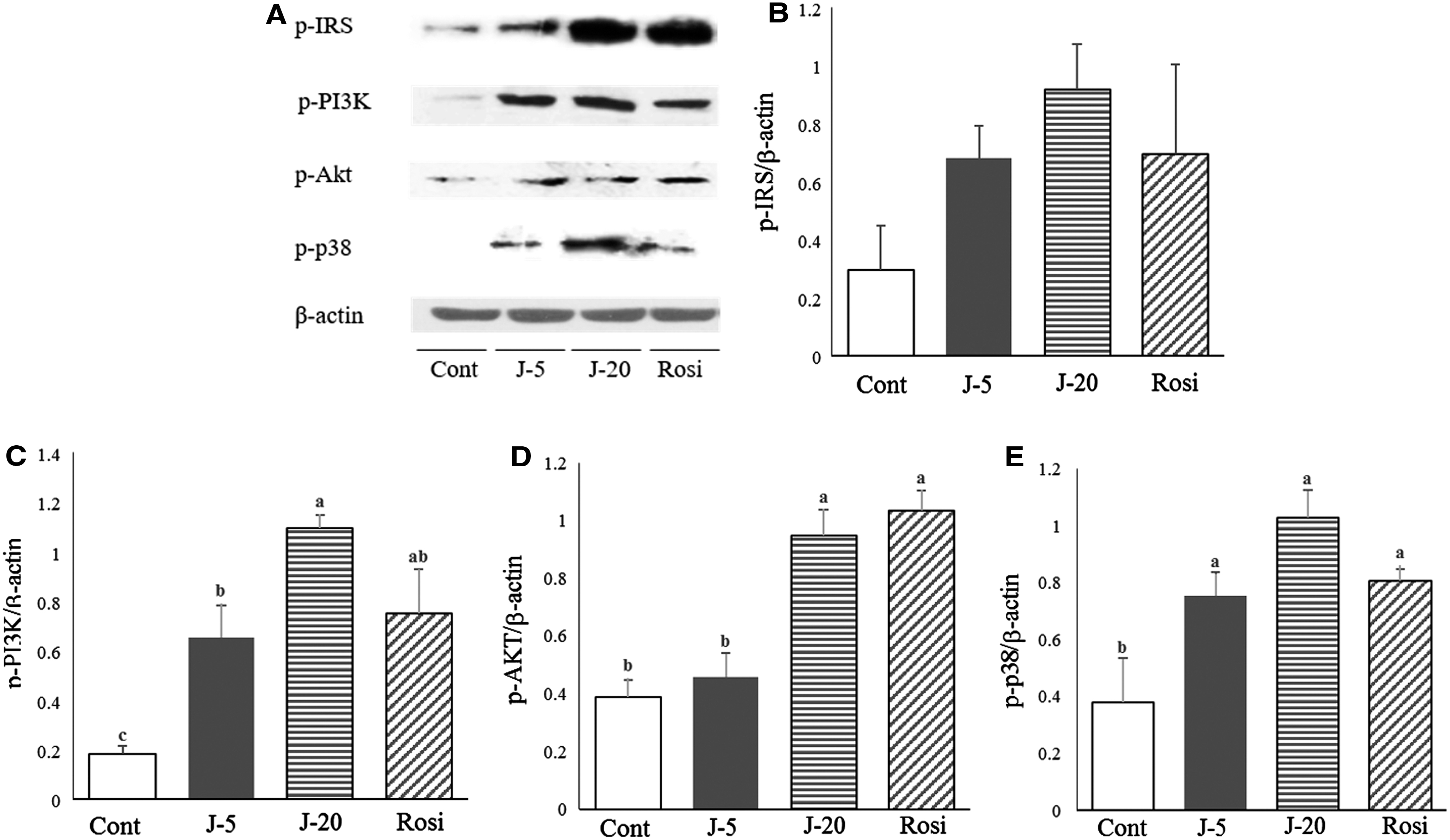
Effects of jaceosidin on the expression of insulin signaling regulatory proteins in the liver. Protein expressions of insulin signaling regulatory proteins in the liver were investigated using Western blotting

Effects of jaceosidin on the expression of insulin signaling regulatory proteins in skeletal muscles. Protein expressions of insulin signaling regulatory proteins in skeletal muscles were examined using Western blotting
Effects of jaceosidin on kidney function and fibrosis
Next, we tested whether the decrease of fasting glucose by jaceosidin supplementation protects against diabetic nephropathy (Fig. 4). The BUN, albumin-to-creatinine ratio, and AGEs were monitored in db/db mice fed with or without jaceosidin to assess the function of kidneys. The two different doses of jaceosidin did not elicit marked changes in BUN (Fig. 4A). The albumin-to-creatinine ratio was substantially lower in the jaceosidin intake groups than the control group, although the differences were not statistically significant (Fig. 4B). AGEs in the kidneys of diabetic mice were profoundly attenuated by jaceosidin supplementation (Fig. 4C).
