
Abstract
Pterostilbene is a natural compound contained in various dietary sources that has received tremendous attention due to its antioxidant properties with promising benefits in cancers and vascular diseases. Currently, little is known about pterostilbene-associated neuroimmune endocrine effects. We aimed to examine the efficacy of pterostilbene for improving stress-related behaviors, neuroinflammation, and hormonal changes in a mouse stress model. To evaluate the efficacy of oral administration of pterostilbene or vehicle for 16 days for improving behavior, inflammation, and hypothalamic–pituitary–adrenal (HPA) axis hyperactivity, mice were divided into a normal control group or one of five restraint stress groups—the vehicle group; the 20, 40, or 80 mg/[kg·day] pterostilbene treatment group; or the 20 mg/[kg·day] resveratrol treatment group. Open field and forced swimming tests were conducted. Hippocampal brain-derived neurotrophic factor (BDNF) levels, endocrine hormone levels, oxidative stress parameters, and histopathological features were assessed. Oral pterostilbene administration significantly increased the measured times in the open field and forced swimming tests, elevated the BDNF levels, decreased the inducible nitric oxide synthase and superoxide dismutase levels in the brain, and reduced the plasma adrenocorticotropic hormone and corticosterone levels. Compared with vehicle treatment, pterostilbene dose dependently increased the numbers of neurons and decreased the numbers of glial and tumor necrosis factor alpha-immunolabeled cells in the hypothalamus. These findings suggest that pterostilbene may effectively modulate stress-related abnormal behaviors, neuroinflammation, and HPA axis hyperactivity.
Introduction
Pterostilbene (trans-3,5
Recently, the antioxidant activity of pterostilbene has been seen in both in vitro and in vivo studies, demonstrating preventative and therapeutic benefits. 8 In particular, pterostilbene has been reported to have various beneficial antineoplastic effects in several common malignancies, 9 to attenuate cardiovascular disease, 10,11 and to ameliorate dysregulated glucose metabolism. 12 Pterostilbene has been reported to have pharmacological properties similar to those of resveratrol due to their structural similarities. Both pterostilbene and resveratrol are stilbenoids, but pterostilbene contains two methoxy groups and one hydroxyl group, whereas resveratrol has three hydroxyl groups. The presence of two methoxy groups makes pterostilbene more lipophilic than resveratrol, increasing its oral absorption and its potential for cellular uptake. 2,13 The half-life of pterostilbene is also longer than that of resveratrol, 105–14 min, 14 because resveratrol is relatively more rapidly metabolized by phase II enzymes in the intestine and liver. 15 The metabolites of pterostilbene, such as those of resveratrol, can cross the blood–brain barrier (BBB) via Uridine 5′-diphospho-glucuronosyltransferases (UDP-glucuronosyltransferases); pterostilbene has advantages within the brain, along with better intestinal absorption than resveratrol. 16
Stress can be induced by internal or external sources, such as psychological or physical stimuli, and is implicated in disrupting homeostasis, affecting both daily performance and, more seriously, psychosocial functions. 17 In addition, measuring biomarkers related to neuroinflammatory and neuroendocrinological characteristics may be important when assessing behavioral and affective changes. 18 The restraint stress model in rodents has been known to combine both the physical and emotional aspects of stress without the use of painful stimuli, causing a robust increase in basal cellular oxidative stress. 19 Oxidative stress has been observed to trigger neuroinflammation and hypothalamic–pituitary–adrenal (HPA) axis hyperactivity and may be a surrogate target for antistress interventions. 19 Restraint stress and immobilization models are among the most commonly adopted protocols to induce stress-related behavioral and biochemical changes. 20 –22 Restraint stress protocols produce inescapable physical and mental stress for 7–21 days while avoiding habituation, with a low rate of adaptation. 17,23,24
To date, data on the effects of pterostilbene in brain health are focused on the neurodegenerative conditions. 25 –27 However, little is known about the neuroinflammatory and neuroendocrine changes associated with the effects of pterostilbene on stress-related behaviors in a young mouse model. This study was performed to evaluate the efficacy of repeated oral pterostilbene administration for improving stress-related behaviors, neuroinflammation, and HPA axis hyperactivity in a restraint stress mouse model.
Materials and Methods
Animals
Male specific pathogen-free Institute of Cancer Research (ICR) mice (5 weeks old, 24.6–28.7 g) were obtained from OrientBio, Inc. (Seongnam-si, Korea). After weighing and examination of their external appearance, the animals were acclimated to the laboratory conditions. Body weight and mortality were observed daily for the 6 days of the acclimation period. This study was performed in the behavior test room of the barrier animal facility area no. 3 of the Gyeonggi Bio Center, and the animals were housed in a room with a constant temperature of 23°C ± 3°C, relative humidity of 55% ± 15%, 12-h light/12-h dark cycle (from 08:00 to 20:00), 150–300 l × of luminous intensity, and 10–20 air changes per hour. Animals were offered Teklad Certified Irradiated Global 18% Protein Rodent Diet (2918C; ENVIGO, London, United Kingdom) purchased from DooYeol Biotech Co., Ltd. (Seoul, Korea) ad libitum. Water disinfected by ultraviolet sterilization and ultrafiltration was provided ad libitum via a water bottle and examined by the authorized official of the Gyeonggi-do Institute of Health and Environment (Suwon-si, Korea) to attest that the quality satisfied the standards for drinking water. All animals were housed in polycarbonate cages (W 170 × L 235 × H 125 mm) during the acclimation, dosing, and observation period. The present study was approved by the Institutional Animal Care and Use Committee (IACUC) of ChemOn (approval no. 2019-04-007). The procedures were carried out in accordance with the National Institutes of Health Guide for the Care and Use of Laboratory Animals (NIH Publications No. 80-23) revised in 1996, the UK Animals (Scientific Procedures) Act of 1986 and associated guidelines, or with the European Communities Council Directive of 24 November 1986 (86/609/EEC). Furthermore, all efforts were made to minimize the number and suffering of animals used.
