
Abstract
The effects of a mixture of fisetin on cytokine-mediated pulmonary damages have not been studied, despite its known antiviral, neuroprotective, and anti-inflammatory activities. Using lipopolysaccharide (LPS)-activated human pulmonary artery endothelial cells (HPAECs), we determined the effects of fisetin on the induction of heme oxygenase-1 (HO-1), inducible nitric oxide synthase (iNOS), and cyclooxygenase-2 (COX-2). In the lung tissue of LPS-treated mice, fisetin was also evaluated for its effect on the regulation of iNOS and tumor necrosis factor (TNF)-α. In LPS-activated HPAECs, fisetin increased nuclear factor erythrocyte 2-related factor 2-antioxidant response element (Nrf2-ARE) reporter activity through the nuclear translocation of Nrf2, and the expression of HO-1, and decreased IL-1β and iNOS/NO production. In particular, the suppression of iNOS/NO expression by the administration of fisetin was dependent on HO-1. Current findings indicate that the anti-inflammatory activity of fisetin was due to its HO-1 dependent downregulation of p-STAT-1 and nuclear factor kappa B (NF-κB) and the resultant inhibition of iNOS, and also suggest TNF-α as a potential target for HO-1. We propose that administration of fisetin may be a novel approach, ideal for the treatment of inflammatory pulmonary disease.
Introduction
In inflammatory processes, nuclear factor kappa B (NF-κB) upregulates the synthesis of various proinflammatory mediators. Contrary to NF-κB, heme oxygenase-1 (HO-1) suppresses the generation of proinflammatory mediators such as interleukin (IL)-1β, IL-6, and tumor necrosis factor (TNF)-α in reaction to oxidation and inflammatory injury. 1 HO-1 has been shown to enhance survival rates in a mouse sepsis model, in which sepsis was induced by the cecal ligation and puncture method, 1,2 and has exhibited favorable effects in the treatment of various vascular inflammatory disorders. 1
Nuclear factor erythrocyte 2-related factor 2 (Nrf2) has been shown to regulate the expression of HO-1. In response to oxidative and also other stresses, Nrf2 controls transcription of phase II antioxidant enzymes by binding to the antioxidant response elements (AREs) on their promoters. 3 For this reason, the Nrf2-ARE signaling is considered as a crucial target for the treatment of inflammation-related diseases. 3
Recently, it has been found that treatments with flavonoids from various herbal extracts can alleviate inflammatory responses. 4,5 Flavonoids are polyphenolic compounds with low molecular weight from vegetables, fruits, and plants, with potent biological effects. The biological effects include antiplatelet, antithrombotic, antioxidant, anticarcinogenic, and cytoprotective activities, 6,7 and have been suggested to regulate immune responses. 8 Fisetin is a polyphenol, from the flavonoid group, commonly found in plants belonging to the genus Rhus (R. cotinus) or Acacia (A. berlandieri, A. greggii), and has been known to have antiviral, anti-inflammatory, 9,10 and also neuroprotective functions. Fisetin suppresses amyloid beta protein aggregation, which is proposed to be responsible for progressive neuronal loss in Alzheimer's disease patients. 11 Based on our understanding of the above-mentioned effects of fisetin, we hypothesized that the administration of fisetin may show beneficial effects against cytokine-induced pulmonary vascular inflammatory responses and determined the regulatory mechanisms of fisetin on the suppression of inflammatory mediators.
Materials and Methods
Cells and reagents
We purchased primary human pulmonary artery endothelial cells (HPAECs) from Cambrex Bio Science (Charles City, IA, USA) and maintained them according to the manuals provided. 12 –14 HPAEC culture passages 3–5 were employed in our study. Fisetin, dimethyl sulfoxide (DMSO), lipopolysaccharide (LPS), and antibiotics (streptomycin and penicillin G) were purchased from Sigma Chemical Co. (St. Louis, MO, USA). siRNA for human HO-1 gene knockdown studies was purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA).
Lung injury mouse model induced by LPS and bronchoalveolar lavage fluid analysis
Our animal protocol was approved by the Kyungpook National University Animal Care Committee (IRB No. KNU 2017-102). Male C57BL/6 mice (body weight: 27 g and 6 or 7 weeks old) were obtained from Orient Bio Co. (Sungnam, Republic of Korea) and used after a standard acclimatization period of 12 days. 15,16 LPS (15 mg/kg), injected with a 28-gauge needle intraperitoneally, was used to induce lung injury. Six hours after each injection of LPS, fisetin (10–200 μg/kg) was intravenously administered in mice. We obtained bronchoalveolar lavage fluid (BALF) with an intratracheal injection of cold phosphate-buffered saline (PBS) (0.7 mL) and gentle aspiration thrice. The BALF was pelleted down, and then the supernatant was maintained at −80°C for further analysis.