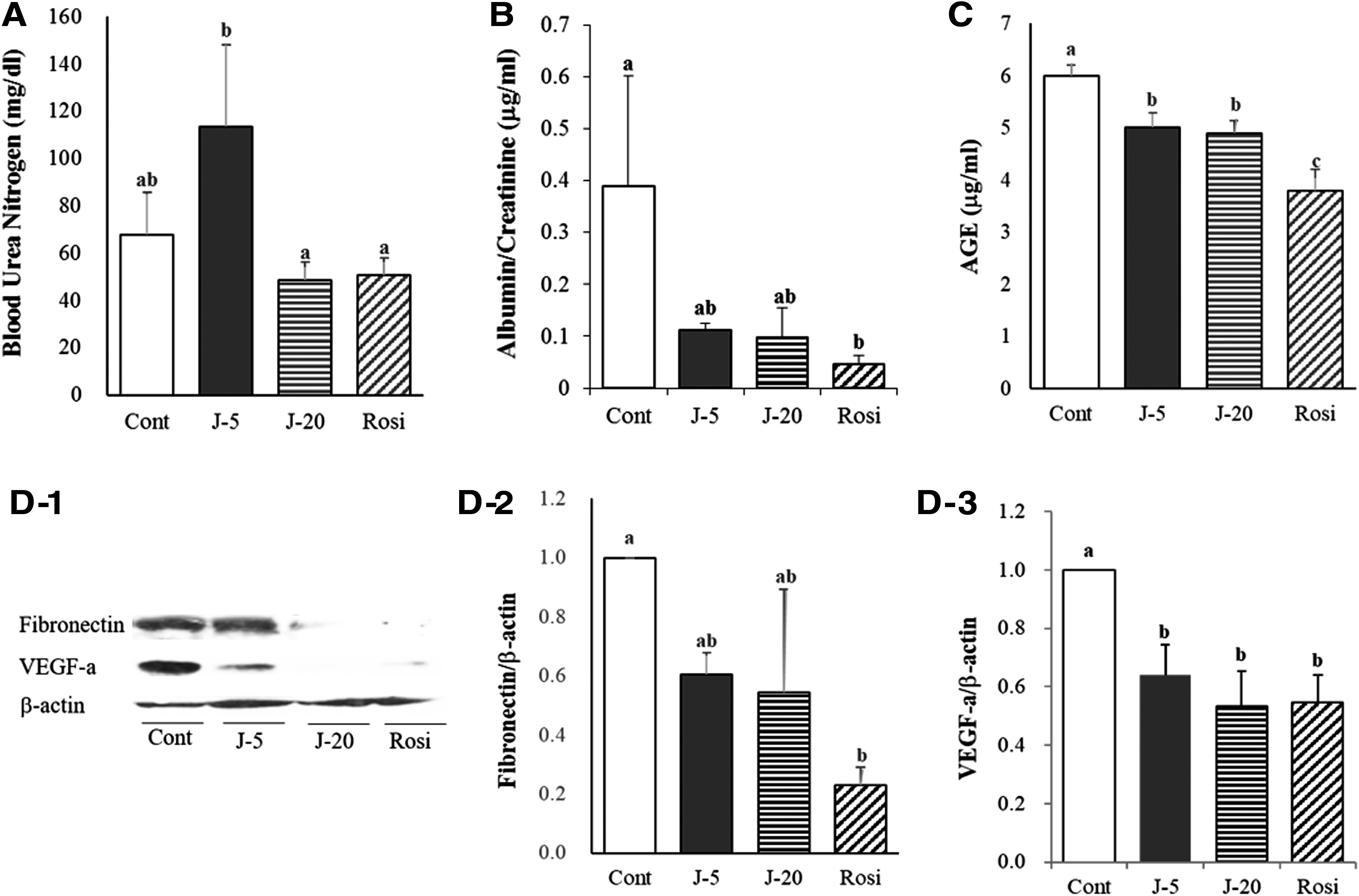
Effects of jaceosidin on kidney function
In the kidneys, an exaggerated accumulation of extracellular matrix (ECM) proteins, including fibronectin is the hallmark of diabetic nephropathy 28 and excessive VEGF-a expression is observed in pathophysiological conditions, such as tumors or diabetes. 29,30 Hence, the present investigation scrutinized whether the expression of these proteins in the kidneys of db/db mice is modulated by jaceosidin supplementation. The expression level of fibronectin was decreased in the kidneys of diabetic mice fed with jaceosidin (Fig. 4D-2). In addition, oral administration of 0.005% and 0.02% jaceosidin provoked significant decreases in VEGF-a expression (Fig. 4D-3).
