
Abstract
Fatigue is a common phenomenon usually observed in healthy, as well as in nonhealthy, individuals that affects their performance and quality of life. Efficient supplementation to relieve fatigue is of significant importance. This study was designed to investigate the efficacy of three prescreened natural resources (Cervus elaphus L. [CEL], Angelica gigas Nakai [AGN], and Astragalus membranaceus Bunge [AMB]) against fatigue symptoms induced by heavy exercise. Effects on muscle fatigue and endurance capacity during exercise were investigated in C2C12 myoblasts and exercised mice. A combination of CEL, AGN, and AMB (CEL:AGN:AMB, 1:2:1) treatment in myoblasts reduced intracellular reactive oxygen species levels induced by hydrogen peroxide by ∼20 times (P < .001). The optimal mixture extract combination was determined as CEL:AGN:AMB, 1:2:1 (CAA), which was recombined by applying the extraction yield of individual substance for in vivo study. Compared to the exercise control (EC) group, the serum lactate dehydrogenase level decreased by ∼40% due to CAA administration. The proliferator-activated receptor gamma coactivator 1-alpha protein expression increased significantly (P < .05) after CAA administration compared to that observed in the normal control group. In parallel, CAA treatment significantly (P < .05) enhanced the maximum running time compared to the EC group. Overall, combinatorial administration exhibited greater efficacy compared to each individual treatment, indicating that CAA could be used as an efficient ergogenic and antifatigue supplement.
Introduction
Fatigue can disrupt physiological functionality and life quality that accompanies ailments such as organic illness, immune diseases, and metabolic dysfunction. 1,2 Since the demand for improvement of fatigue is constantly increasing, it is necessary to develop an effective supplement that reduces fatigue.
Natural products have been widely used for energy recovery. Among them, Cervus elaphus L. (CEL), Angelica gigas Nakai (AGN), and Astragalus membranaceus Bunge (AMB) have often been used to ameliorate fatigue. 3,4 CEL (deer antler) is one of the well-known traditional resources in oriental medicine and has been extensively used for improving fatigue symptoms and treating various physical disorders. 5 –7 Its active compounds include polypeptides, polysaccharides, and phospholipids. 7 –9 AGN is known as “female ginseng” and has often been provided as a supplement for various health promoting effects such as antioxidant, anticancer, and anti-inflammatory effects, and its active compounds include decursin, pyranocoumarins, polyacetylenes, and essential oils. 10,11 AMB is also known as a fundamental in oriental medicine, possessing multiple pharmacological properties, such as internal organ protection, antioxidant, hypoglycemic, and antiviral properties. 12,13 In addition, CEL, AGN, and AMB are generally included in the prescribed medicines for ameliorating degenerative diseases. 14 However, CEL can cause a number of side effects like fever, headache, diarrhea, and vomiting if it does not suit one's constitution. Thus, the combinational application with other plant resources is desirable, and this combination of natural substances has been documented in traditional medical literature based on their complementary effects. 14,15 However, the scientific evidence available on its effective combination is insufficient.
To validate the complementary effects of CEL, AGN, and AMB against muscle fatigue and endurance capacity, we aimed to determine the optimal combination ratio in cell culture models, which has been further confirmed by animal studies. To obtain scientific evidence, we analyzed both single and combined forms to investigate their antifatigue properties and potential action mechanisms in acutely heavy exercised mice.
Materials and Methods
Sample preparation
The extracts of CEL, AGN, AMB, and a mixture (indicated as CAA which was recombined by applying the extraction yield of individual substance; CEL:AGN:AMB, 1:2:1) were obtained from Kwangdong Pharmaceutical Co., Ltd. (Seoul, Korea). The materials were extracted in boiling water for 8 h, filtered, evaporated, and spray dried. The dry residue was stored at −80°C.
Total polyphenol content
Total polyphenol content of samples was determined by the Folin–Ciocalteu method 16 with some modifications. Briefly, 500 μL of the samples (1 mg samples dissolved in 1 mL distilled water) were reacted with 1 mL of 2% Na2CO3 solution and 50 μL of Folin–Ciocalteu solution. The absorbance was measured at 720 nm using Perkin Elmer Lambda 35 UV/VIS spectrometer (Perkin Elmer, Waltham, MA, USA), and the results were presented for gallic acid equivalence value.
