
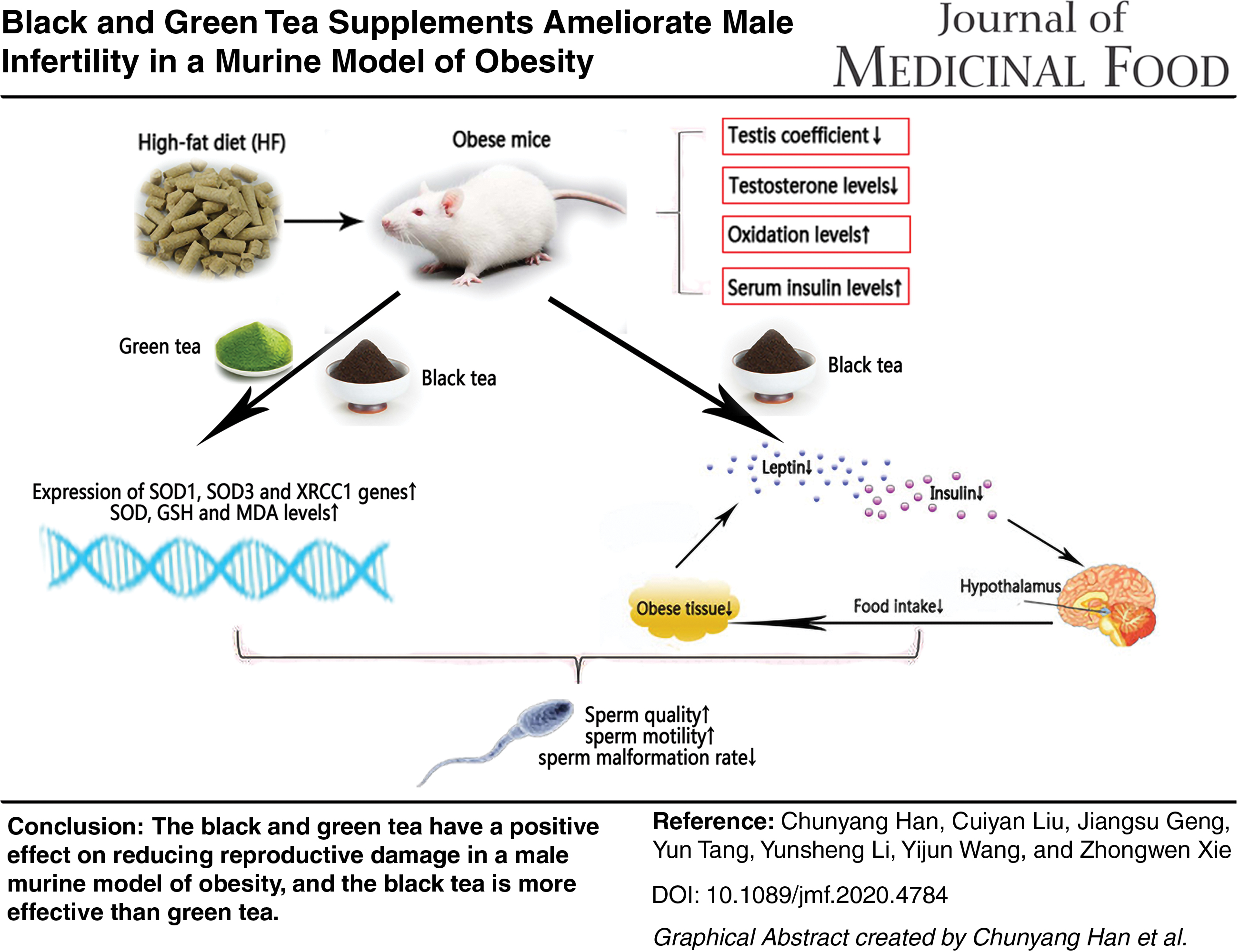
Abstract
Obesity, a chronic metabolic disorder, can affect male reproductive function. As a functional beverage, tea has many biological activities and potential in the treatment of obesity. However, its effects on male reproductive damage induced by obesity are not yet clear. In this study, a murine model of obesity was established by feeding with high-fat diet (HF). A total of 24 male mice were divided into four groups: normal diet (control), HF, HF supplemented with 5% green tea powder (HF+G), and HF supplemented with 5% black tea powder (HF+B). The results showed that the HF + B significantly reduced the mouse body weight gain and testicular coefficient and lowered the serum insulin and leptin levels compared with the HF group. The sperm malformation rate of mice in the HF group had a significant increase when compared with the control group, the HF + B group had a significant decrease compared with the HF group, and no difference from the control group. The HF + G and HF + B significantly increased testosterone levels in serum compared with the HF group. The testosterone production-related gene cytochrome P450 family 11 subfamily a member (CYP11A1) and cytochrome p450 family 17 subfamily a member 1 (CYP17A1) expressions in testis were significantly increased in the HF + G group compared with HF group. In addition, the HF + G and HF + B abolished the effects of HF on superoxide dismutase (SOD), malondialdehyde, and glutathione levels in testis and antioxidant-related gene expressions of XRCC1 and SOD1. Overall, our findings have provided evidence that black and green tea has a positive effect on reducing reproductive damage in a male murine model of obesity, and that black tea is more effective than green tea.