Effects of jaceosidin on the expression and activity of endogenous antioxidant enzymes in the kidney
We sought to elucidate whether antidiabetic nephropathy effects of jaceosidin is related to the endogenous antioxidant system. Firstly, the concentration of TBARS, which is a byproduct of lipid peroxidation, was assessed in the diabetic kidney. We found that 0.02% jaceosidin administration noticeably reduced the oxidative degradation of lipids (Fig. 5A). Then, we assessed the effects of jaceosidin on the upregulation of endogenous antioxidants, including catalase and SOD, at the translational levels. While jaceosidin interventions did not alter catalase expression (Fig. 5B-2), Cu, Zn-SOD in the diabetic kidney was markedly upregulated by the relatively higher dosage of jaceosidin (Fig. 5B-3). However, the expression of Mn-SOD was not changed in the presence of both dietary interventions (Fig. 5B-4). Analysis of the activity of endogenous antioxidant enzymes revealed that SOD activity was strikingly augmented by jaceosidin administration (Fig. 5C), whereas the enzymatic activity of GPx was not affected by jaceosidin (Fig. 5D). Therefore, it is plausible that the jaceosidin-mediated increase in SOD activity may be attributed to the augmented expression of Cu, Zn-SOD in the kidney.
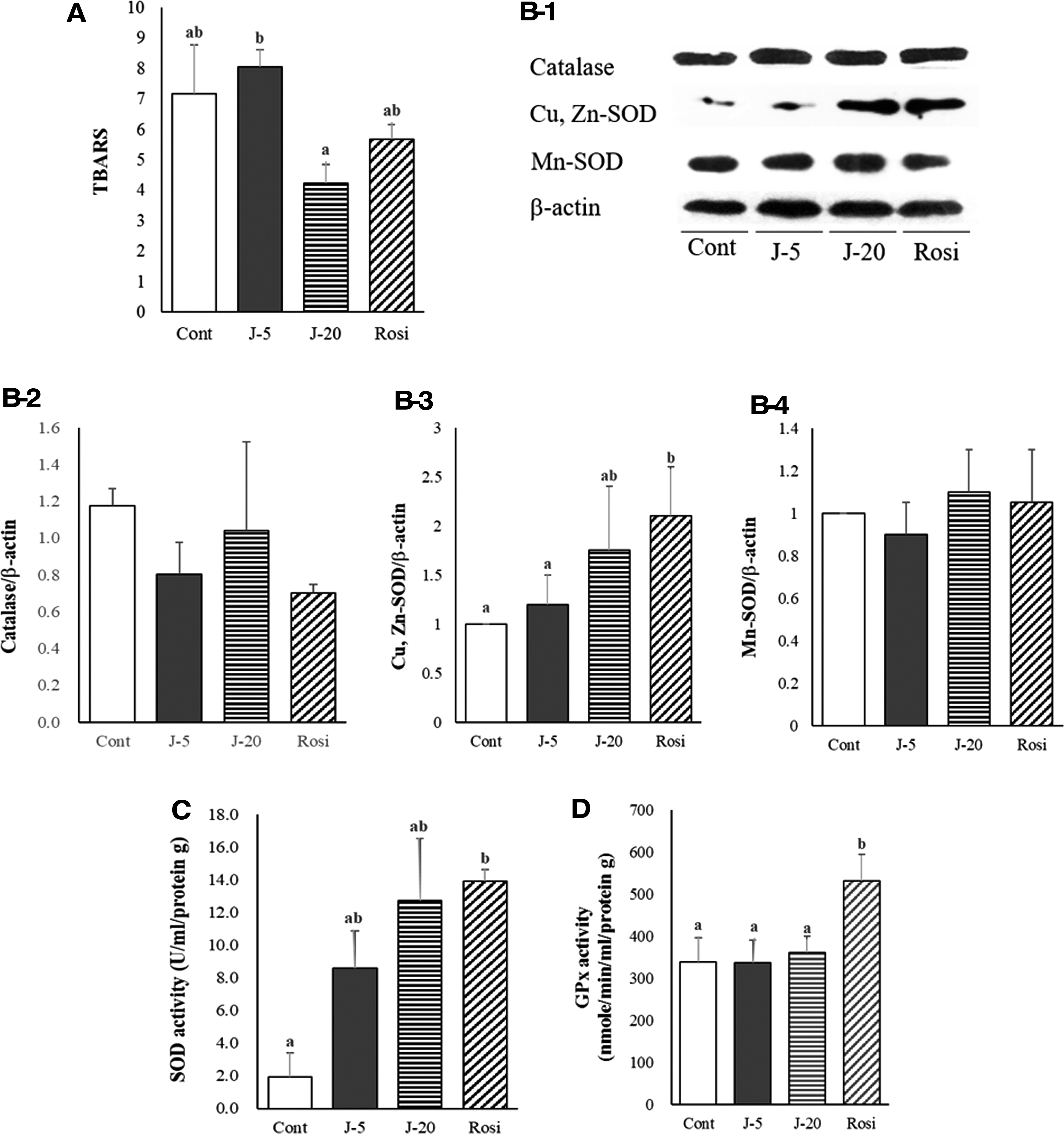
Effects of jaceosidin supplementation on TBARS, and antioxidant enzyme expression and activities in kidney tissue. TBARS concentration was measured in the kidney
Discussion
This study revealed that jaceosidin attenuates fasting plasma glucose levels and insulin resistance by stimulating insulin signaling pathways (i.e., IRS-1–PI3K–Akt). Furthermore, this investigation revealed that jaceosidin supplementation ameliorates diabetic nephropathy by suppressing fibrosis in the kidney and enhancing the expression and activity of Cu, Zn-SOD. Collectively, this study provides consistent evidence for the potential role of jaceosidin in mitigating hyperglycemia and highlights that jaceosidin has potential therapeutic effects to combat diabetic renal complications.
Rosiglitazone, in the thiazolidinedione class, was conventionally used as an antidiabetic drug and exhibits a meaningful blood glucose lowering effect. However, it also causes adverse events that is common for synthetic pharmacological drugs, rosiglitazone was found to induce weight gain or obesity by considerably increasing body fat. 31 Indeed, rosiglitazone intervention for 8 weeks markedly increased body weight and FER in this study. As obesity is an independent risk factor for cardiovascular diseases and metabolic syndromes, natural bioactive substances need to be used to reduce adverse events of antidiabetic drugs. In this context, jaceosidin used in this study did not cause a significant weight gain. It is, therefore, indicated that jaceosidin may be a promising and suitable agent to treat T2DM.