ABTS free radical scavenging assay
The ABTS (2,2′-azino-bis (3-ethylbenzothiazoline-6-sulfonic acid) free radical scavenging assay was performed using the cation decolorization method 17 with some modifications. In brief, 1 mM AAPH [2,2′-azobis(2-amidinopropane) dihydrochloride] (Sigma-Aldrich, St. Louis, MO, USA) and 2.5 mM ABTS (Sigma-Aldrich) were dissolved in 1 L of phosphate-buffered saline. Following this, 1470 μL of ABTS radical solution was mixed with 30 μL of samples and reacted for 10 min in the dark. The absorbance was measured at 734 nm using a Perkin Elmer Lambda 35 UV/VIS spectrometer (Perkin Elmer), and the results were presented for ascorbic acid equivalence value.
Cell culture and cytotoxicity test
The skeletal muscle cell line C2C12 myoblasts (ATCC, Manassas, VA, USA) were maintained in high glucose Dulbecco's modified Eagle's medium supplemented with 10% fetal bovine serum and 1% penicillin solution. Briefly, C2C12 myoblasts were seeded in 96-well plates (1 × 104 cell/well) and cultured at 37°C in 5% CO2 atmosphere for 24 h. The media was then changed to enable 0.01–1 mg/mL concentration of CEL, AGN, and AMB samples and maintained for additional 24 h. After sample treatment, 10 μL of MTT (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide, a tetrazole) (Sigma-Aldrich) solution (5 mg/mL) was added to each well and maintained for 3 h to evaluate the cytotoxicity of C2C12 myoblasts. The formazan precipitates were dissolved in 100 μL of dimethyl sulfoxide (DMSO) (Samchun Pure Chemical Co., Ltd., Seoul, Korea), and the absorbance was measured at 595 nm using Bio-Rad 680 microplate reader (Bio-Rad, Hercules, CA, USA). 18
Cytoprotective effects
The C2C12 myoblasts were seeded in 96-well plates (1 × 104 cell/well) and cultured for 24 h. Fresh hydrogen peroxide (H2O2) (Sigma-Aldrich) solution (final concentration, 0.5 mM) and the CEL, AGN, and AMB samples (final concentration, 1 mg/mL) were cotreated for additional 24 h. Ten microliters of 5 mg/mL MTT solution were added to each well and maintained for 3 h to evaluate the cytoprotective effects of CEL, AGN, and AMB extracts. The formazan precipitates were dissolved in 100 μL of DMSO, and the absorbance was measured at 595 nm using Bio-Rad 680 microplate reader (Bio-Rad). 18
Intracellular reactive oxygen species assay
A fluorescent probe (2,7-dichlorofluorescin diacetate; DCFH-DA) was used to analyze the formation of intracellular reactive oxygen species (ROS) in the C2C12 myoblasts. Briefly, C2C12 myoblasts were seeded into 96-well plates (1 × 104 cell/well) and cultured at 37°C in a 5% CO2 atmosphere for 24 h. The media was changed to enable 0.01–0.02 mg/mL concentration of CEL, AGN, AMB and combined extract pretreated for 1 h. H2O2 solution (final concentration, 0.5 mM) was then treated for additional 2 h to generate intracellular ROS. Finally, the cells were loaded with the probe (final concentration, 10 mM) and incubated for 30 min. After incubation, the cells were immediately placed in a spectrofluorometer to determine ROS generation at the excitation and emission wavelengths of 535 and 485 nm using Perkin Elmer 1420 multi label counter (Perkin Elmer), respectively. 19
Animal model
The 60 male ICR (6 weeks old) mice were procured from Raon Bio, Inc. (Yong-in, Korea). The mice were allowed to adapt to their surroundings for 1 week before starting the experiments with controlled lighting (12-h light/12-h dark cycle), humidity (50–60%), and temperature (20–23°C). Experimental procedures and animal care were approved by the Animal Care Committee of Korea University (KUIACUC-2019-0052).