Color images are available online
Introduction
The prevalence of obesity has markedly increased over the past three decades and is now a major international public health challenge. It arises from the excessive accumulation of fat, the dysregulation of energy balance due to excessive energy intake, and insufficient energy expenditure. Obesity is considered to be a significant risk factor for various chronic diseases, such as diabetes, cardiovascular disease, and cancer. 1 –3
Obesity, it can also cause the occurrence of reproductive disorders. Obesity reduces the quality and quantity of eggs and increases the rate of miscarriage during pregnancy. 4 –6 Obesity reduces oocyte quality by affecting meiosis, disrupting spindle assembly, and promoting oxidative stress and abnormal mitochondrial distribution. 7 For males, obesity has a severe impact on the development and function of the testicles, epididymis, prostate, and other male reproductive organs. 8 With the increase of body mass index and abdominal circumference, ejaculation volume gradually reduces, and the total sperm count in semen also decreases. 9 In addition, obesity damages sperm chromatin 10 and further increases miscarriage risk. 11 Weight loss can increase testosterone levels in males, which is closely related to the decreased serum concentration of insulin and leptin, effectively reducing the risk of infertility.
As a functional beverage, tea has known benefits for improving diseases caused by obesity. 12 The main components of tea, such as tea polyphenols and tea polysaccharides, can regulate protein kinase (AMPK) to enhance the phosphorylation of the AMPK signaling pathway to turn white adipose tissue into brown adipose tissue, which can reduce body weight (BW). 13
Green tea mainly inhibits digestive enzyme activity and nutrient absorption, leading to the reduction of actual intake of energy, thus leading to weight loss 12,14 At the same time, green tea can also reduce serum cholesterol, triglyceride, and adiponectin levels in obese mice, thus preventing the adverse effects of obesity. Black tea is a fully fermented tea, and the extract of black tea can inhibit the absorption of intestinal fat induced by diet in rats, thus achieving the goal of weight loss. 15,16 Black tea can also regulate postprandial blood glucose, reduce lipid absorption, and decrease blood glucose and lipid levels. 17 Overall, both black tea and green tea have a certain effect on weight loss, but whether tea can have beneficial effects on obesity-induced male reproductive diseases in mice has not been reported.
In view of the above, the aim of this study was to explore the effect of green and black tea on male reproduction organ damage induced by a high-fat diet (HF) in mice. Our study found that black tea can reduce the rate of sperm malformation and improve obesity-related male reproductive disorders by regulating the change in hormone levels and oxidative stress. Our current study may provide a research basis for the effect of tea on male reproduction.
Materials and Methods
Tea samples
The green tea and black tea used in this study were produced as previously described. 18 Fresh leaves of Longjing 43 (Camellia sinensis) were plucked from number 916 tea plant breeding garden in Shucheng, Anhui, China. All of the fresh leaves were divided into two equal portions and processed into green tea and black tea using typical manufacturing approaches. Briefly, half of the fresh leaves were first fixed at 220°C to terminate the endogenous enzymatic reaction, then rolled for 30 min, and then being directly dried into green tea. The other half of the leaves were withered at room temperature and 70% humidity for 5 h, then being rolled for 30 min, applied heat-moisture treatment at room temperature and 90% humidity for 3 h, and immediately dried into black tea.
Experimental design and dietary treatment
Twenty-four 10-week-old male C57BL/6J mice weighing 17–21 g were purchased from Cavens Experimental Animal Co., Ltd. (Changzhou, Jiangsu, China), license number SCXK (su) 2011–0003, and kept in the animal feeding center of Anhui Agricultural University. Animals were used and handled in accordance with the regulations of Anhui Agricultural University on the management of experimental animals and approved by the Anhui Agriculture University Committee on Animal Care and Use (approval no. AHAU2018028). The mice were housed in a controlled environment at 23°C ± 2°C, with 60% ± 5% humidity, a 12-h light/dark cycle, and provided free access to water.