Enzyme-linked immunosorbent assays for p-STAT-1 (total or phosphor), HO-1, iNOS, PGE2, IL-1β, and TNF-α
We used enzyme-linked immunosorbent assay (ELISA) kits (R&D Systems, Minneapolis, MN, USA) to determine the nuclear expressions of the total and phosphorylated STAT1 (Abcam, UK) in HPAECs and the levels of HO-1, inducible nitric oxide synthase (iNOS), PGE2, IL-1β, and TNF-α in the supernatant of cell culture medium. We independently performed all ELISA measurements in triplicate.
Nitrite production measurement
The relative degrees of NO produced were calculated from the detection of a stable end product of NO metabolism, nitrite (NO2–), in the cell culture medium. The 1:1 mixtures of the culture supernatant and the Griess reagent were incubated for 15 min at RT, and the reactions were spectrophotometrically analyzed (λ = 540 nm). The level of nitrite was calculated using known concentrations of sodium nitrite, which was used as the standard.
Cell viability assay
HPAECs, grown in 96-well plates with a cellular density of 5 × 103 cells/well, were incubated with fisetin for 2 days. One hundred microliters of 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (1 mg/mL) was added to each well, and incubated for 4 h, and 150 μL of DMSO per each well was added to solubilize the formazan salt formed. The amount of solubilized formazan salt was spectrophotometrically analyzed (λ = 540 nm).
Isolation of cytoplasmic and nuclear fractions and immunoblotting
HPAECs were rapidly collected by centrifugation, and subsequently both nuclear and cytoplasmic fractions were isolated on ice according to an established protocol. 12 For immunoblotting, we used anti-Nrf2, anticyclooxygenase-2 (COX-2), anti-iNOS, antilamin B, and β-actin antibodies (Santa Cruz). Detections of β-actin and lamin B were used to represent loading controls for the cytoplasmic and nuclear extracts, respectively.
Quantitative real-time polymerase chain reaction analysis
Listed here are the primers used: forward primer for COX-2 5′-CCC CAT TAG CAG CCA GTT-3′, reverse primer for COX-2 5′-CAT TCC CCA CGG TTT TGA-3′; forward primer for iNOS 5′-GTT CTC AGC CCA ACA ATA CAA GA-3′, reverse primer for iNOS 5′-GTG GAC GGG TCG ATG TCA C-3′; and forward primer for β-actin 5′-TCGTGCGTGACATCAAAGA-3′, reverse primer for β-actin 5′-CAT ACC CAA GAA GGA AGG CT-3′. We normalized the levels of iNOS and COX-2 to that of β-actin.
Transfection of reporter plasmids and siRNA
NF-κB luciferase reporter plasmid, ARE luciferase reporter plasmid, HO-1 siRNA, and control nonsense siRNA were transfected into cells with SuperFect Transfection Reagent (Qiagen, Valencia, CA, USA).
Reporter assay
After each indicated treatment, PBS-washed cells were lysed in a supplied lysis buffer in a dual luciferase kit (Promega, Madison, WI, USA), and assayed with a luminometer. All measurements were conducted three times, and the results are shown as the ratio between firefly and Renilla luciferase activity.
Histopathological examination
Male mice (n = 5) were first injected with LPS, and 6 h later, intravenously administered with fisetin (200 μg/kg). The mice were then sacrificed, and lung tissues were harvested. The phenotypic changes in the lungs were microscopically analyzed from hematoxylin and eosin-stained tissue slides. 17
Statistical analysis
Results are presented as the mean value ± standard deviation from three independent experiments. Post hoc testing of the differences was additionally performed using Tukey's tests, after analysis of variance (ANOVA) showed a significant difference between different groups. A P value <.05 is statistically significant.
Results
Fisetin regulation of iNOS and COX-2 expression in LPS-activated HPAECs
To determine the anti-inflammatory effect of fisetin, we characterized the expression levels of iNOS and COX-2 as representative proinflammatory genes. HPAECs were stimulated with LPS for 6 h, and then the cells were treated with indicated doses of fisetin for an additional 6 h. Our quantitative real-time polymerase chain reaction, ELISA, and immunoblot analyses revealed that the transcriptional and translational levels of iNOS and COX-2 were reduced with the treatment of fisetin in a concentration-dependent manner (Fig. 1A–D). Each level of corresponding products of iNOS and COX-2, NO and PGE2, was also reduced with fisetin treatment (Fig. 1E, F), indicating that fisetin primarily suppresses LPS-induced production of NO and PGE2 through the fisetin-mediated downregulation of iNOS and COX-2, respectively.