Exercise training and endurance capacity
After 2 weeks of acclimation on a treadmill (MiraeST Corp., Daejeon, Korea) at 0% inclination, the mice were divided randomly into six groups (n = 10) as follows: A: normal control (NC) group (saline, no exercise); B: exercise control (EC) group (saline, exercise); C: CEL-treated group (concentration: 150 mg/kg, exercise); D: AGN-treated group (concentration: 150 mg/kg, exercise); E: AMB-treated group (concentration: 150 mg/kg, exercise); and F: CAA-treated group (concentration: 150 mg/kg, exercise). Physical fatigue was evaluated by treadmill exercise. All mice were trained on the treadmill thrice a week for 3 weeks—during the first week, 20 m/min for 5 min, 25 m/min for 5 min, and 30 m/min for 5 min; during the second week, 20 m/min for 5 min, 25 m/min for 10 min, and 30 m/min for 5 min; and during the third week, 20 m/min for 5 min, 25 m/min for 15 min, and 30 m/min for 15 min. 20 After 21 days of sample treatment, endurance capacity was determined. At 30 min after the final treatment, the mice were placed individually on treadmill lanes. The speed was then increased from 20 to 40 m/min every 3 min, and the time was measured until the point where the mouse failed to follow the speed increase on three consecutive occasions and lag occurred. At the point of exhaustion, the mouse was removed from the treadmill. The total running time until exhaustion was recorded and used as the index of endurance capacity. 20
Blood serum parameters and tissue collection
The serum levels of glucose, lactate dehydrogenase (LDH), blood urea nitrogen (BUN), aspartate aminotransferase (AST), and alanine transaminase (ALT) were determined immediately following exercise. After 30 min of the last treatment, the five mice remaining from each group underwent a 40-min treadmill exercise at 30 m/min speed. After the treadmill exercise, the mice were euthanized with avertin (200 mg/kg dosage), and the blood samples were collected immediately from the heart. The serum was separated by centrifugation at 13,000 g for 20 min at 4°C. The levels of serum glucose, LDH, BUN, AST, and ALT were determined using an AU480 Chemistry Analyzer (Beckman Coulter, Brea, CA, USA). Liver, gastrocnemius muscle, and epididymal fat tissues were rapidly removed, washed in cold saline, and stored at −80°C before tissue analysis.
Gastrocnemius muscle protein expression
The gastrocnemius muscle tissue was lysed in lysis buffer with proteinase inhibitor. The total protein concentration was determined using Bradford solution. The protein (15 μg) was separated by 10% and 12% sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to a nitrocellulose membrane. The membranes were blocked with 3% bovine serum albumin in tris-buffered saline-Tween 20 solution for 1 h at room temperature. The membranes were then incubated with the primary antibodies (NF-κB [nuclear factor kappa light chain enhancer of activated B cells; sc-372] and IL-6 [interleukin 6; sc-1265-R] were purchased from Santa Cruz Biotechnology [Dallas, TX, USA]; AMPK [AMP-activated protein kinase; CST2532s], P-AMPK [phospho AMP-activated protein kinase; CST2531s], and NRF1 [nuclear respiratory factor 1; CST12381] were purchased from Cell Signaling Technology [Danvers, MA, USA]; PGC-1α [proliferator-activated receptor gamma coactivator 1-alpha; ab191838] and TFAM [transcription factor A of the mitochondrion; ab131607] were purchased from ABCAM [Cambridge, United Kingdom]) overnight at 4°C, followed by incubation with the corresponding horseradish peroxidase-conjugated secondary antibody (Sigma-Aldrich) for 1 h at room temperature. The blots were developed using enhanced chemiluminescence reagents (Bio-Rad), captured by an ImageQuant LAS-4000 imager (General Electric, Pittsburgh, PA, USA), and quantified using NIH ImageJ software (National Institutes of Health, Bethesda, MD, USA). 21
Statistical analyses
Statistical analyses were performed using the IBM SPSS v 25.0 software (IBM Corp., Armonk, NY, USA). Data are represented as the mean ± standard error of mean from three independent experiments, unless stated otherwise. Differences between groups were analyzed by Student's t-test and one-way analysis of variance followed by Duncan's multiple range test, and P < .05 was considered to indicate a significant difference.