Reagents
Pterostilbene and resveratrol were obtained from ChemOn, Inc. (Suwon-si, Korea) and stored under refrigeration protected from moisture and light. For oral administration, pterostilbene and resveratrol were dissolved in a cremophor (a vehicle for poorly water-soluble reagents; Sigma Chemical Co., St. Louis, MO, USA) mixture (1:1:8 cremophor:ethanol:saline). Pterostilbene and resveratrol were administered once daily for 16 days by oral gavage using a 1 mL syringe. The administered volume was calculated based on the dose (10 mL/kg) and the body weight measured on the day of administration.
Experimental design
To evaluate the efficacy of repeated oral pterostilbene administration for improving stress-related behaviors, neuroinflammation, and HPA axis hyperactivity, the animals were divided into six groups, including the normal control group (G1) and five restraint stress groups (G2–G6). The restraint stress groups consisted of the vehicle group (G2); groups treated with the test article pterostilbene at 20, 40, and 80 mg/[kg·day] (G3, G4, and G5, respectively), and a group treated with the reference compound, resveratrol at 20 mg/[kg·day] (G6), based on the low-dose range used in previous cancer studies. 9 Each group comprised seven mice (Fig. 1).
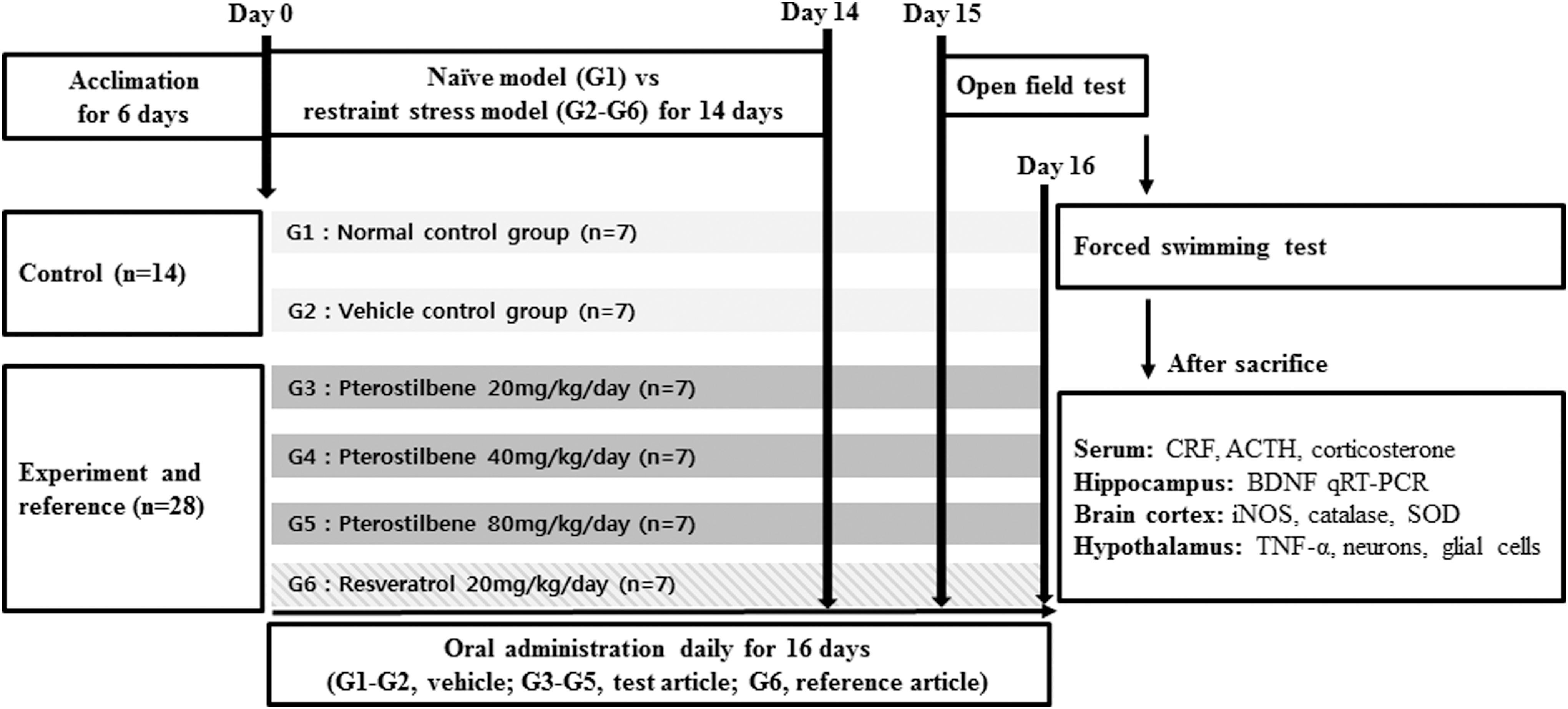
Flowchart of the study design. ACTH, adrenocorticotropic hormone; BDNF, brain-derived neurotrophic factor; CRF, corticotropin-releasing factor; iNOS, inducible nitric oxide synthase; qRT-PCR, quantitative real-time polymerase chain reaction; SOD, superoxide dismutase; TNF-α, tumor necrosis factor alpha.
