
Abstract
Isoamylamine (IA) is an aliphatic monoamine molecule present in cheese, eggs, and wine. It belongs to the family of polyamines and also can be synthesized endogenously. It has been known that regulation of polyamines in cells is related to cell cycle and tumor formation. Malignant melanoma is difficult to treat and easily resistant to chemotherapy/radiotherapy through autophagy. In this study, we aim to clarify whether IA has a growth control effect on melanoma tumor cells and the regulatory mechanism. We treated B16-F1 melanoma cells with IA at concentrations of 0, 200, 400, and 600 ppm for 24 h. The 3-(4, 5-dimethylthiazol-2-yl)-2, 5-diphenyltetrazolium bromide (MTT) assay was checked for cell viability and results showed that IA has an inhibitory effect on B16-F1 melanoma cells. The signaling molecules, which included Raf/MEK/ERK, were activated, while MSK1 and protein kinase B (AKT) were suppressed. Autophagy was also confirmed to be induced by IA. The acridine orange stain-positive cells were increased and BECN-1/LC3 upregulated. The data also showed that the autophagy regulatory molecule, 5′-adenosine monophosphate-activated protein kinase (AMPK), was induced after IA treatment, so we used dorsomorphin to inhibit AMPK and found that it could suppress autophagy. In conclusion, IA has an effect of inducing autophagy in B16-F1 cells and it is regulated through AMPK.
Introduction
Isoamylamine (IA; 1
The mitogen-activated protein kinase (MAPK)/extracellular signal-regulated kinase (ERK)/90 kDa ribosomal S6 kinase (RSK) pathway and mitogen- and stress-activated protein kinase (MSK1) are signaling molecules that are involved in cell proliferation. MSK1 is more related to regulation of the process of autophagy when cells are stimulated by cellular stress or by other proinflammatory/cytokine factors. 8 Previous studies have indicated that when cardiac myocytes encounter ischemia or hypoxia, MSK1 would get activated and induce autophagy, leading to cell survival in a stressful environment. 9 It is also seen that in glioblastoma multiforme (GBM; an aggressive brain tumor), MSK1 activation is increased and is resistant to PI3K/mTOR inhibition. This outcome has resulted in limited efficacy during the clinical use of PI3K/mTOR inhibitor as a single agent for treatment. 10 Another study has pointed out that activation of 5′-adenosine monophosphate (AMP)-activated protein kinase (AMPK), a metabolic energy sensor that can be activated by cellular adenosine triphosphate decrease/AMP increase, leads the cells toward autophagy, and inhibition of AMPK-dependent autophagy can promote oncogenic Ras, causing resistance in tumor regression. 11,12 According to the study, it was observed that trace IA has an effect on activating the intracellular cAMP level and is related to insulin secretion. 13 It suggested that IA might induce activation of AMPK through the metabolism of AMP, but it is still unconfirmed.
Melanoma is a tumor of melanocytes, which produces melanin in the skin and other organs. Metastatic melanoma is associated with poor prognosis, and treatment of malignant melanoma is difficult. 14,15 The development of melanoma in hypoxic or acidic environments is greatly related to the activity of inducing autophagy. 11,16 Few studies have also shown that some therapeutic chemicals or drugs reduce melanoma cell proliferation by inducing autophagy. 17,18 Ma et al. measured autophagy in melanoma cells to predict their invasiveness, chemotherapy resistance, and survival. 19 Regarding regulation of the signal transduction pathway, Chen et al. showed that activation of AMPK could inhibit melanoma cell proliferation. 20 Other studies have also indicated that liver kinase B (LKB)/p-AMPK and p-Unc-51-like autophagy-activating kinase 1 (ULK) were activated during the process of autophagy development in melanoma. 21 The activation of AMPK due to metabolic stress conditions and inhibition of serine/threonine protein kinase B-Raf (BRAF) signaling (the upstream for LKB) would have synergistic effects, promoting apoptosis to suppress melanoma. 22 However, BRAF inhibitors, which are used clinically to treat melanoma tumors, are ineffective in dealing with BRAF V600E mutation (about 50% of melanoma), which is prone to relapse because of this. 21,23 Thus, autophagy is a potential prognostic factor and therapeutic target for treating melanoma and it is significantly related to AMPK regulation.
The above information highlights that IA may play a role in regulation of cell viability. It is worth exploring to verify whether IA regulates melanoma tumor cell viability through signaling transduction molecules. In this study, we treated B16-F1 melanoma tumor cells with IA and found that there was a growth inhibitory effect. Hence, we aim to clarify whether IA is involved in the ERK, MSK1, and AMPK pathways, leading to autophagy.
Materials and Methods
Materials
The molecular formula of IA is C5H13N and the molecular weight is 87.16 g/mol (Fig. 1). We purchased IA from Sigma, St. Louis, MO, USA (Cat NO. M820716, 99% purity), and it was dissolved in H2O to obtain a stock solution of 104 ppm. Dorsomorphin (Compound C; ab120843), a potent, selective, and reversible AMPK inhibitor, was purchased from Abcam (Cambridge, United Kingdom). Other reagents were purchased from Sigma or Bio-Rad (Hercules, CA, USA), unless otherwise indicated. Antibodies against pc-Raf (Ser338), p-MEK1/2 (Ser217/221), p-p44/42-MAPK (Thr202/Tyr204), p-p90RSK (Ser380), p-MSK (Thr581), LKB1, Becline-1 (BECN-1), microtubule-associated protein light chain 3 (LC3A/B), p-AMPKα ( Thr172), and fatty acid synthase (FASN) were purchased from Cell Signaling Technology (Denver, MA, USA). Anti-β-actin and horseradish peroxidase (HRP)-conjugated secondary antibodies were obtained from Sigma Co.