Results
Antioxidant activity
The abundance of total polyphenol content and ABTS free radical scavenging activity of CEL, AGN, and AMB are presented in Figure 1. Among these, total polyphenol content of AGN was more than 1.5 times higher compared with CEL and AMB. In parallel, AGN exhibited more than five times higher free radical scavenging ability than both CEL and AMB. Therefore, AGN was anticipated to have greater antioxidant capacity than CEL and AMB.
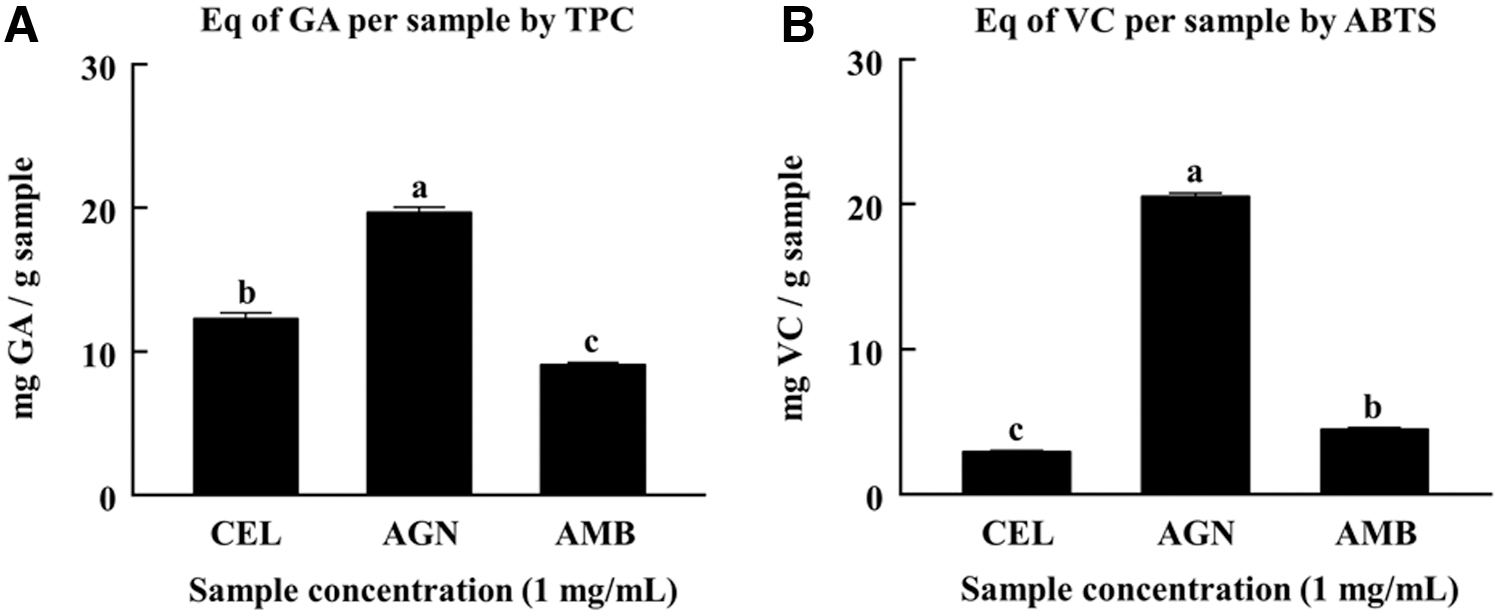
The abundance of
Cytoprotective effects against ROS-induced myoblast damage
As presented in Figure 2, the extract treatment on C2C12 myoblasts did not elicit visible cytotoxicity up to 1 mg/mL concentration. As estimated in the MTT assay (Fig. 3), the H2O2 concentration up to 1 mM showed significant cytotoxicity in a dose dependent manner when added to C2C12 myoblasts. Therefore, we chose 0.5 mM concentration of H2O2 which could induce about 30% cytotoxicity in myoblasts to assess the cytoprotective effects of test samples. As a result, the AMB treatment significantly restored the cell viability level by 10% compared to the control (H2O2 treated). However, CEL and AMB treatment had little effect compared to the control.