To elicit chronic stress, 28,29 animals were restrained in a ventilated polypropylene 50 mL conical tube. One hour after administration of either the vehicle only or the reagents, animals were restrained for 3 h. The restraint procedure was performed daily for 14 days between 10:00 and 13:00. The protocols for the administration of each reagent and the restraint procedure were performed identically under an identical environment for all restraint stress groups (G2–G6). No food and water were provided to the control group (G1), while G2–G6 being under the restraint procedure.
Body weight measurement and behavioral tests
Clinical signs and body weight
The mortality of all animals was monitored once daily and individually recorded for 16 days. The day of the first drug administration was defined as day 1. The body weight was measured once weekly.
Open field test
On day 15, the open field test was conducted 1 h after the drug or vehicle administration. The test was conducted in an acrylic box (40 × 40 × 40 cm) open to the air. The floor of the box was divided by lines into 16 squares (10 × 10 cm). The animals were placed in the center of the box and allowed to explore the environment for 5 min. 30,31 Movements were recorded for later analysis using a video tracking system (SMART ver. 3.6.1; Harvard Apparatus, Holliston, MA, USA). The time spent in the central area (“time in center”) of the apparatus was calculated.
Forced swimming test
On day 16, the forced swimming test was conducted for 10 min in a swimming pool with a diameter of 18 cm and a height of 50 cm filled to a depth of 20 cm with warm water (23 ± 2°C). One hour after drug or vehicle treatment, mice were placed individually in the pool. A weight (5% of the body weight) was attached to the tail root of the tested mouse. The swimming time, defined as the time until the mouse could not stay above the water surface for more than 5 sec, was measured. 32,33
Blood sampling and tissue collection
Endocrine hormone measurement
Mice were euthanized after the forced swimming test. After the mice were anesthetized with isoflurane, blood was drawn from the inferior vena cava. The collected blood was injected into a vacutainer tube containing a clot activator and allowed to stand at room temperature for 15–20 min. The coagulated blood was centrifuged for 10 min, and the serum was collected and stored in a freezer at −80°C for the measurement of endocrine hormone levels. Hormones were measured using enzyme-linked immunosorbent assay (ELISA) kits for corticotropin-releasing factor (CRF; Cusabio Biotech Co., Ltd., Houston, TX, USA), adrenocorticotropic hormone (ACTH; Cusabio Biotech Co., Ltd.), and corticosterone (Arbor Assays, Ann Arbor, MI, USA).
Brain-derived neurotrophic factor and oxidative stress measurements
After blood collection, brains were harvested. The hippocampus and cortex were frozen, and the hypothalamus was fixed in 10% neutral buffered formalin solution. Hippocampal brain-derived neurotrophic factor (BDNF) was quantitated by quantitative real-time polymerase chain reaction (qRT-PCR) using hippocampal samples. The primer sequences and product size were as follows: Forward: 5′-GAC AAG GCA ACT TGG CCT AC-3′ Reverse: 5′-CCT GTC ACA CAC GCT CAG CTC-3′ Product size: 356 base pairs
The levels of inducible nitric oxide synthase (iNOS), catalase, and superoxide dismutase (SOD) in the brain cortex were measured using ELISA kits for iNOS, catalase, and SOD, respectively (all Cusabio Biotech Co., Ltd.).
Histopathological assessment
Approximately equal regions of individual hypothalamus tissues were cross-trimmed. All hypothalamus tissue samples were fixed in 10% neutral buffered formalin solution for 24 h. Subsequently, paraffin blocks were prepared using an automated tissue processor (Shandon Citadel 2000; Thermo Scientific, Waltham, MA, USA) and embedding center (Shandon Histostar; Thermo Scientific), and 3–4-μm sections were prepared as two serial sections from each paraffin-embedded hypothalamus using an automated microtome (RM2255; Leica Biosystems, Nussloch, Germany). Representative sections were stained with 0.1% cresyl violet for general histopathology or with avidin/biotin/peroxidase complex-based immunohistochemical staining for tumor necrosis factor alpha (TNF-α) according to previously established methods. Samples were observed using a light microscope (Eclipse 80i; Nikon, Tokyo, Japan).
The mean neuron and glial cell numbers (cells/mm2) in the paraventricular nucleus (PVN) regions were calculated for histomorphometric analysis of the cresyl violet-stained samples using a computer-assisted image analysis program. Furthermore, cells with over 20% immunoreactivity for anti-TNF-α antibodies were considered immunopositive, and the mean numbers of immunopositive cells (cells/mm2) in the PVN regions were determined by an automated image analyzer. One histological field per hypothalamus section, with a total of seven hypothalamus tissue samples per group, was considered for statistical analysis in the current study. The histopathologist was blinded to the experimental conditions when analyzing the samples.