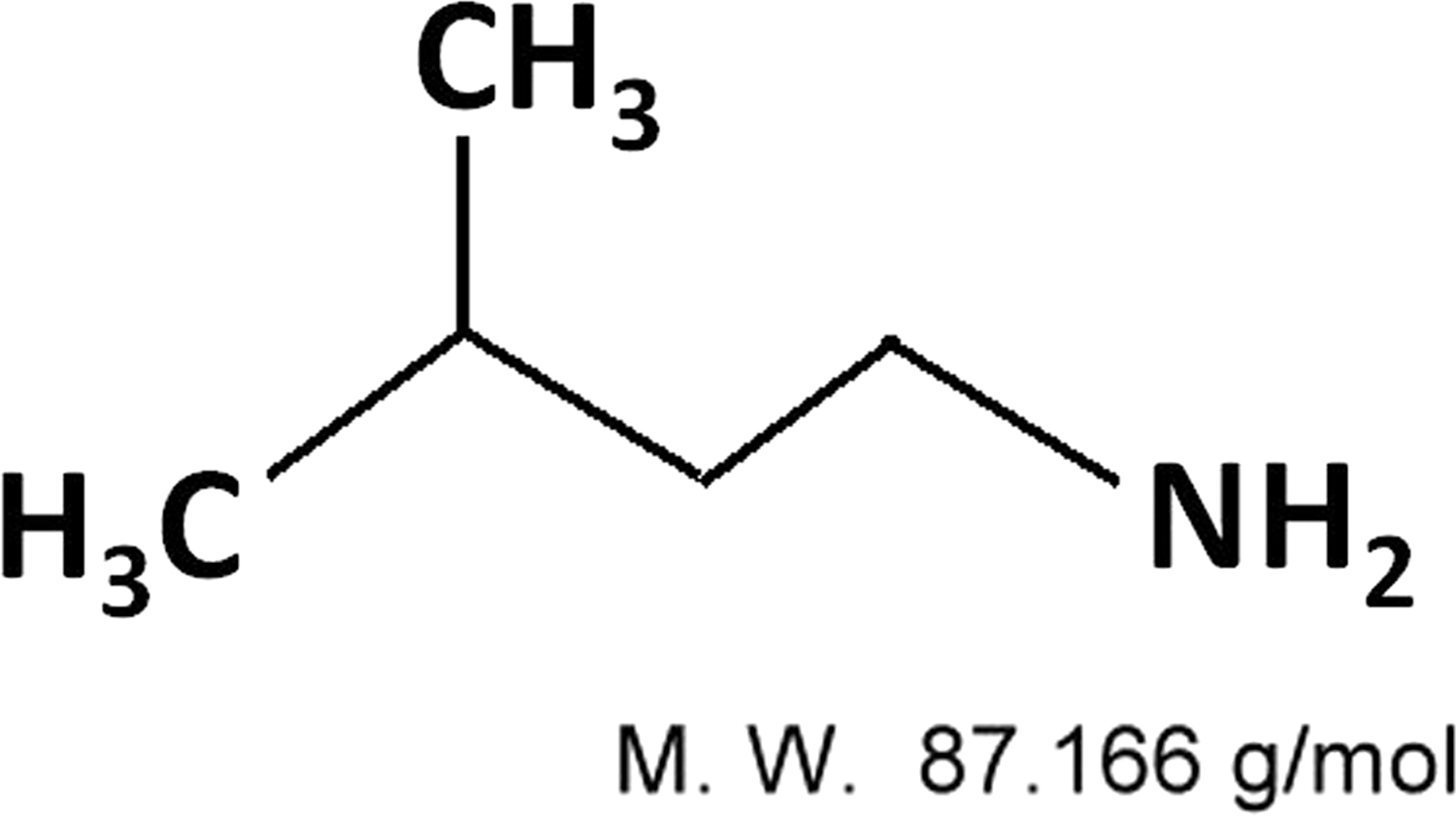
The chemical structure of IA. IA, isoamylamine.
Methods
Cell culture and treatment
Mouse melanoma B16-F1 cells (epithelial cells) were maintained in Dulbecco's modified Eagle's medium (Gibco/BRL, Gaithersburg, MD, USA) supplemented with 10% fetal bovine serum and antibiotics (100 U/mL penicillin and 100 μg/mL streptomycin) and cultured at 37°C in 5% CO2. Next, B16-F1 cells were seeded (density, 60 × 104 cells) in 60-mm Petri dishes for 18 h before initiating treatment with IA.
Wound healing and invasion assays
Wound healing and invasion assays were performed as described previously. 24 B16-F1 cells were grown into confluent monolayers in six-well Petri dishes containing a serum-free medium for 18 h, after which the medium was replaced with serum-containing medium. Next, the cells were divided into the following groups: untreated control and 200, 400, and 600 ppm IA-treated cells, respectively. Cells in the monolayer were disrupted (wounded) by scraping it with the P200 micropipette tip and washed twice with phosphate-buffered saline (PBS) at indicated time points (0, 8, 16, and 24 h) after scraping the cells. The number of cells in the denuded (scraped) zone of each Petri dish was counted at 100 × magnification (in a blinded manner). Cells in each dish were counted three times to ensure accuracy.
Cell viability (MTT assay)
B16-F1 cells were seeded (density, 5 × 104 cells/well) in 24-well plates and treated with the indicated concentrations of IA for 24 h. After incubation, cell viability was determined by performing the MTT [3-(4, 5-dimethylthiazol-2-yl)-2, 5-diphenyltetrazolium bromide] assay. Briefly, 5 mg/mL MTT solution was added to each well, and cells were incubated for 4 h at 37°C. After washing with PBS, the cells with the purple blue formazan crystals were dissolved in 1 mL of isopropanol, and absorbance was measured at 563 nm. Cell viability was proportional to the amount of formazan present in cells.
Acridine orange staining
Autophagy was assessed by detecting and quantifying acidic vesicular organelles (AVOs; lysosomes and autophagolysosomes) and by performing immunofluorescence analysis (acridine orange [AO] staining). Cells were cultured and treated with IA and then stained with AO (final concentration: 1 μM), followed by incubation for 15 min, as mentioned previously. 18 Photographs were obtained using a fluorescence microscope equipped with a 340/380 nm excitation filter (Nikon Diaphot-300; Nikon, Japan). The percentage of AVO-positive cells was calculated by dividing autophagic cells with the total number of cells and then multiplying it with 100. Each experiment was conducted thrice and at least 300 cells were counted in each experiment.