After those healthy mice were acclimatized for 1 week, they were then randomly divided into four groups (n = 6 per group), caged separately. From the beginning of the experiment, they were fed with a normal diet (control, 10% fat calories), HF (45% fat calories, Luofei company, Jiangsu, China), HF with 5% green tea powder (HF+G), or HF with 5% black tea powder (HF+B) for 18 consecutive weeks.
BW was measured weekly throughout the experiment. At the end of the experiment, all the mice were anesthetized by intraperitoneal injection of 50 mg/kg of 2.0% pentobarbital sodium, then weighed, and blood was collected from the mice orbit for serum testosterone, insulin, and leptin level assay. After killing the mice by cervical dislocation, the testes were weighed on both sides. The right testis was stored at −80°C for detection of the oxidation index and gene expression, and the left testis was fixed in 4% paraformaldehyde solution for hematoxylin and eosin (H&E) tests. The testicular coefficient was calculated as (testicular weight/BW) × 100.
Sperm quality assay
The sperm quality testing was performed according to a previous publication. 19 To obtain mature sperm, both the epididymides and vasa deferentia were removed. Spermatozoa contained in each vas deferens were drawn to the epididymis by using a pair of forceps. Then, vasa deferentia were cut off and epididymides were placed in a 35 mm plastic culture dish containing HEPES-buffered modified Tyrode's medium (T1788; Sigma, St Louis, MO, USA). The dishes were kept at 37°C on a warm plate during the whole procedure.
Sperm motility was evaluated by placing 8 μL of the sperm suspension between a prewarmed slide and a 22 × 22 mm coverslip, and examining it at 100 × magnification under phase-contrast optics. The percentage of motile sperm (ranging between 0%, when no motile spermatozoa were observed, and 100%, when all spermatozoa were moving) was estimated subjectively by at least two independent experienced observers; estimations from the different observers were averaged and rounded to the nearest 5% value.
For the abnormal spermatozoa test, 10 μL of the diluted seminal fluid was smeared onto a glass slide, fixed with SpermBlue fixation solution (Microptic SL, Barcelona, Spain) for 15 min, and then stained with SpermBlue staining solution for 10 min. After drying, 200 spermatozoa were observed and counted under an optical microscope, the number of abnormal spermatozoa was counted, and the rate of abnormal spermatozoa was calculated.
Histological examination
The right testes were fixed with 4% formaldehyde for 24 h and embedded to prepare paraffin sections according to conventional histological methods. H&E staining was performed to observe the morphological changes of the testes under a microscope (IX71, Olympus, Japan). All stained slides were analyzed in a blinded manner by an expert pathologist.
Serum parameter assay
The serum insulin (EZRMI-13K; Millipore, Burlington, MA, MA, USA), leptin (EZML-82K; Millipore), and testosterone (KGE010; R&D Systems Minneapolis, MN, USA) concentrations were evaluated using the ELISA kits according to the kit manufacturer's instructions.
Detection of oxidative damage index of testicular
The superoxide dismutase (SOD), malondialdehyde (MDA), and glutathione (GSH) levels in testicular tissue were evaluated using the kits (Jiancheng Biological Engineering Institute, Nanjing, China). SOD levels were detected using the hydroxylamine method, MDA levels were detected by the thiobarbituric acid method, and GSH levels were detected by colorimetry. All assays were performed according to the kit manufacturer's instructions.
Quantitative reverse transcription polymerase chain reaction
Total RNA was extracted from testis using the TRIzol method. RNA was reverse transcribed into cDNA using a reverse transcription kit (Vazyme Biotech, Nanjing, China). Forward and reverse primers directed toward apurinic/apyrimidinic endonuclease 1 (Apex1), SOD1, SOD2, SOD3, X-Ray repair cross complementing 1 (Xrcc1), 8-oxoguanine DNA glycosylase (Ogg1), cytochrome P450 family 11 subfamily a member 1 (Cyp11a1), cytochrome p450 family 17 subfamily a member 1 (Cyp17a1) and cytochrome P450 family 19 subfamily a member 1 (Cyp19a1), and the housekeeping gene β-actin are listed in Table 1.