Statistical analyses
All data were tested for a normal distribution and homogeneity of variance, and abnormal data were eliminated. The results are expressed as the means ± standard errors of the mean. Comparisons between the vehicle group and the normal control group were performed with Student's t-test. Then, one-way analysis of variance followed by Dunnett's test was used for comparisons between the vehicle group and the experimental groups. Statistical analyses were conducted using SPSS 12.0K for Windows software (SPSS, Inc., Chicago, IL, USA). Differences with a P value of <.05 were considered significant.
Results
Clinical signs and body weight
No mortality was observed in any of the six groups of mice. The vehicle group (G2) exhibited a significant decrease in body weight compared with the normal control group (G1) on day 8 and 15 (P < .01 and P < .05, respectively). The body weight changes in the test article groups (G3–G5) and the reference article group (G6) did not significantly differ from those in the vehicle group (G2; Table 1).
Changes in Weights
The data are expressed as the mean ± SEM. The results were statistically analyzed using Student's t-test and one-way ANOVA.
Significantly different from G1, P < .01. **Significantly different from G1, P < .05.
ANOVA, analysis of variance; SEM, standard errors of the mean.
Open field test
The vehicle group (G2) had a significantly decreased mean value for the parameter “time in center” compared with the normal control group (G1) (P < .01). The group treated with 80 mg/[kg·day] pterostilbene (G5) spent significantly more time in the center than the vehicle group (G2, P < .05), whereas this parameter was not significantly different between the 20 mg/[kg·day] pterostilbene treatment group (G3) and the vehicle group (G2). The 40 mg/[kg·day] pterostilbene treatment group (G4), as well as the reference article group (G6), exhibited a tendency toward an increased “time in center” compared with that in the vehicle group (G2; Fig. 2).
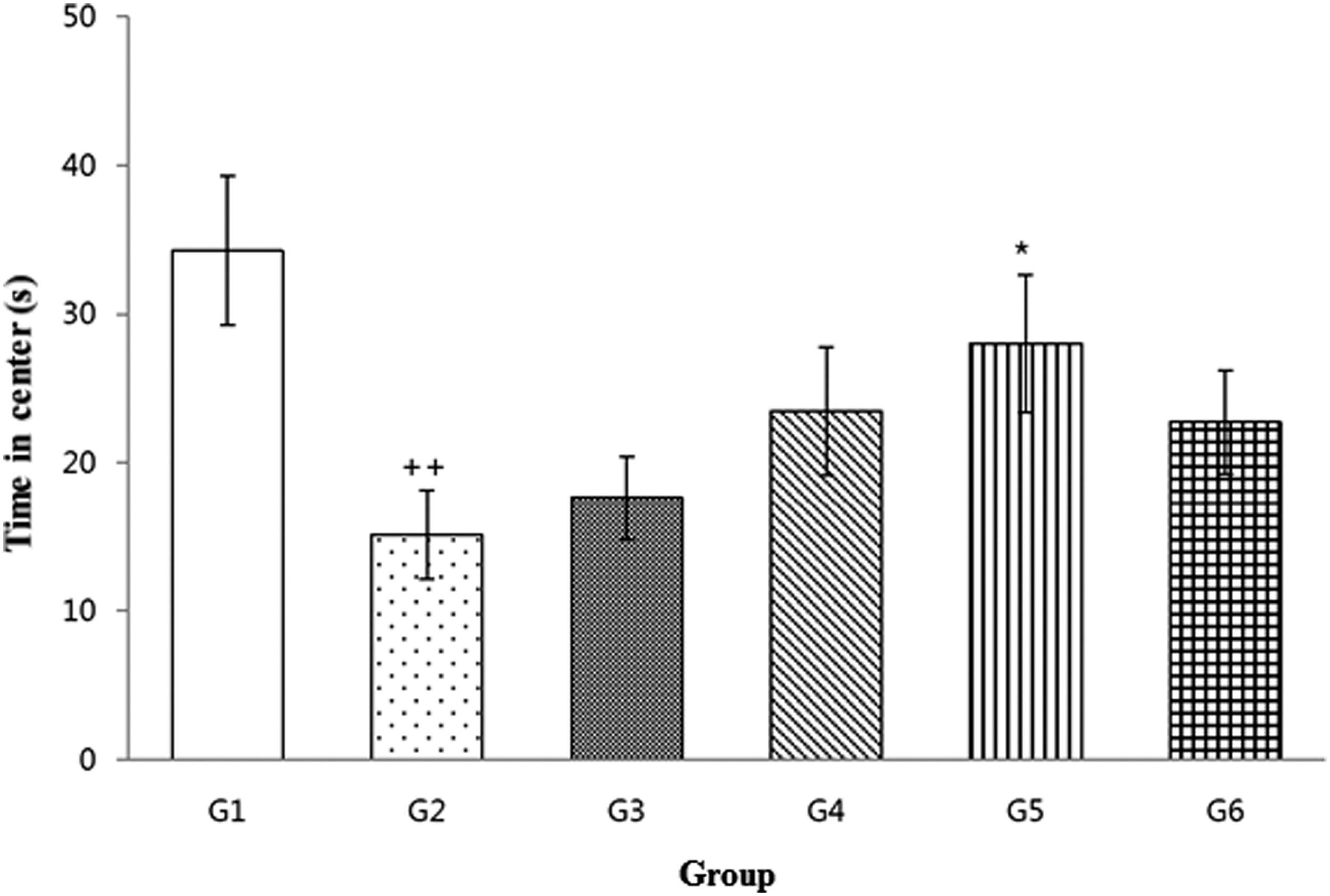
Open field test. The data are expressed as the mean ± SEM. The results were statistically analyzed using Student's t-test and one-way ANOVA. ++Significantly different from G1, P < .01; *significantly different from G2, P < .05. ANOVA, analysis of variance; SEM, standard errors of the mean.