This study provides evidence that jaceosidin supplementation affects fasting blood glucose levels, glucose tolerance, and insulin resistance in T2DM models. A prior study demonstrated that Artemisia extracts have antihyperglycemic effects and enhance insulin sensitivity. 32 The dietary jaceosidin extracted from A. princeps consistently and profoundly lowered the fasting blood glucose level after 4 weeks. It may, therefore, be postulated that oral jaceosidin administration augments insulin action and, subsequently, evokes the hypoglycemic condition through further stimulation of insulin-dependent biological actions.
The antidiabetic effects of medicinal plants can be explained through various mechanisms such as enhancing insulin secretion, stimulating peroxisome proliferator-activated receptors (PPARs), augmenting endogenous antioxidant activity, increasing the capacity of glucose transporters, and reducing AGE production. 33,34 Thus, multiple approaches were used in this study to address how the natural flavone jaceosidin mitigates insulin resistance and treats diabetic nephropathy. Therefore, we initially concentrated on the impacts of jaceosidin on the insulin signaling pathways in the liver and skeletal muscles of diabetic animal models. The IRS-1–PI3K–Akt cascade, followed by the activation of insulin receptors, plays a crucial role in glucose metabolism. 35 This study consistently suggested that the fundamental IRS-1–PI3K–Akt pathway was promoted by jaceosidin supplementation in the liver and skeletal muscles, implicating that this upregulation contributes to improving insulin-mediated glucose uptake and reducing insulin resistance. Since IRS proteins play a vital role in initiating the downstream pathways of insulin signaling (i.e., PI3K-Akt-p38), it is worthwhile discussing putative regulatory mechanisms underlying flavonoid-dependent expression of IRS proteins. It is well known that the expression of IRS proteins is regulated by proteasomal degradation 36,37 that may be specifically controlled by mammalian target of rapamycin (mTOR) through the activation of S6 ribosomal protein kinase or protein kinase C. 38,39 It was found that mTOR-induced hyperphosphorylation on serine/threonine residues of IRS proteins results in proteasomal degradation and in turn decreases the expression of those proteins. 40 Dietary flavonoids found in natural products were suggested to elicit proteasome inhibition, which is an appealing therapeutic approach to diabetes and cancers. 41,42 Emerging evidence has recently indicated that flavonoids suppress mTOR activity. 43 Thus, although the present investigation did not directly address whether jaceosidin is involved in the regulatory mechanisms for the expression of insulin signaling cascades, it may be assumed that the dietary flavonoid jaceosidin supplementation may interrupt the serin/threonine hyperphosphorylation of IRS proteins and the degradation of downstream signaling.