Immunoblotting assay
Total proteins were extracted by lysing the cells in lysis buffer (1% NP-40, 50 mM Tris base, 0.1% sodium dodecyl sulfate [SDS], 0.5% deoxycholic acid, and 150 mM NaCl [pH 7.5]) containing 17 μg/mL leupeptin and 10 μg/mL sodium orthovanadate. Protein concentration was measured by using the Bradford protein assay kit. Equal amounts of protein (50 μg) were resolved by performing SDS-polyacrylamide gel electrophoresis and transferred onto nitrocellulose membranes (Millipore, Bedford, MA, USA). The membranes were blocked with 5% nonfat milk powder and 0.1% Tween-20 in tris-buffered saline (TBS) and incubated overnight with the specific primary antibodies at 4°C. Next, the membranes were washed three times with TBS containing Tween-20 (TBST) and incubated with the HRP-conjugated secondary antibody (GE Healthcare, Little Chalfont, Buckinghamshire, United Kingdom). Subsequently, the membranes were washed extensively with TBST, and reactive signals were detected when enhanced chemiluminescence western blotting detection reagents (Amersham Pharmacia Biotech, United Kingdom) were used. β-Actin was used as an internal control. Band intensities were detected by exposing the membranes to LAS-3000 (Fujifilm, Tokyo, Japan). Protein levels were determined by performing densitometric analysis with Multi Gauge, V2.2, software (Fujifilm).
Statistical analysis
Data were analyzed by using an unpaired t-test and expressed as mean ± standard deviation. P < .05 was considered statistically significant.
Results
IA inhibits the cell viability of B16-F1 melanoma cells
To clarify the effect of IA on melanoma tumor cells, we treated B16-F1 melanoma cells with different concentrations of IA. We seeded 5 × 105 B16-F1 melanoma cells per 60-mm dish. After 18 h, it was seen that the cells have reached a density of about 35–40% as IA was added in concentrations of 0, 200, 400, and 600 ppm, respectively.
As seen in Figure 2, 24 h after being treated with IA, there is a significant effect on the number of B16-F1 cells. The B16-F1 cell number decreases when the concentrations of IA are increased. IA at a concentration of 600 ppm caused a decline in the B16-F1 cell number by ∼40–50%. The microscopic view of cells demonstrated that there is not much cell debris floating when the concentration of IA is 600 ppm and the shape of cells becomes elongated and deformed (Fig. 2 upper).
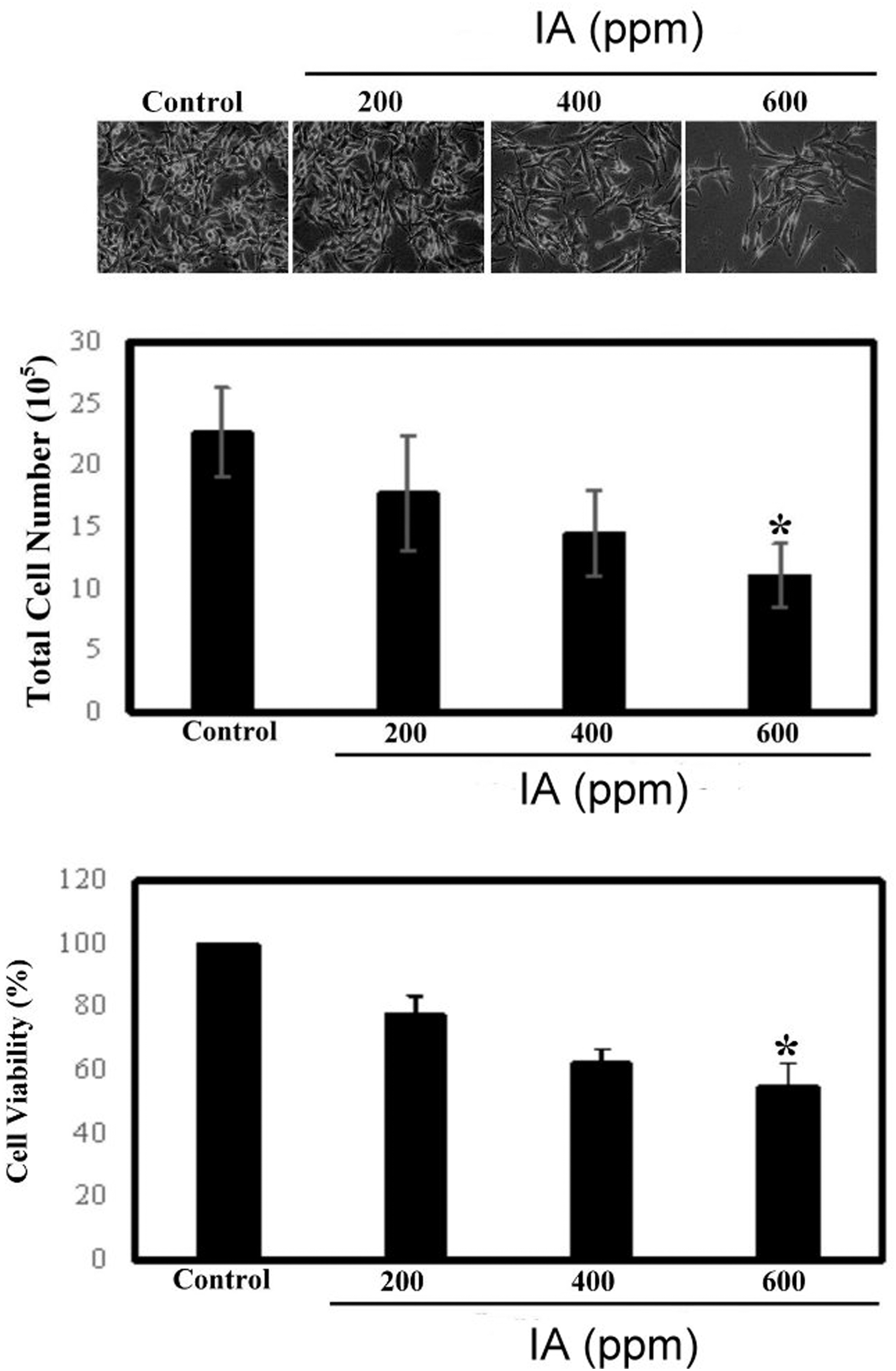
IA inhibits the growth and survival rates of B16-F1 melanoma cells. We treated B16-F1 cells with IA at concentrations of 0, 200, 400, and 600 ppm, respectively, for 24 h. Upper: Cell morphology detected by a microscope at a magnification of 100 × . Middle: Total cell number. The data are expressed as total cell number in every condition and presented as mean ± SD from three independent experiments. Lower: Cell viability was analyzed by MTT assay. The data are expressed as a percentage of the control (dose = 0) and presented as mean ± SD from three independent experiments. All the results were statistically analyzed by Student's t-test. *P < .01 compared with the untreated cell data. MTT, 3-(4, 5-dimethylthiazol-2-yl)-2, 5-diphenyltetrazolium bromide; SD, standard deviation.