Forced swimming test
The swimming time was significantly decreased in the vehicle group (G2) compared with the normal control group (G1) (P < .01). All test article (G3–G5) and reference article (G6) groups displayed a tendency toward an increased swimming time compared with that in the vehicle group (G2; Fig. 3).

Forced swimming test. The data are expressed as the mean ± SEM. The results were statistically analyzed using Student's t-test and one-way ANOVA. ++Significantly different from G1, P < .01.
Hippocampal BDNF expression
The vehicle group (G2) exhibited a significantly lower relative BDNF expression than the normal control group (G1) (P < .05). Treatment with 20 and 80 mg/[kg·day] pterostilbene (G3 and G5, respectively) significantly increased the relative BDNF expression level compared with that in the vehicle group (G2, P < .05), whereas the group treated with 40 mg/[kg/day] pterostilbene (G4) showed a mean relative BDNF expression level similar to and not significantly different from that in the vehicle control group (G2). The reference article group (G6) also exhibited a significantly higher relative BDNF expression level than the vehicle group (G2, P < .01; Fig. 4).

Relative BDNF expression in the hippocampus. The data are expressed as the mean ± SEM. The results were statistically analyzed using Student's t-test and one-way ANOVA. +Significantly different from G1, P < .05; *significantly different from G2, P < .05; **significantly different from G2, P < .01.
Endocrine hormonal assays
The vehicle group (G2) had significantly higher corticosterone levels than the normal control group (G1, P < .05). The test article groups (G3–G5) exhibited a tendency toward decreased corticosterone levels compared with those in the vehicle group (G2), whereas the reference article group (G6) showed a statistically significant difference compared with the vehicle group (G2, P < .05; Fig. 5A). Furthermore, we also measured the ACTH levels in blood. The vehicle group (G2) exhibited significantly higher ACTH levels than the normal control group (G1, P < .01). The group treated with 80 mg/[kg·day] pterostilbene (G5) exhibited significantly lower ACTH levels than the vehicle group (G2, P < .05), whereas for the other test article groups (G3–G4), a nonsignificant tendency toward reduced ACTH levels was observed with respect to the vehicle group. Moreover, the reference article group (G6) exhibited significantly lower ACTH levels than the vehicle group (G2, P < .01; Fig. 5B). However, the CRF levels in the vehicle group (G2) were not significantly different from those in any other group (G1 and G3–G6; Table 2).

Corticosterone
Endocrine Hormone Levels
The data are expressed as the mean ± SEM and were statistically analyzed using Student's t-test and one-way ANOVA.