Glucose transporters, such as GLUT4, are indispensable for cellular glucose utilization. Akt is well known to evoke the translocation of GLUT4 after the activation of insulin receptors. 44 Although the translocation of glucose transporters to the cell membrane was not directly observed in this study, we did find that jaceosidin augmented the expression of GLUT4 in the skeletal muscle. Previous investigations have indicated that PPARγ is involved in PI3K–Akt signaling. 20,45,46 Moreover, p38 mitogen-activated protein kinase (MAPK) has been reported to presumably play a critical role in insulin-mediated glucose uptake in the liver and skeletal muscle. 47 The p38 MAPK also modulates the glucose transporter system and enhances GLUT4 activity in insulin-sensitive skeletal muscles, 48 –50 which was consistent with our findings that p38 phosphorylation in soleus muscles increased after dietary jaceosidin treatment. It is therefore speculated that oral jaceosidin administration may provoke PPARγ–PI3K–Akt-dependent or p38 MAPK-mediated increases in GLUT4 expression and trafficking to the plasma membranes, thereby enhancing glucose uptake and ameliorating insulin resistance.
Diabetic nephropathy commonly occurs under prolonged hyperglycemic conditions, so this study sought to identify diabetic kidney dysfunctions and pathological mechanisms. Furthermore, the novel finding of the present study is that oral jaceosidin administration downregulated ROS-generating proteins and enhanced endogenous antioxidant SOD expression and activity, which attenuated diabetic kidney damage. Previous studies observed that T2DM induces hyperplasia-mediated enlargement of the kidney and disturbs glomerular filtration. 51 In addition, high levels of BUN and a high albumin-to-creatinine ratio are associated with impaired glomerular filtration and kidney damage. 52,53 While kidney size and BUN were not altered by long-term jaceosidin treatment in this study, the intake of the natural flavonoid contributed to the lowering of the albumin-to-creatinine ratio. It is conceivable that jaceosidin treatments may be additionally suitable for treating diabetic kidney injuries rather than suppressing changes in kidney morphology and enhancing filtration functions. Furthermore, T2DM is known to elevate AGEs that generate ROS and upregulate VEGF in mouse or rat kidneys. 54,55 Nicotinamide adenine dinucleotide phosphate oxidase enzymes play a critical role in the pathogenesis of diabetic nephropathy, and diabetes-induced proteinuria may result from excessive VEGF that augments glomerular permeability. 55,56 In terms of these adverse effects of hyperglycemic conditions, the results revealed that jaceosidin markedly decreased AGE, implying that the levels of ROS may be attenuated in the diabetic kidney. This implication was consistently supported by the jaceosidin-induced reduction in the levels of TBARS, a byproduct of lipid peroxidation. Collectively, we found that jaceosidin ameliorated the expression and activity of the endogenous antioxidant Cu, Zn-SOD enzyme in the diabetic kidney. It is therefore plausible that jaceosidin supplements may have potential therapeutic effects on kidney malfunctions in T2DM by inhibiting abnormal fibrosis and improving the ROS defense system.
In summary, oral jaceosidin supplementation improved impaired glucose metabolism and insulin resistance in a mouse model of T2DM. Specifically, jaceosidin contributed to augmenting the phosphorylation of insulin receptor downstream signaling pathways in the liver and skeletal muscles. Furthermore, T2DM-induced nephropathy is ameliorated by therapeutic mechanisms by which jaceosidin intervention attenuates fibrosis and suppresses oxidative stresses in the kidney. The major findings in this study suggest that jaceosidin isolated from Artemisia can be used as a potential drug for treating type 2 diabetes, with few adverse side effects.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the National Research Foundation of Korea (NRF) grant funded by the Korean Government (Grant No. NRF-2013R1A1A2006028).