To confirm whether the cell survival rate is reduced or not, we used the MTT assay to determine the proportion of cell viability. During the detection process, cells were treated in the same conditions. MTT assay was performed after the cells were treated with IA for 24 h. In Figure 2 lower, the MTT assay data show that the cell survival rate was significantly decreased by IA. It was reported that at a concentration of 600 ppm of IA, the cell viability is about 40–50%. This result is consistent with the trend of decreasing cell number.
Effects of IA on p-RSK and p-MSK1 signaling pathways in B16-F1 cells
Studies have shown that the role of the Ras/Raf/MEK/ERK pathway had a strong correlation with proliferation and progression of melanoma and is a key mechanism in the treatment of this tumor. 25,26 Therefore, we have first used western blotting to determine its relationship with the regulation of molecule performance.
B16-F1 melanoma cells were collected in the same way as shown in Figure 2 and cell lysates were collected after 24 h for western blot measurement. In our present study, we checked the expression of MAPK signaling pathway molecules, MSK1, and p-protein kinase B (AKT). Figure 3 showed that pc-Raf, p-MEK1/2, p-MAPK, and p-90RSK are all activated, but p-MSK1 and p-AKT showed a decline in activity. Our data indicated that p-RSK and p-MSK1/p-AKT are opposite in managing their activation, and this is also reflected in the research, as the activators regulated by these two are different. 8,10 The p-ERK1/2 signaling pathway (MAPK) regulates cell growth, proliferation, and differentiation by stimulation of growth factors, but p-MSK1 is activated by stimulation caused due to cellular stress and hypoxia, etc. These two molecules maintain coordination for survival of cells. Activation of MSK1 guides cells during autophagy. 9 Our data also showed that MSK1 is activated at concentrations of 200 and 400 ppm of IA, but demonstrates a weak response at a concentration of 600 ppm. This implies that the cells first promotes autophagy by raising MSK1, and later on a rise in the activity of MEK(MAP2K1, Dual specificity mitogen-activated protein kinase kinase 1)/RSK initiates the mechanism by which melanoma cells protect themselves from apoptosis. However, more investigation needs to be conducted to confirm these findings.