Significantly different from G1, P < .05; ++significantly different from G1, P < .01; *significantly different from G2, P < .05; **significantly different from G2, P < .01.
ACTH, adrenocorticotropic hormone; CRF, corticotropin-releasing factor.
Measurement of oxidative stress in the cerebral cortex
Cerebral cortex samples from the vehicle group (G2) exhibited significantly higher iNOS levels than the normal control group (G1, P < .01). The iNOS levels in the 80 mg/[kg·day] pterostilbene group (G5) were significantly lower than those in the vehicle group (G2, P < .05). All other test article groups (G3 and G4) and the reference article group (G6) exhibited a trend toward decreased iNOS levels compared with those in the vehicle group (G2; Fig. 6A).
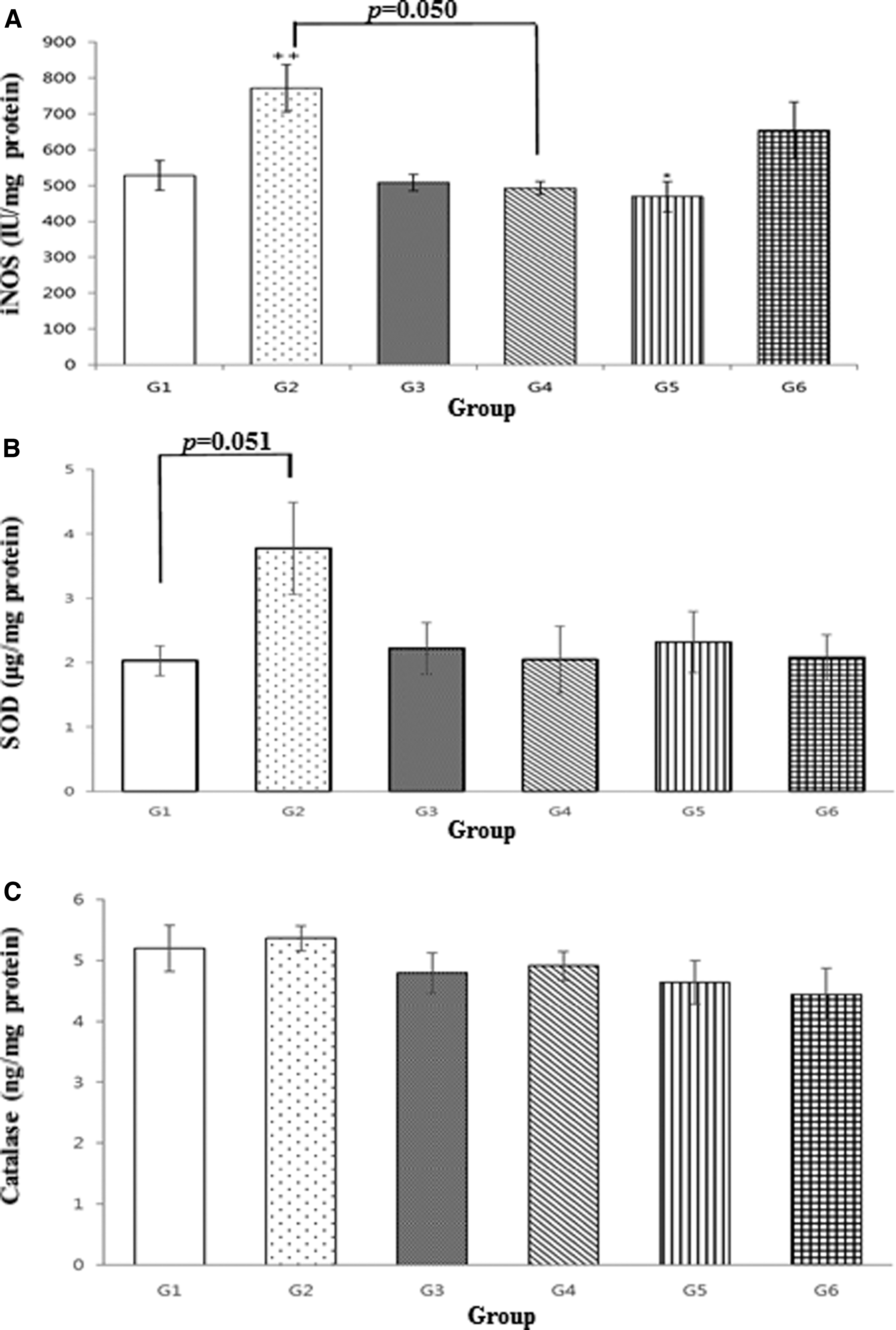
iNOS
In further measurements of the SOD levels, the vehicle group (G2) tended to exhibit higher levels than the normal control group (G1). By contrast, the test article (G3–G5) and reference article (G6) groups presented a tendency toward decreased SOD levels compared with those in the vehicle group (G2; Fig. 6B). However, the catalase levels in the vehicle group (G2) were not significantly different from those in any other group (G1 and G3–G6; Fig. 6C).
Histopathological analysis of hypothalamus tissue
We also determined the numbers of neurons in brain slices containing hypothalamus tissue. These numbers were significantly lower in the vehicle group (G2) than in the normal control group (G1, P < .01). This effect was reversed in the test article (G3–G5) and reference article (G6) groups, which exhibited significantly increased numbers of neurons compared with the vehicle group (G2, P < .01). By contrast, the numbers of glial cells were significantly higher in the vehicle group (G2) than in the normal control group (G1, P < .01). The groups treated with 40 and 80 mg/[kg·day] pterostilbene (G4 and G5) had significantly fewer glial cells than the vehicle group (G2, P < .01), whereas the group treated with 20 mg/[kg·day] pterostilbene (G3) showed only a tendency toward a decrease. Similar to the groups treated with higher pterostilbene concentrations, the reference article group (G6) exhibited significantly fewer glial cells than the vehicle group (G2, P < .05). Regarding the numbers of TNF-α immunopositive cells, the vehicle group (G2) had significantly higher numbers of immunolabeled cells than the normal control group (G1, P < .01). All test article groups (G3–G5) exhibited statistically significant decreases in the numbers of TNF-α-immunolabeled cells compared with the vehicle group (G2, P < .05 or P < .01). Similarly, the reference article group (G6) exhibited significantly fewer TNF-α-immunolabeled cells than the vehicle group (G2, P < .01; Fig. 7 and Table 3).