Cytotoxicity of
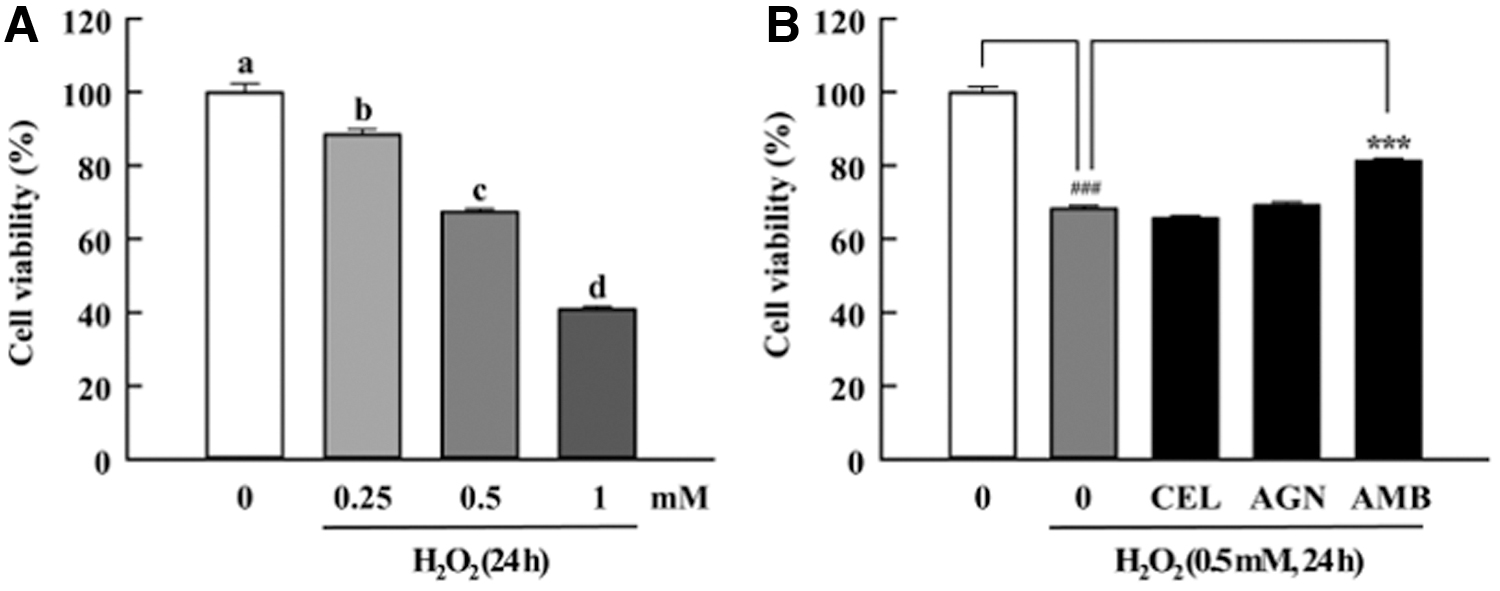
Cytoprotective effects against ROS-induced myoblast damage. CEL, AGN, and AMB. Data are expressed as mean ± SEM (n = 10). Different letters above the bars indicate significant differences at P < .05. ### P < .001 and ***P < .001 indicate statistical significance. ROS, reactive oxygen species.
Intracellular ROS modulation effects
To demonstrate the intracellular ROS modulation effects of the natural resources, we selected H2O2 to induce the oxidative stress in myoblasts that may occur during prolonged exercise. The description of each sample treatment group is displayed in Table 1. As shown in Figure 4, the H2O2 treatment induced cellular ROS levels up to 20 times higher than that in normal control. However, CEL and AMB treatment exerted little effect compared to the H2O2-treated group. In contrast, AGN treatment at low and high doses demonstrated significant intracellular ROS scavenging effect compared to H2O2, CEL, and AMB treatment. Notably, all combined extract-treated groups demonstrated substantial ROS removal efficacy, similar to that in the AGN-treated group administered with a low dose of AGN. In particular, group I exhibited the greatest antioxidant capacity compared to other combination groups. Accordingly, we determined the optimal combination ratio (CEL:AGN:AMB, 1:2:1) to make an extract blend (CAA) which was recombined by applying the extraction yield of individual substance for the in vivo study. These fundamental data were used to design the animal studies to evaluate the potential antifatigue activity.