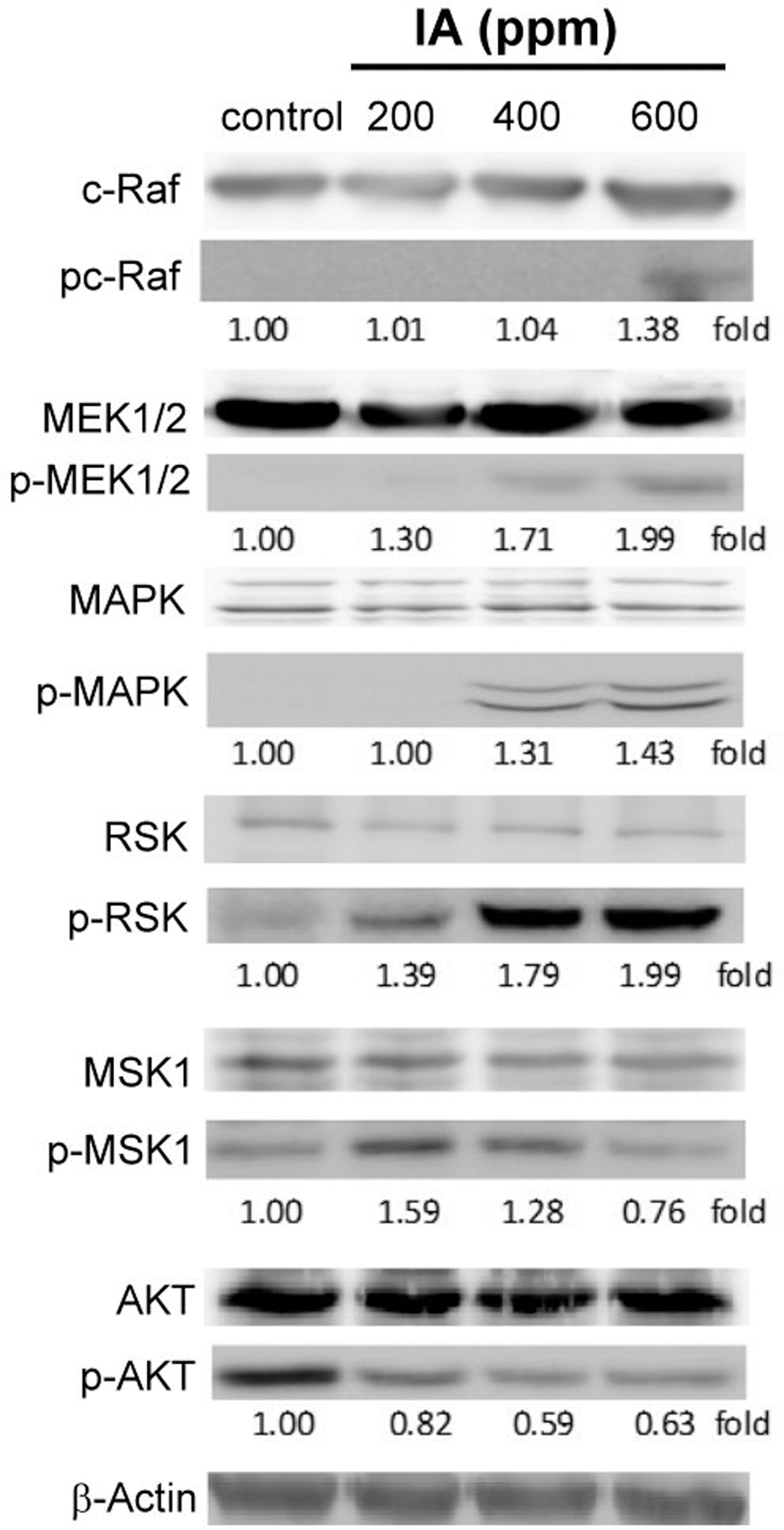
Western blotting was performed to detect the proliferation-related cell signaling molecule regulation in IA-treated B16-F1 melanoma cells. B16-F1 cells were treated with IA at concentrations of 0, 200, 400, and 600 ppm, respectively, for 24 h. The cell extracts were prepared as described in the Materials and Methods section. Equal amounts (40 μg) of total protein were loaded in each lane of SDS-polyacrylamide gel. Western hybridization was performed with antibodies against pc-Raf, p-MEK1/2, p-MAPK, p-90RSK, p-MSK1, p-AKT, and β-actin. Western blot analysis of β-actin was used as an internal control. All data are presented as the fold relative to the control. Three independent experiments were conducted, all of them showed similar patterns of change. AKT, protein kinase B; MAPK, mitogen-activated protein kinase; MEK, MAP2K1, dual specificity mitogen-activated protein kinase kinase 1; MSK, mitogen- and stress-activated protein kinase; RSK, 90 kDa ribosomal S6 kinase; SDS, sodium dodecyl sulfate.
IA inhibits motility and MMP9 expression in B16-F1 melanoma cells
Since IA inhibits the viability of B16-F1 melanoma cells, we also tested cell motility. A previous review has shown that ERK1/2, focal adhesion kinase (FAK), and PI3K/mTOR affect the ability of cells to undergo motility and angiogenesis. 27 We found that treatment with IA upregulated the expression of MEK and ERK1/2, whereas it downregulated the expression of MSK1. Whether this outcome is related to cell motility was also checked. First, B16-F1 cells were treated with different concentrations of IA (0, 200, 400, and 600 ppm, respectively) for 24 h. Then, we used the wound healing assay to detect the cell migration ability. It was evident (as shown in Fig. 4a) through the wound healing assay that B16-F1 cell mobility was drastically reduced when IA concentration was increased. Regarding the activity of signaling molecules, the results confirmed that FAK activity was downregulated (Fig. 4b).
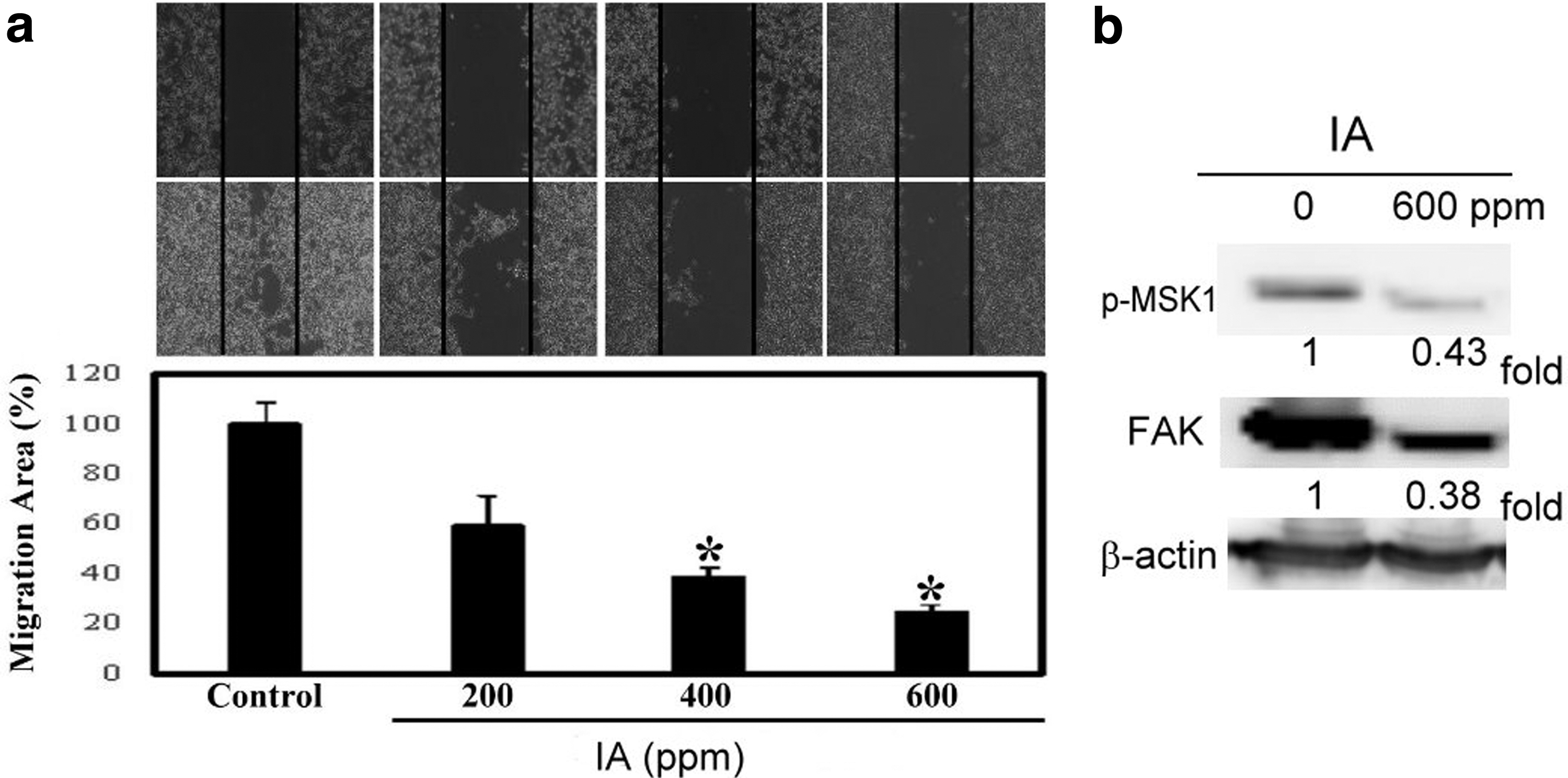
Effect of IA on inhibition of cell mobility in B16-F1 cells.