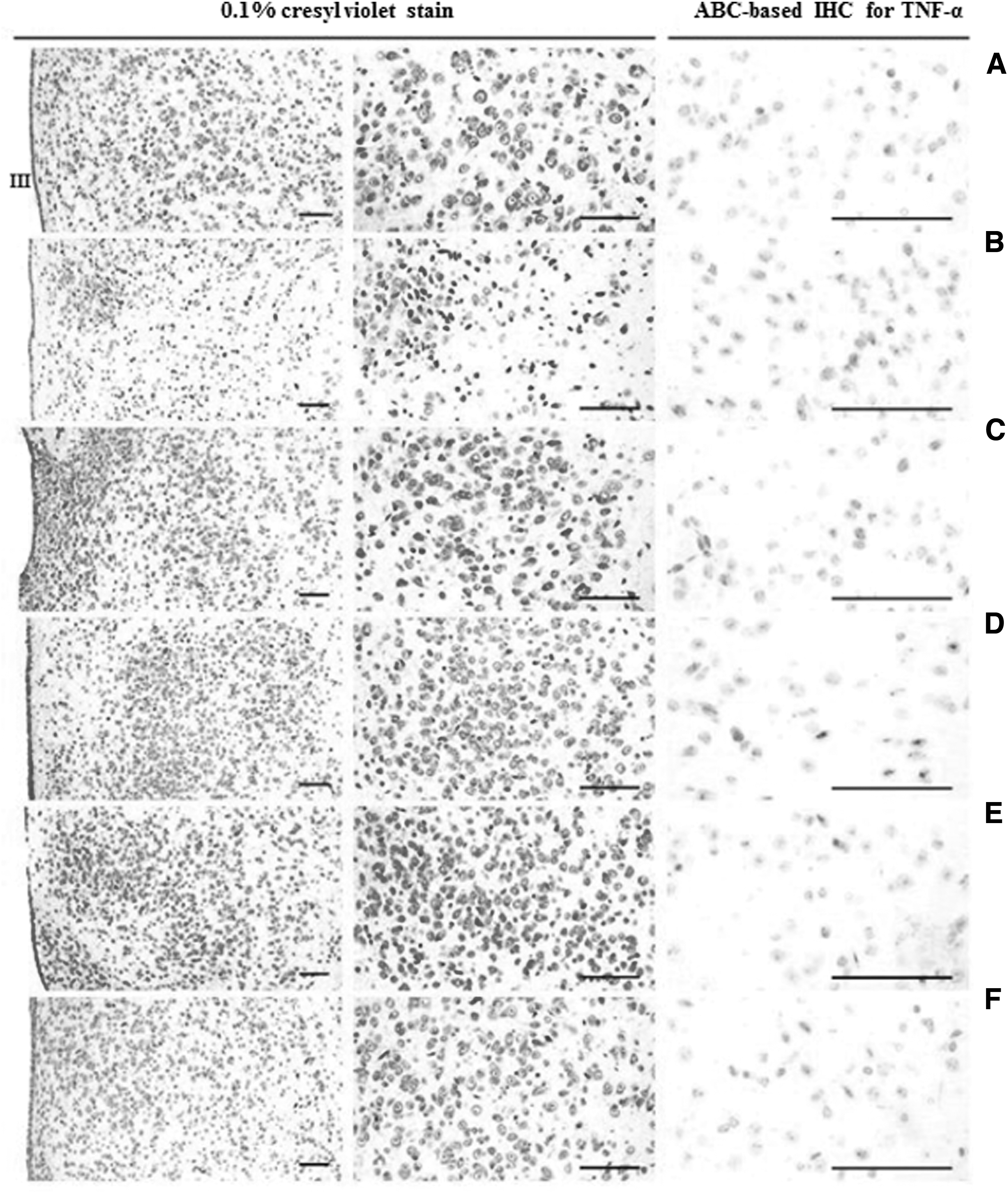
Representative histopathological profiles of the hypothalamic PVN regions.
Histomorphometric Analysis of the Hypothalamic Paraventricular Nucleus Regions
The data are expressed as the mean ± SEM and were statistically analyzed using Student's t-test and one-way ANOVA.
Significantly different from G1, P < .01; *significantly different from G2, P < .05; **significantly different from G2, P < .01.
TNF, tumor necrosis factor.
Discussion
We found that in a mouse stress model, oral pterostilbene administration ameliorated stress-related behavioral anxiety and despair, which were associated with neuroinflammatory modulations, including restoration of BDNF levels, reduction in the oxidative stress burden, and inhibition of HPA axis hyperactivity.
The stress model comprises the response of the animal to various harmful factors, the corresponding defense reactions, and the general maladaptation syndrome. The restraint stress model in rodents simultaneously induces physical and mental stress and is usually performed to study behaviors associated with anxiety and depression in humans, thus helping to assess pathogenic mechanisms and potential clinical drug effects. 34
This study was performed to evaluate the efficacy of repeated oral pterostilbene administration for 16 days on stress-related behaviors, neuroinflammation, and HPA axis hyperactivity in a mouse stress model. Compared with the normal control group, the stress-exposed vehicle group had a significantly decreased body weight on days 8 and 15, which may have been induced by prolonged physical and emotional stress. In the open field and forced swimming tests, the vehicle group spent significantly less time in the central area than the normal control group, and the swimming time was similarly reduced. Pterostilbene treatment partially reversed these effects; we observed a tendency toward increased times in the pterostilbene-treated groups compared with those in the vehicle group. The open field and forced swimming tests are commonly used to measure the effects of molecules in behavioral patterns, 32,35 which are in agreement with the results of other behavior test paradigms for anxiety and depression, respectivly. 30 The open field test is usually performed to evaluate general locomotor and exploratory behaviors, which are associated with anxiety-like behaviors in mouse models. 36 The forced swimming test has been extensively adopted because the immobility time during stressful conditions may reflect the tendency for behavioral despair or helplessness in animals. 37 Based on our study results, pterostilbene effectively improves stress-induced abnormal behaviors.