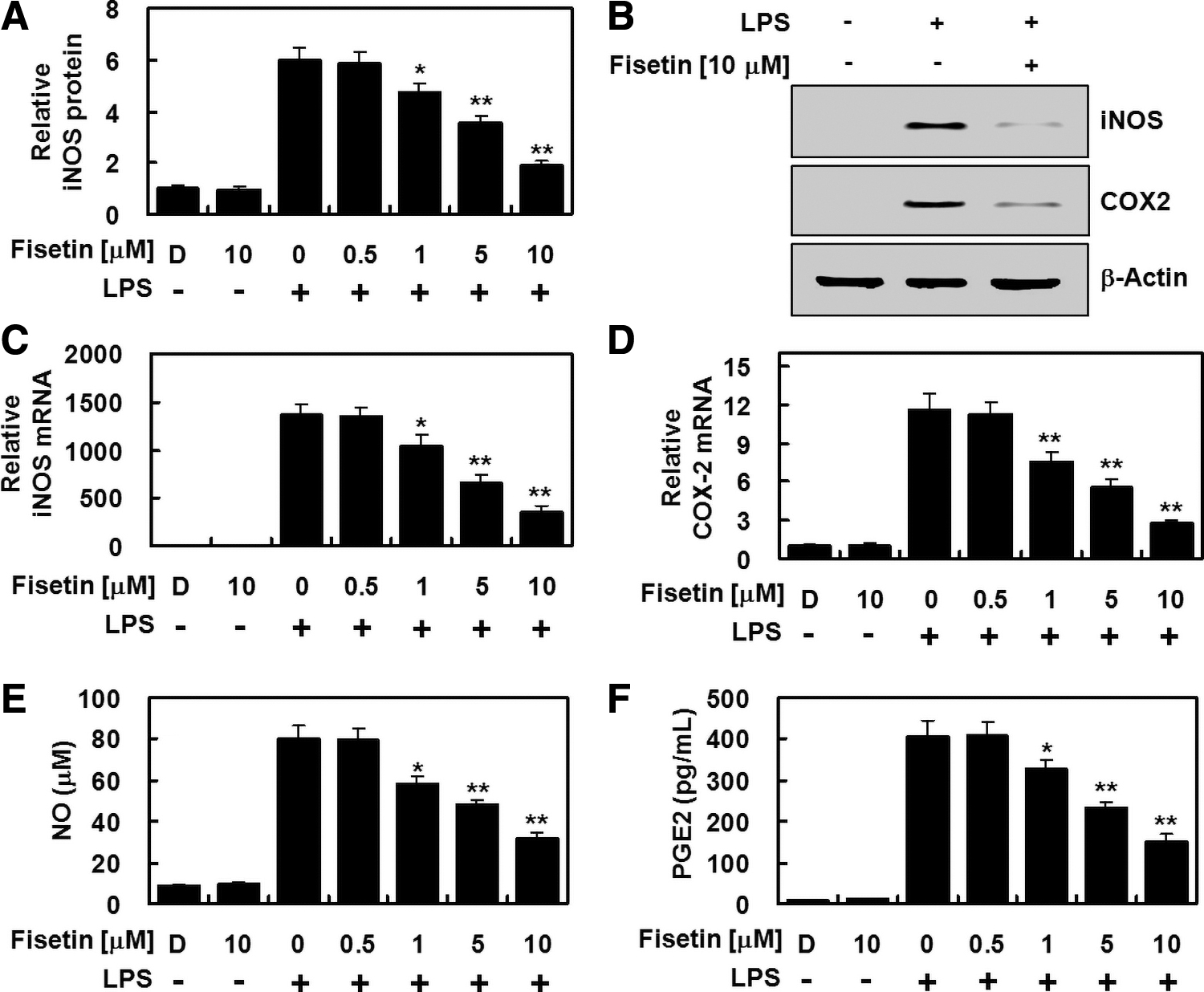
Fisetin suppressed expression of iNOS and COX-2 protein, induced in LPS-activated HPAECs. HPAECs were stimulated with LPS (1 μg/mL) for 6 h, treated with the indicated different concentrations of fisetin for an additional 6 h, and the levels of
Fisetin regulation of NF-κB activity, the STAT-1 phosphorylation, and the HO-1 expression
Since NF-κB is an important regulator of inflammatory gene expression, we investigated fisetin inhibition of NF-κB. NF-κB luciferase activity was suppressed with fisetin treatment in a concentration-dependent manner (Fig. 2A). Because it is well established that, in an LPS-activated state, the activation of the janus kinase/signal transducers and activators of transcription (JAK/STAT) signal is essential for the upregulation of iNOS and COX-2, 18,19 we determined whether the STAT-1 phosphorylation, which represents the activation of JAK/STAT signaling, was suppressed with fisetin treatment in LPS-activated HPAECs, and this was found to be the case (Fig. 2B). We also found that expression of anti-inflammatory HO-1 was significantly upregulated with fisetin treatment (Fig. 2C). Moreover, the cytotoxicity possible from administration of fisetin was determined, and data showed that fisetin did not affect the cellular viability up to 30 μM and that fisetin suppressed inflammatory responses in a physiological, nonstressful manner in our study (Fig. 2D).