IA induced autophagy in B16-F1 melanoma cells
Studies have pointed out that melanoma tumor cells easily induce autophagy when stimulated by stress or chemotherapy drugs. 28 Therefore, we treated B16-F1 cells with IA to investigate its autophagy-related measurements. We first identified the cells involved in autophagy by AO staining. The results show that when B16-F1 cells are stimulated with IA at a concentration of 600 ppm, 59% of AVO-positive cells are detected (Fig. 5). Therefore, it is clear that the effect of IA on B16-F1 cells may cause cell death through the autophagy pathway. The AO-positive cells are then observed under a fluorescence microscope (Fig. 5 upper). The status of cell death due to apoptosis was evaluated by 4′,6-diamino-2-phenylindole staining, but no difference in the outcome was noticed after treatment with IA (data not shown).
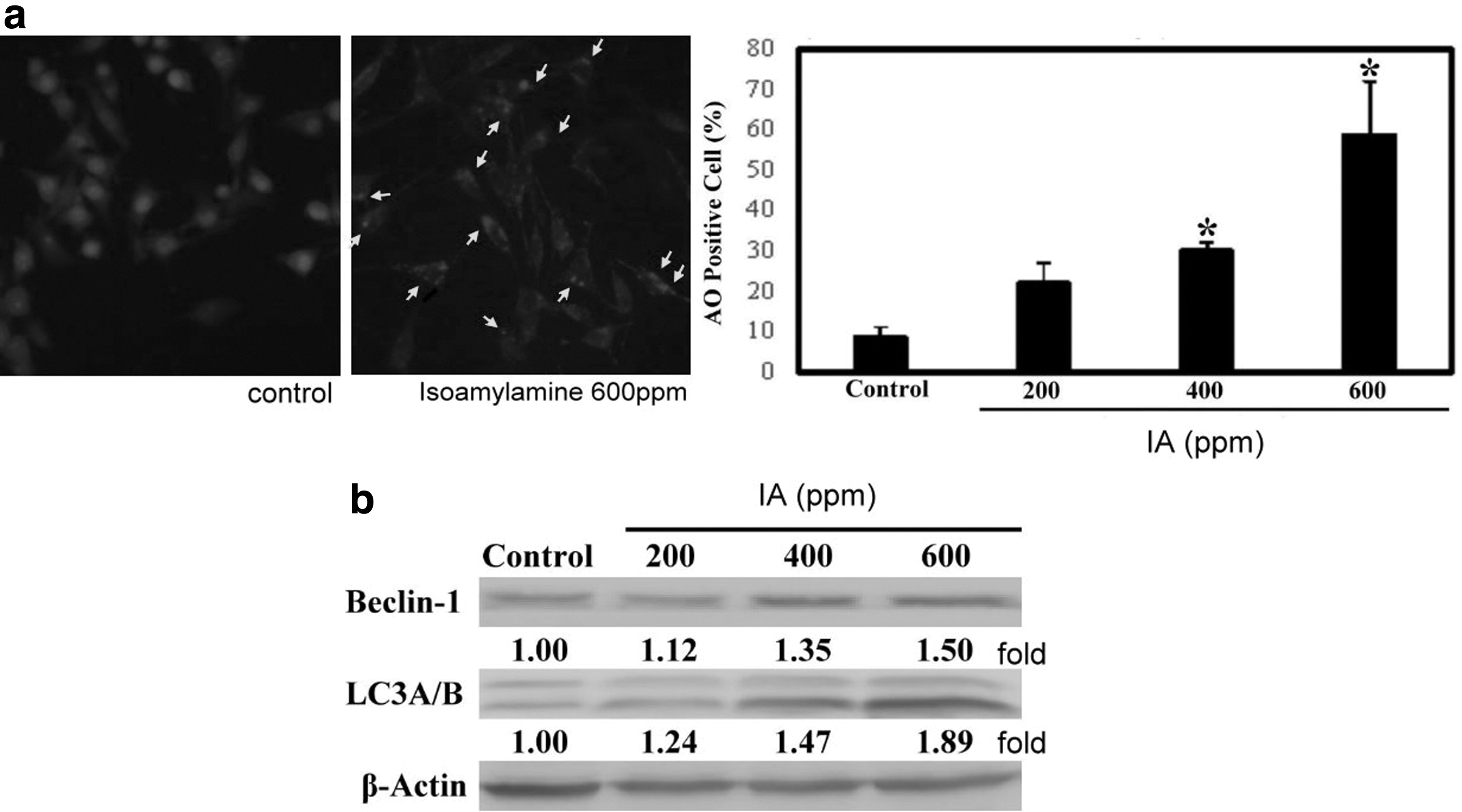
Autophagy induced by IA in B16-F1 melanoma cells. B16-F1 cells were treated with IA at concentrations of 0, 200, 400, and 600 ppm, respectively, for 24 h and later subjected to autophagy detection.