Intracellular ROS modulation effects. CEL, AGN, and AMB. Data are expressed as the mean ± SEM (n = 10). Different letters above the bars indicate significant differences at P < .05.
Sample Treatment Groups
AGN, Angelica gigas Nakai; AMB, Astragalus membranaceus Bunge; CEL, Cervus elaphus L.
Effect of administration on physiological parameters
To assess the possible toxicity of sample treatment in mice, body weight and food consumption were monitored throughout the duration of the study. As presented in Figure 5, none of the extract treatments induced apparent toxicity or side effects, marked by sustained body weight increase and food intake during all periods. Moreover, there was no significant difference in liver weight of mice in any of the groups (Table 2). Mice in the EC group had the highest gastrocnemius muscle weight compared to those from the NC group, while no significant difference was observed between EC and sample treatment groups. However, there was a significant reduction in epididymal fat weight in all exercise-trained mice compared to NC mice.
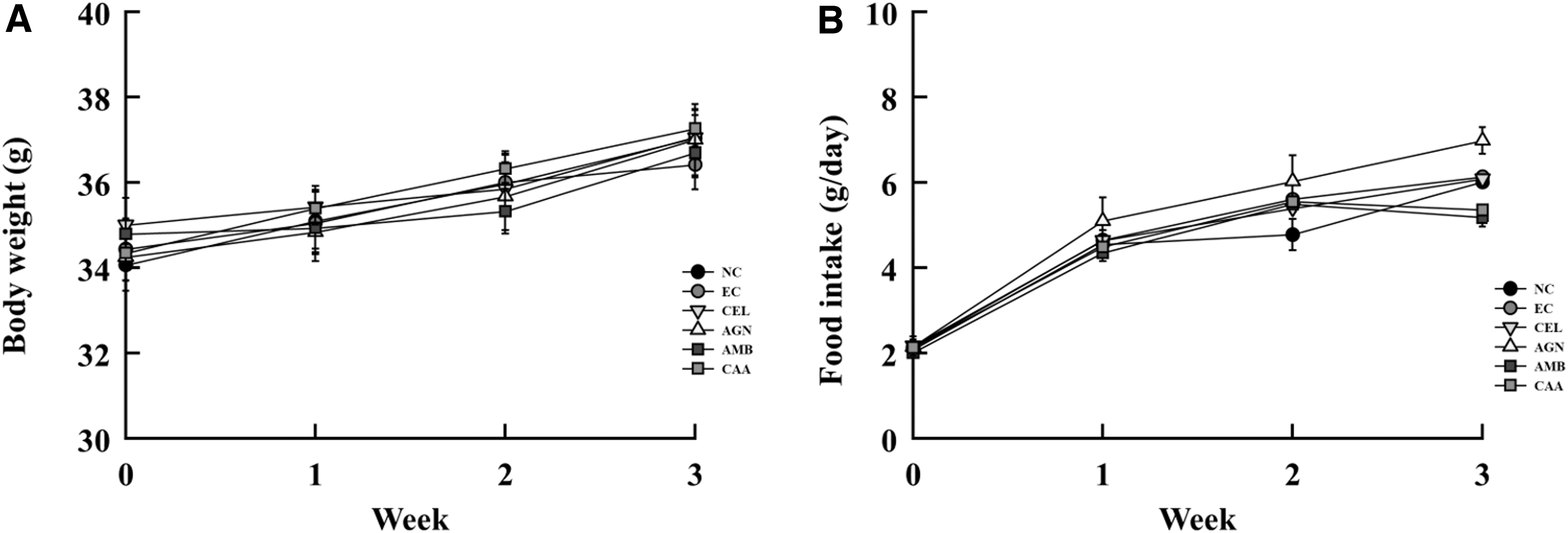
Effects of administration on
Effects of Administration on Organ Weight Change
Data are presented as mean ± SEM (N = 10).
BW, body weight; EC, exercise control; NC, normal control; CAA, Cervus elaphus L., Angelica gigas Nakai, and Astragalus membranaceus Bunge complex-treated group.
P < .05, ## P < .01, compared to the NC group.