Fisetin suppressed NF-κB luciferase reporter activity and STAT-1 phosphorylation induced by LPS stimulation, but upregulated expression of HO-1 protein. HPAECs were stimulated with LPS (1 μg/mL) for 6 h, and then treated with the indicated different concentrations of fisetin for an additional 6 h.
Nrf2-ARE signaling and HO-1 are necessary for fisetin-mediated anti-inflammatory response
Nrf2 is an important transcription factor in the transcription of antioxidant proteins, including HO-1. We investigated whether fisetin-mediated activation of Nrf2-ARE pathway was responsible for the increase in HO-1 expression shown in Figure 2C. We showed that fisetin induced the nuclear translocation of Nrf2 (Fig. 3A) and also upregulated Nrf2-ARE reporter activity (Fig. 3B). Next, RNAi gene knockdown of HO-1 was conducted to confirm that the fisetin inhibition of iNOS expression was mediated by the induction of HO-1. When HO-1 expression was suppressed, the downregulation of iNOS protein and its product, NO, by fisetin treatment was abolished (Fig. 3C, D). This indicates that fisetin induced the expression of HO-1, which was consequently responsible, at least in part, for the downregulation of iNOS expression. Fisetin treatment inhibited production of proinflammatory IL-1β in LPS-treated HPAECs (Fig. 3E), again validating the anti-inflammatory effect of fisetin.
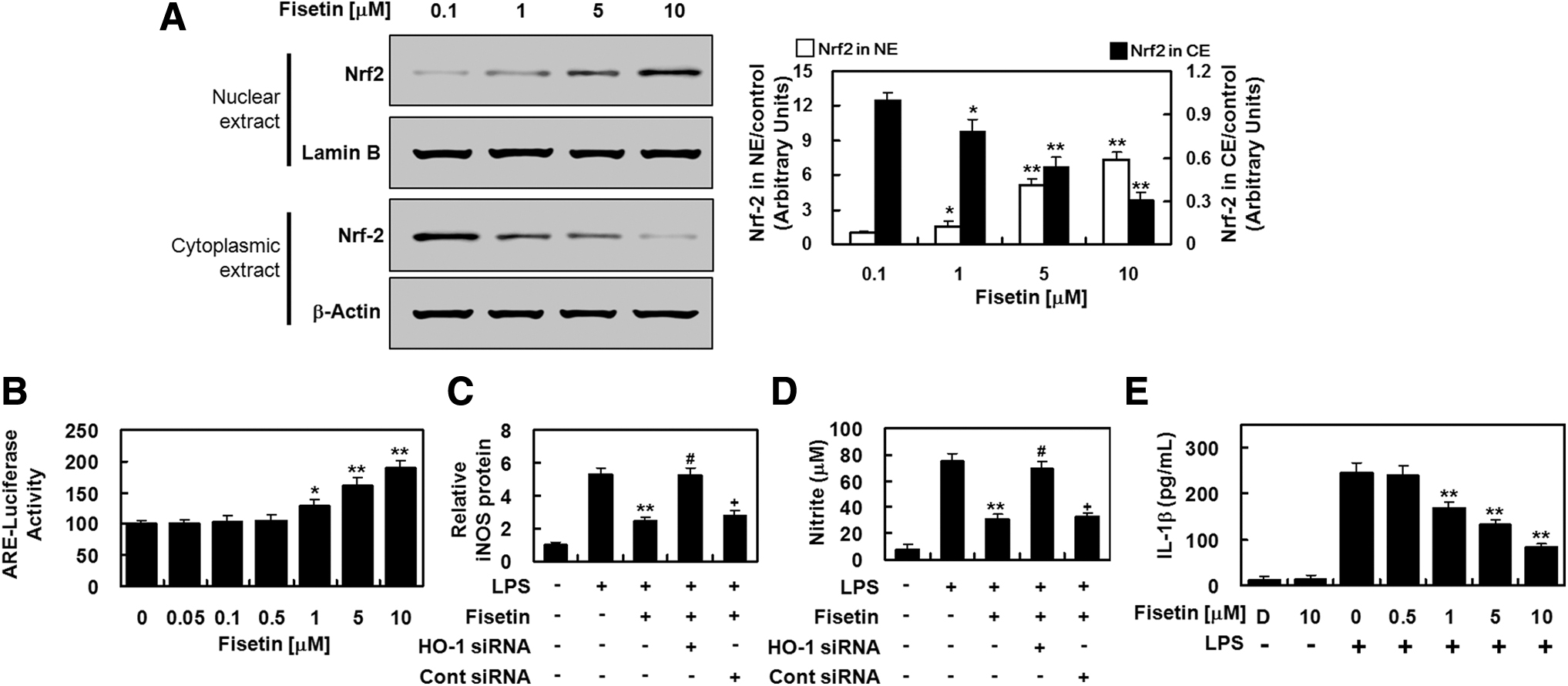
Fisetin induced the nuclear translocation of Nrf2 and anti-inflammatory responses in HPAECs.
Fisetin regulation of TNF-α and iNOS in mouse lung tissue