To further confirm the autophagy pathway associated with IA treatment, we have also investigated regulatory proteins related to autophagy. Therefore, two autophagy proteins, namely BECN-1 and LC3A/B, were used for detecting the process. The western blot findings demonstrated that both BECN-1 and LC3A/B were upregulated (Fig. 5b).
The LKB1/p-AMPKα/FASN pathway was regulated in IA-induced autophagy seen in B16-F1 cells
Previous research has shown that melanoma cells enhance the pathway linked with Ras to ERK1/2 through BRAF, resulting in inhibition of LKB1 activity. It is documented that this leads to separation from AMPK and cells get the advantage of proliferation. The studies also mentioned that if LKB1 and AMPK come together to promote AMPK phosphorylation, it will cause inhibition in the growth and proliferation of cells. 22 AMPK activation induces cell death by autophagy through FASN, BECN-1, and LC3, which also correlates with tumor suppression. 29,30 Thus, we checked the autophagy-related molecules by western immunoblotting. The data highlighted that the level of p-AMPKα was increased, while a decrease was noticed in FASN (Fig. 6a). This response is similar to that seen during the autophagy mechanism. Since p-AMPKα is correlated with autophagy flux, we planned to use dorsomorphin (AMPK inhibitor) to block its activity. Therefore, we used 5 μM of dorsomorphin to treat B16-F1 melanoma cells for 30 min, then 600 ppm of IA was added as planned in the original design. The western blot data showed (Fig. 6b) that in the absence of dorsomorphin, IA induced p-AMPKα activity and led to a rise in its level. BECN-1 and LC3A/B also increased, while FASN showed a reduction in its level, which correlates with results seen in previous studies. However, when the dorsomorphin-pretreated group was observed, it was seen that the rise in p-AMPKα level and decline of FASN became less insignificant, and even the rise associated with the autophagy markers (BECN-1 and LC3A/B) became weak.
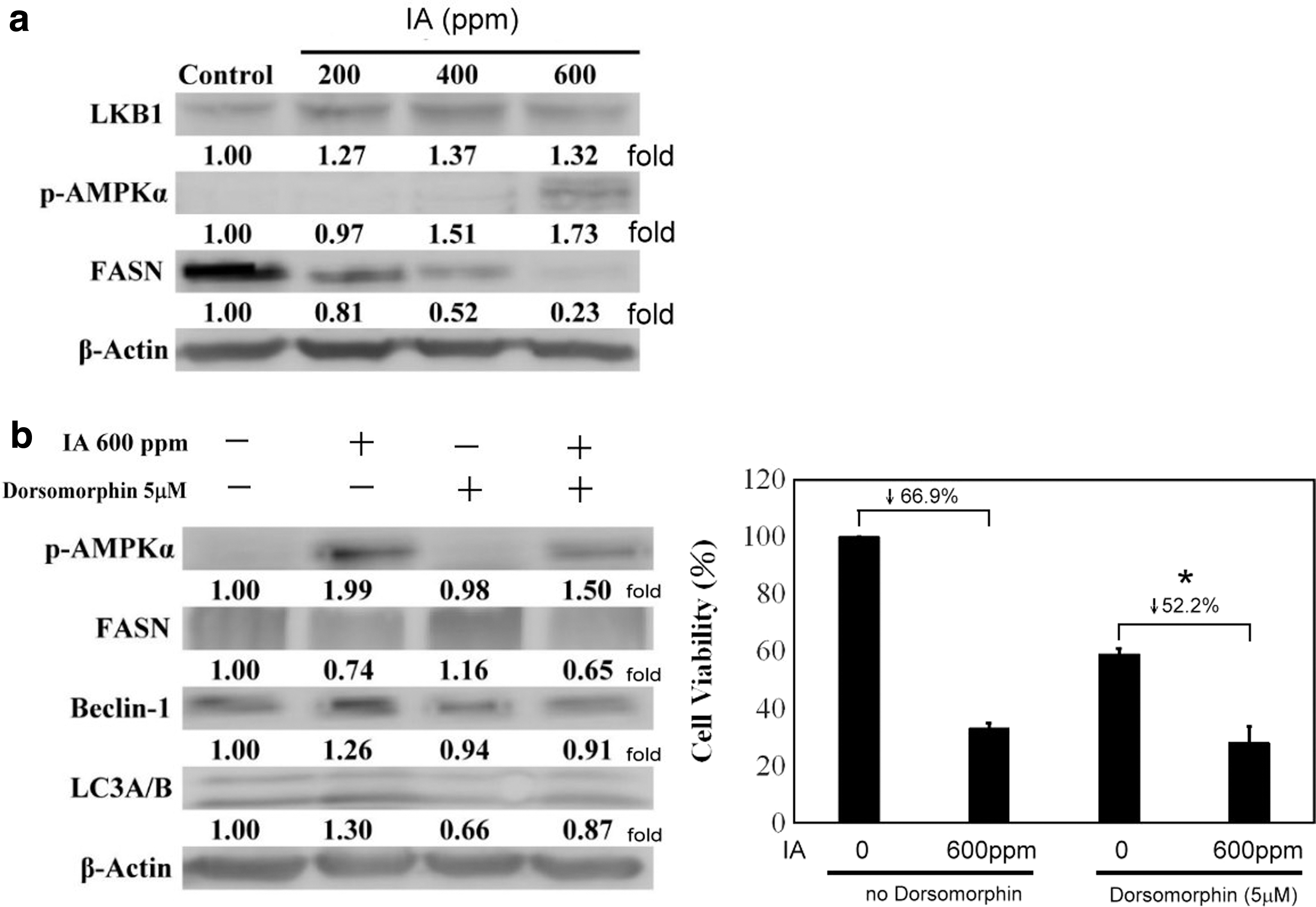
IA has an effect on activation of the AMPK/autophagy pathway in relation to B16-F1 melanoma cells.
The cell survival rate, as shown by MTT assay (Fig. 6c), demonstrated that IA at a concentration of 600 ppm inhibited about 66.9% of B16-F1 cell viability and also inhibited AMPK in the absence of dorsomorphin. On the other hand, results showed that when dorsomorphin is used to suppress AMPK, the survival rate can be improved (suppressed only 52.2%). The present study confirmed that 5 μM of dorsomorphin can promote the viability of B16-F1 cells by about 22% and this is statistically significant. Therefore, in melanoma cells, AMPK is one of the most important factors involved in the process of autophagy induced by IA.