SEM, standard error of the mean.
Antifatigue effect
Endurance capacity of exercise-trained mice was determined by the maximum running time recorded on treadmill. As presented in Figure 6, the time to exhaustion running time tended to increase in the sample treated groups compared to the EC group. Specifically, the maximum running time increased significantly by CAA administration, compared to that observed in control mice. This indicated that the combination of CEL, AGN, and AMB exerted synergistic or complementary effects against fatigue induced by treadmill exercise compared to single treatments.

Antifatigue effect. Data are presented as mean ± SEM (n = 5). *P < .05 indicates statistical significance.
Effect of administration on serum parameters
The serum levels of glucose, LDH, BUN, AST, and ALT have been used to investigate muscle injury during exhaustive exercise. In this study, there was no significant difference observed in serum glucose levels among the groups (Table 3). The LDH levels in CAA-treated mice were marginally lower than that in the EC group (P < .07). In our study, there were no significant differences in serum BUN levels among the groups. Furthermore, the CEL group exhibited higher AST and ALT levels compared to the EC group. Conversely, CAA administration maintained lower serum ALT levels compared to the EC group (P < .01).
Effects of Administration on Serum Parameters
Data are presented as mean ± SEM (N = 5).
P < .05, compared to the NC group; ** P < .01, compared to the EC group.
ALT, alanine transaminase; AST, aspartate aminotransferase; BUN, blood urea nitrogen; IU, international units; LDH, lactate dehydrogenase.
Protective effect on cytokine expression after treadmill exercise
As depicted in Figure 7, the EC group had a significantly (P < .05) higher NF-κB expression than the NC group, indicating that intensive treadmill exercise promoted muscle damage through NF-κB activation. Conversely, none of the individual sample treatment groups displayed significant differences from the EC group. Furthermore, the CAA group had a lower NF-κB expression compared to CEL, AGN, and AMB groups, while there was no significant difference compared to the EC group. In terms of IL-6 protein expression, none of the exercised groups exhibited significant difference with the EC group.

Protective effect on cytokine
The protein expression associated with muscle mitochondrial biogenesis was analyzed. In the present study, our results indicated that all sample treatments tended to increase the P-AMPK and AMPK expression compared to the expression in NC mice, while there was no significant difference in P-AMPK/AMPK ratio among the groups. There was a significantly higher PGC1-α expression in CAA group compared to the NC group. Furthermore, AGN and CAA treatment tended to enhance the NRF1 expression. These data indicated that AGN might have acted as the main active source that ameliorated muscle damage. There was no significant difference in TFAM expression among the groups. Overall, stimulation of muscle protein expression in exercised mice was more pronounced with CAA treatment.
Discussion
This study confirmed that the optimized combinatorial treatment reduced the cellular ROS level in damaged myoblasts. Furthermore, CAA administration enhanced the running endurance capacity of mice and ameliorated the muscle oxidative properties through AMPK-mediated signaling pathway. It is reported that the beneficial effects of oriental medicine on antifatigue effects are mediated by various active components. 3,4 Polyphenols are the primary active compounds present in most herbal medicines that elicit multiple physiological properties, such as antioxidant, anticancer, and anti-inflammatory properties. 22,23 Many researchers have investigated the antioxidant activity of CEL (polypeptides, polysaccharides, and phospholipids 7 –9 ), AGN (decursin, decursinol angelate, and essential oils 10,11 ), and AMB (astragaloside IV and polysaccharides 12,13 ) due to their bioactive components. Our results showed that AGN exhibited significantly higher polyphenol content and free radical scavenging ability than CEL and AMB. Taken together, AGN was anticipated to have greater antioxidant capacity than CEL and AMB. Cellular oxidative stress was induced by H2O2 treatment that could raise the cellular ROS levels to induce myoblast damage and necrosis. 19 In view of this, AMB exerted higher cytoprotective effect than AGN against oxidative stress-induced myoblast damage. These results indicated that AGN was efficient in removing ROS, and AMB was potent in preventing cell damage. Comprehensively, AGN and AMB could be expected to exhibit higher antioxidant capacity than CEL at the cellular level.