Finally, we questioned if the administration of fisetin could also produce an anti-inflammatory effect in vivo. Fisetin treatment significantly reduced levels of iNOS (Fig. 4A) and TNF-α (Fig. 4B) in BALF from LPS-treated mice, indicating that fisetin effectively suppresses inflammatory responses in vivo. Fisetin also considerably ameliorated the pulmonary injury induced by LPS (Fig. 4C). Since circulating blood volume in a mouse is estimated to be 72 mL/kg, 20,21 we calculated the average blood volume in each mouse as 2 mL. Thus, we assumed that an injection of 10, 20, 100, or 200 μg/kg of fisetin yielded an estimated concentration of fisetin in the peripheral blood ∼0.5, 1, 5, or 10 μM, respectively.
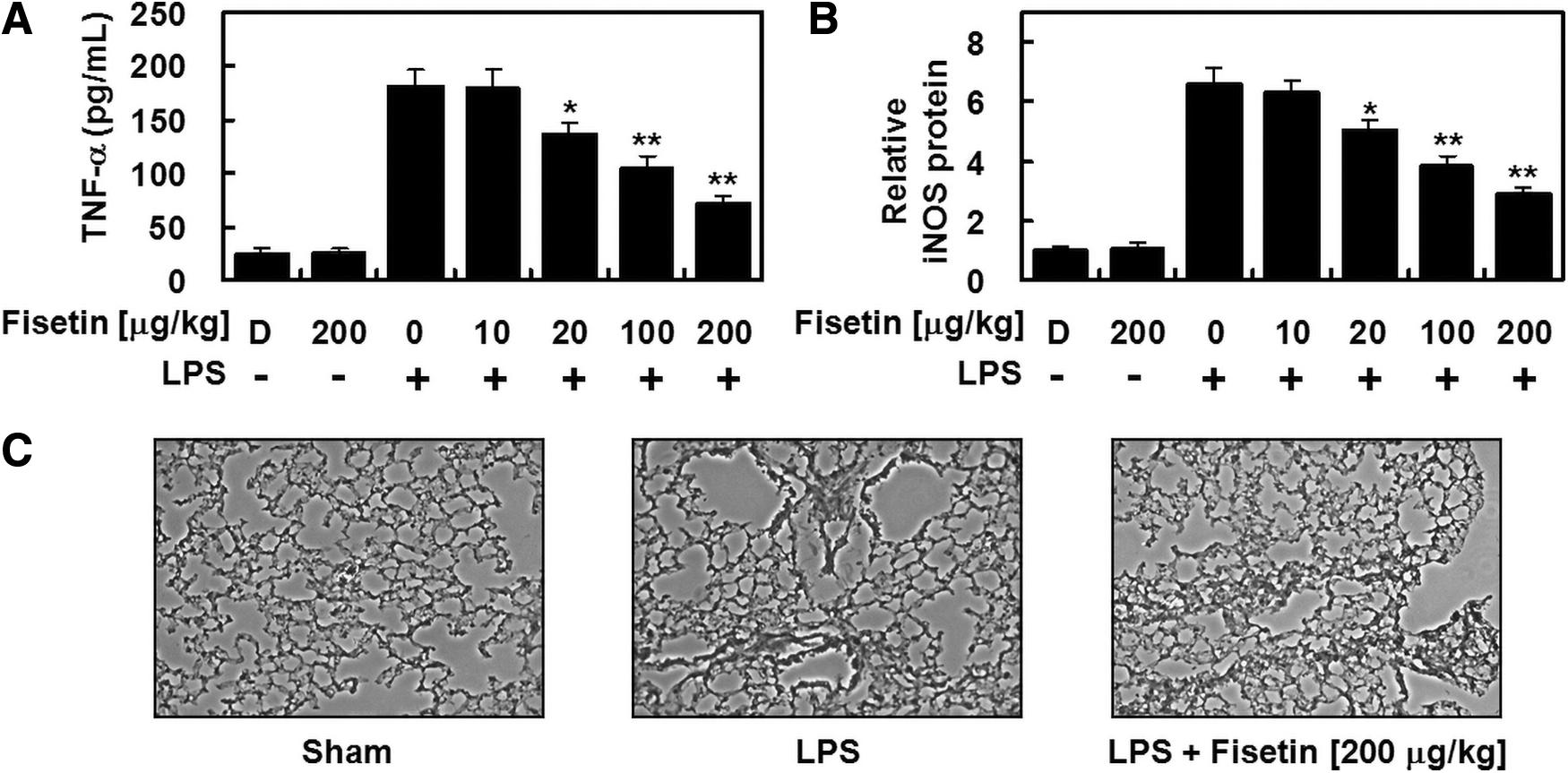
Fisetin suppressed the levels of TNF-α and iNOS, and reduced lung tissue injury in LPS-injected mice. Male mice were intraperitoneally injected with LPS (15 mg/kg, n = 5), and different concentrations of fisetin (10–200 μg/kg, n = 5) were intravenously administered 6 h later. LPS was not injected in control mice (n = 5). The mice were anesthetized and sacrificed 24 h after LPS challenge, and BALFs and lung tissues were collected. Levels of TNF-α
Discussion
Our study demonstrated that fisetin concentration-dependently induced the expression of HO-1 in HPAECs and suppressed NF-κB, COX2/PGE2, and iNOS/NO activity, induced by LPS treatment. NF-κB is known to be involved in various physiological and pathological processes such as cell adhesion, cell proliferation, developmental signals, host immune responses, and protection against cell apoptosis. 