BDNF regulates the density of inhibitory synapses in organotypic hippocampal cultures and is important for modulating synaptic transmission and neuronal survival. 38,39 Animal studies have demonstrated that the pathophysiological changes in repeated stress situations or mood disorders include decreased BDNF levels, which may lead to decreased neurogenesis and increased remodeling of dendrites, thus inducing vulnerability to subsequent episodes of depression. 40,41 In our experiments, the vehicle group had significantly lower BDNF levels than the normal control group due to the stress conditions, whereas the pterostilbene treatment groups generally exhibited higher BDNF levels than the vehicle group. Pterostilbene effects on neurotrophic pathways in the hippocampus, besides BDNF levels, have been involved in the phosphorylation of extracellular signal-regulated kinase and cAMP responsive element binding protein. 25,42
Although the mechanisms underlying anxiety and depression are incompletely understood, stress is an important inducing factor and is usually accompanied by dysfunction of the neuroimmune-endocrine connection. 30 Indeed, HPA axis hyperactivity can be a crucial biological mechanism underlying maladaptive behavioral changes. 43 In our study, the vehicle group exhibited significantly higher ACTH and corticosterone levels than the normal control group. These stress-related effects were reversed by pterostilbene treatment, with significant decreases—or at least trends toward a decrease—in ACTH and corticosterone levels compared with those in the vehicle group. Therefore, pterostilbene is effective in reversing stress-induced abnormalities in BDNF, ACTH, and corticosterone levels.
The HPA axis can be affected by many factors, such as cytokines and nitric oxide (NO). Cytokines, which are generally secreted by immune cells, can also be produced and secreted by nonimmune cells to signal to neuroimmune cells. Proinflammatory cytokines, including interleukin-1β, interleukin-6, and TNF-α, are secreted by all components of the HPA axis. 30,44 NO, acting as a neurotransmitter in the brain, is generated by neuronal NOS (nNOS) and iNOS in the brain and is involved in the regulation of the HPA axis, but overexpression of nNOS and iNOS can lead to HPA hyperactivity with stress-related behavioral changes. 45 In our measurements of iNOS, SOD, and catalase levels, the vehicle group presented significant increases—or trends toward increases—in these levels compared with those in the normal control group. Pterostilbene treatment either significantly decreased or caused a trend toward a decrease in the levels of oxidative stress markers compared with those in the vehicle group. Our data suggest that administration of pterostilbene reduced reactive oxygen species generation and macrophage activation induced by restraint stress.
Finally, we analyzed histopathological changes in the hypothalamus. Compared with the normal control group, the vehicle group had significantly decreased numbers of neurons and, simultaneously, increased numbers of glial cells and TNF-α-immunolabeled cells in the hypothalamic PVN regions. This pattern suggests that restraint stress can induce neuroinflammation-related gliosis and neuronal loss in the hypothalamic PVN. However, these effects, stress-induced gliosis and neuronal loss in the hypothalamic PVN, were significantly inhibited by pterostilbene in a dose-dependent manner. These results suggest that the oral administration of pterostilbene exerts beneficial, dose-dependent, protective effects against stress-induced damage to the hypothalamic PVN, potentially indicating anti-inflammatory activity via the inhibition of TNF-α, a proinflammatory cytokine and a marker of gliosis. In addition, pterostilbene exerts favorable protective effects against neuroinflammation in a dose-dependent manner, while additionally exhibiting beneficial properties that reduce oxidative stress in the brain. Based on the good safety profile of pterostilbene, 46 higher dosages may be considered in a future study. Taken together, these observations indicate that pterostilbene reverses stress-related upregulation of oxidative stress markers and hypothalamic hormones. In addition, pterostilbene administration prevented stress-induced upregulation of proinflammatory markers and neuronal loss in the hypothalamus. In addition, the effects of pterostilbene on mitochondria may be, in part, linked to an increase in the oxidative stress-related threshold for mitochondrial permeability transition, thereby preventing the potential apoptosis of vulnerable cells. 8
This study provided the first convincing data that induced stress alters neuroinflammatory and neuroendocrinological parameters and that these changes are, at least partially, reversed by oral administration of pterostilbene, a substance that can be found in natural products such as blueberries. This finding may lead to novel insights into the pathophysiology of depression and anxiety, ultimately leading to new therapeutic approaches. However, we should acknowledge the limitations of our study. To evaluate stress-related hormonal changes, blood samples were obtained from the inferior vena cava. The pterostilbene treatment groups had lower levels of ACTH and corticosterone but not CRF than the vehicle group. To evaluate CRF activity more accurately, examination of CRF receptors or CRF gene transcription in the brain is a better assessment approach. Second, resveratrol as the reference article showed inconsistent effects when compared with the same dose as pterostilbene. Although pterostilbene has a relatively higher oral bioavailability than resveratrol, we did not perform quantitative assay on the degree of BBB penetration, and despite their structural similarity, different mechanisms may have worked. Further research is needed to assess the biochemical mechanisms with dose responses in the brain. In addition, it is necessary to assess whether there are any synergistic effects among them and compare them with other natural compounds that have shown anxiolytic effects.
In conclusion, the oral administration of pterostilbene for 16 days in restraint-induced stress mice significantly increased the times in the open field and forced swimming tests, elevated the BDNF levels, decreased iNOS and SOD levels in the cerebral cortex, and reduced plasma levels of ACTH and corticosterone. In addition, pterostilbene was shown to dose dependently increase the numbers of neurons and decrease the numbers of glial cells and TNF-α immunolabeled cells in hypothalamic PVN regions in comparison with the vehicle group. These findings suggest that pterostilbene could be an effective treatment option in stress-related abnormal behaviors that are based on the modulation of neuroinflammation and hyperactivity of the HPA axis.
Footnotes
Acknowledgment
We are very grateful to the ChemOn Nonclinical Research Institute in the Gyeonggi Bio Center for the superb laboratory technical help.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.