Excessive fatigue condition in this study was simulated by H2O2 treatment in myoblasts. To investigate the intracellular ROS modulation effects of sample treatment, DCFH-DA probe was used for measuring the total cellular ROS such as H2O2, hydroxyl radicals (OH•), and hydroperoxides (ROOH). According to Figure 4, H2O2 treatment significantly elevated the intracellular ROS level in myoblasts compared to normal control (H2O2 untreated). In contrast, all combined groups showed significant reduction in intracellular ROS level compared to control group. Importantly, AGN treatment significantly reduced the oxidative damage by dose dependent manner. Therefore, we prospected that AGN could be considered as major source regulating oxidative stress.
Swimming pool or treadmill exercises are the main experimental systems used to investigate the fatigue behavior of mice. The present study used treadmill running to confirm the effects of combinatorial administration on endurance capacity of mice because of the large variation in data from swimming pool model. In this study, only the CAA group significantly increased the maximum running time compared to control group. Consequently, it was demonstrated that CAA administration exerted synergistic or complementary effects against fatigue induced by treadmill exercise compared to individual treatment.
Fatigue mediated serum parameters have been used to consider muscle injury during exhaustive exercise. Maintenance of serum glucose is crucial in extending exercise endurance. 24 Hypoglycemia induced by prolonged exercise can lead to the deterioration of overall brain function, reducing the ability to continue exercise. 25 In this study, there was no significant difference observed in serum glucose levels among the test groups (Table 3). LDH regulates lactate formation, and conversion from pyruvate occurs during anaerobic condition in muscle, which is released in the blood. 26 Increased serum LDH level can be considered an early indicator of cellular necrosis and tissue damage. 27 The LDH levels in CAA-treated mice were marginally lower than that in the EC group (P < .07). BUN is a well-known byproduct of energy metabolism, which can estimate the degree of kidney dysfunction and fatigue symptoms. In fact, there is a positive correlation between BUN and exercise tolerance. 28 In our study, there was no significant difference in BUN level between each group. Moreover, the elevation of AST and ALT can predict liver damage and is also recognized as a response to exhaustive physical exercise. 29,30 Our data showed that increased oxidative hepatic damage induced by exhaustive exercise was efficiently attenuated by CAA administration.
As an indicative marker of oxidative damage to muscle, the occurrence of inflammatory protein expression was evaluated. Highly intensive exercise can induce muscle damage by enhancing inflammation that is directly associated with expression of cytokines such as NF-κB and IL-6. 31 Accordingly, the EC group showed significant higher inflammatory protein expression than NC group. It was indicated that intensive treadmill exercise promoted muscle damage through NF-κB activation. However, there were no significant differences in NF-κB and IL-6 expression among sample treatment groups. Under different circumstances, treadmill exercise can promote muscle mitochondrial biogenesis to delay skeletal muscle fatigue and enhance endurance exercise capacity. 32 Among them, AMPK is considered an important initiator of muscle mitochondrial biogenesis using PGC1-α, a critical regulator for NRF1 activation which can further upregulate the expression of TFAM that activates mitochondrial DNA duplication and gene transcription. 32,33 We did not observe any significant change in above markers among the exercised groups except for PGC1-α. Meanwhile, AGN and CAA treatment tended to increase the P-AMPK, PGC1-α, and NRF1 expression. These data indicated that AGN might have acted as the main active source that ameliorated muscle damage. Thus, CAA treatment may be able to enhance exercise-mediated mitochondrial biogenesis in skeletal muscle.
In summary, this study demonstrated that combinatorial administration exhibited a cytoprotective effect by modulating intracellular ROS levels in damaged myoblasts. CAA treatment exerted an antifatigue effect by enhancing the endurance capacity in exercised mice, which was insignificant in individual administration. We also observed the reduced serum levels of LDH and ALT with CAA administration in exercised mice, indicating that the restoration was enabled during exercise. Moreover, CAA administration increased the PGC1-α expression, the key regulator involved in mitochondrial biogenesis in skeletal muscle of exercise-trained mice. In conclusion, this study provided a better understanding of the combined application of CAA as a valuable ergogenic and antifatigue supplement.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by Kwang-Dong Pharmaceutical Co., Ltd., and Korea University Research Grant.