22 In addition, during inflammatory processes, NF-κB upregulates the generation of proinflammatory mediators such as ROS, COX2, NO, TNF-α, and IL-6. High levels of NO also play a crucial function in pulmonary inflammatory responses by regulating secretion of chemokines, while COX-2 and iNOS induced by LPS treatment require sufficient activation of NF-κB signaling. As stated above, the activation of the JAK/STAT signaling is essential for upregulation of COX-2 and iNOS with LPS stimulation. 18,19 Thus, the suppressive effects of fisetin on the levels of ILs, iNOS, and COX2 were mediated by its suppression of NF-κB signaling and STAT1 phosphorylation. We show that suppressive effects of fisetin on iNOS expression and NO production were significantly reversed by the specific knockdown of the HO-1. We also show the suppressive effect of fisetin on iNOS and TNF-α levels in the BALFs of LPS-treated lung injury model. Taken together, our findings suggest the importance of HO-1 in the suppression of inflammatory responses, and also emphasize that TNF-α is a potentially important mediator of HO-1 functions.
In conclusion, our study shows that fisetin effectively suppressed representative proinflammatory markers, including NO, iNOS, and IL1-β, in LPS-activated HPAECs and iNOS and TNF-α in LPS-injected mice tissue. Thus, we suggest that fisetin may be a potentially important drug candidate for inflammatory disorders, particularly vascular lung injury.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This research was supported by the National Research Foundation of Korea (NRF) grant funded by the Korea government (2020R1A2C1004131 and 2020R1F1A1071980) and by the Bio & Medical Technology Development Program of the National Research Foundation (NRF) funded by the Ministry of Science & ICT (2017M3A9G8083382).