In summary, our data indicated that IA induces autophagy through the AMPK pathway. Upstream signaling included activation of Raf/MEK/ERK, while MSK1 and AKT were suppressed. B16-F1 melanoma cell mobility was also decreased, and Matrix Metalloproteinase-9 (MMP9) enzyme activity and FAK were downregulated when the treatment was conducted with IA.
Discussion
This is the first study that has indicated that IA can induce autophagy and is related to phosphorylation of AMPK in cells. This shows that IA and ODC have similar functions in regulating cell death and IA tends to cause cell growth restriction through autophagy in melanoma cells. There are not many studies that have focused on the physiologic effect of IA. According to a study conducted by Yen et al., it was demonstrated that IA improves the mortality rate of mice suffering from endotoxemic shock. It is reported that IA, at a concentration of 1000 ppm, ameliorates lipopolysaccharides-induced damage noticed during extrinsic (prothrombin time) pathways and during MMP9 expression. Through activation of ERK/p-P38 in liver cells, MMP9 secretion is reduced. 31 A previous study has also shown that activation of MSK1 induces MMP9 expression. 10 In our study, we also used the gelatinase zymography assay to detect the expression of MMP, and results pointed out that MMP-9 expression tends to decline, whereas MMP-2 response is not clearly known (data not shown). This outcome is consistent with the above observation, but the regulation aspect is still unclear. It is seen that IA also plays a significant role in maintaining heart function. 32 Studies have shown that when endogenous IA is elevated, the septal wall in the heart is thinner. In addition, the higher the level of ODC, the larger is the size of the left atrium. The findings have confirmed that the concentration of IA is directly related to the probability of encountering congestive heart failure. This phenomenon is possibly caused by the IA-induced autophagy effect. It is another important issue that needs to be explored.
A study indicated that AZs affect cell proliferation and viability through regulating cellular polyamines. 6 AZs can combine with ODC and reduce ODC dimer, eventually affecting the uptake and excretion of cellular polyamines. When the cellular polyamine level is increased, its cytotoxicity leads to cell death. However, if the level of polyamines is too low, it causes cell growth inhibition. 33,34 Therefore, it is important for cells to fine-tune the polyamine level, which is useful for demonstrating the antitumor effect. Our results provide additional evidence that IA can also cause cell death by regulating autophagy and is also related to antiproliferation/antitumor activity. During a study, Tobias et al. exposed mouse myeloma cells to the ODC overproducing condition, also treating them with ornithine, to result in a massive conversion of ornithine to putrescine. They detected that 213 nmol/mg of putrescine had an apoptotic cell death effect. In our present study, the IA concentration was 600 ppm, which is almost 6.9 μM/mg and higher compared with the above study. Although the types of polyamines and cells involved are different in both the studies, there is a similar tendency with regard to cell death, as induced by polyamine overdose.
In association with the autophagy regulator molecule, AMPK, degradation of it by a cancer-specific ubiquitin can downregulate an oncogenic driver seen in cancer. 23 From a different perspective, autophagy can also be considered as a protective or functional cell death mechanism. 35 Our results have indicated that B16-F1 melanoma cells may have a dual mechanism to escape or overcome cellular stress. As seen in the study, there is an inclination to suppress MSK1 and then induce autophagy. On the other side, ERK/RSK was activated and might be an important factor for melanoma cells to survive under stress. Previous studies have shown that autophagy can not only induce cell death and is one of the cancer treatment options but it is also regarded as a prosurvival mechanism that causes resistance during chemotherapy and can interact with the process of apoptosis. 36 In a panduratin A-induced autophagy study, mTOR was suppressed and p-AMPK was activated. However, when autophagy was inhibited by 3MA, it caused cells to undergo apoptosis. 37 There is evidence indicating that autophagy and apoptosis are two types of cell death mechanisms that are interchangeable. When autophagy was suppressed, cells tend to turn to apoptosis. 18,36 Whether inhibition of AMPK by dorsomorphin in B16-F1 cells causes cells to transfer to apoptotic cell death and what is the role played by ERK/RSK in this process are still unclear. These will need further validation in the future.
A study involving GBM showed that MSK1 was activated by ERK and cell proliferation was also detected. 10 Marino et al. showed that acidic stress in melanoma cells induced protective autophagy by activation of ERK and AKT. 35 When cardiac myocytes were involved, MSK1 was activated and showed cardioprotective effects. In our study, ERK was activated, while MSK1 was suppressed, which differs from the above studies. Whether ERK and MSK1 activation occurs simultaneously or is regulated by other signaling molecules is an essential factor for understanding the mechanism behind protective autophagy. We need to conduct more investigations to prove it. There might be other regulatory mechanisms operating to control this result because it was observed in our study that 800 and 1000 ppm of IA will still cause severe cell death (data not shown).
In summary, IA leads to a decrease in the viability and mobility of B16-F1 melanoma cells and induces autophagy by regulating AMPK. The study elucidated the efficacy of IA in demonstrating melanoma tumor cell biology. It is valuable for understanding the mechanism of melanoma cells during chemoprevention and this will help during the course of prognosis in the future.
Footnotes
Authors' Contributions
Conceptualization was developed by T.-P.Y., S.-R.W., and Y.-C.P.; data curation was performed by T.-P.Y., Y.-F.L., and K.-L.L.; funding acquisition was done by Y.-C.P.; experiment execution was performed by Y.-F.L. and S.-J.L.; supervision and validation were performed by T.-P.Y.; writing—original draft—was done by T.-P.Y. and N.-M.T.; and writing—review and editing—was done by T.-P.Y. and S.-R.W. All authors have read and agreed to the published version of the manuscript.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
We thank the Taichung Veterans General Hospital Research Program for funding this work, grant numbers are TCVGH1083302C and TCVGH1073302C.