Primer Sequences for Quantitative Reverse Transcription Polymerase Chain Reaction
FastStart SYBR Green Master Mix (Vazyme Biotech) was used for quantitative polymerase chain reaction (qPCR). The amplification reaction included template denaturation and polymerase activation at 95°C for 30 sec, followed by 35 cycles of denaturation at 95°C for 15 sec, annealing at 60°C for 30 sec, and extension at 72°C for 30 sec. Melt curve analyses were performed for all genes, and the specificity, as well as integrity, of the PCR products was confirmed by the presence of a single peak.
Data analysis
All experiments were carried out at least three times. All the data were analyzed using one-way analysis of variance (SPSS 19.0) and presented as mean ± standard error of the mean. A P value of less than .05 was considered to be statistically significant.
Results
BW, physiological index of testes, and insulin and leptin levels
The mice of the HF group exhibited a rapid BW gain from the first week of the feeding period in comparison with those of the control group (Fig. 1A). After 18 weeks of feeding, the BW of the HF group significantly increased compared with the control group (40.50 ± 2.35 vs. 24.55 ± 1.00 g, P < .01). The HF + G group (36.58 ± 0.83 g) was significantly higher than that of the control and HF + B group (32.54 ± 0.83 and 24.55 ± 1.00 g, P < .05). However, no difference was found between the HF and HF + G groups (40.50 ± 2.35 vs. 36.58 ± 0.83 g, P > .05) (Fig. 1B).

Effects of a normal (control), HF, and HF supplemented with green tea (HF+G) and black tea (HF+B) diet on
Next, we measured testicular weight and testicular coefficient. Testis weight showed no significant differences among the four groups (HF, 0.23 ± 0.01 g vs. the control, 0.20 ± 0.01 g vs. HF+G, 0.23 ± 0.02 g vs. HF+B, and 0.23 ± 0.02 g, P > .05) (Fig. 1C). However, the testis coefficient of the HF (5.72 ± 0.35) and HF + G groups (6.46 ± 0.36) was significantly lower than the control group (7.81 ± 0.53) (P < .05) (Fig. 1D). There were no significant differences among the HF+G, HF+B, and HF groups using the testis coefficient (6.46 ± 0.36, 7.05 ± 0.45, vs. 5.72 ± 0.35, P > .05).
The serum insulin levels in the HF (3.80 ± 0.68 ng/mL) and HF + G groups (4.60 ± 1.07 ng/mL) were significantly higher than that of the control (0.57 ± 0.22 ng/mL) (P < .01) and HF + B groups (1.48 ± 0.50 ng/mL) (P < .05) (Fig. 1E). There was no significant difference between HF + G and HF groups for serum insulin levels (3.80 ± 0.68 vs. 4.60 ± 1.07 ng/mL, P > .05). Leptin in HF group (29.10 ± 8.25 ng/mL) was significantly higher than that in the control (1.50 ± 0.58 ng/mL) (P < .01), HF + G (6.34 ± 1.52 ng/mL) (P < .05), and HF + B groups (1.72 ± 0.85 ng/mL) (P < .01) (Fig. 1F). Leptin in HF + G group (6.34 ± 1.52 ng/mL) was significantly higher than that in the control (1.50 ± 0.58 ng/mL) (P < .05) and HF + B groups (1.72 ± 0.85 ng/mL) (P < .05) (Fig. 1F). There were no significant differences between the control (1.50 ± 0.58 ng/mL) and HF + B groups (1.72 ± 0.85 ng/mL) (P > .05) (Fig. 1F).
Effects of black and green tea on sperm quality and the histology
The sperm motility and malformation rate were analyzed. Compared with the HF group (72.50% ± 10.41%), the sperm motility in the HF + G (85.00% ± 6.78%) and HF + B (87.50% ± 8.53%) groups was significantly increased (P < .05) (Fig. 2A), but there were no significant differences among the control (81.00% ± 6.52%), HF + B (87.50% ± 8.53%), and HF + G groups (85.00% ± 6.78%) (P > .05) (Fig. 2A). Compared with the control group (11.94% ± 2.53%), the sperm malformation rate in the HF group (16.54% ± 1.23%) had a significant increase (P < .05) (Fig. 2B), there were no significant differences among the HF+G, HF, and control groups (13.24% ± 2.87%, 16.54% ± 1.23%, vs. 11.94% ± 2.53%, P > .05) (Fig. 2B), but the HF + B group (11.57% ± 3.22%) had a significant decrease compared with the HF group (16.54% ± 1.23%) (P < .01) (Fig. 2B).
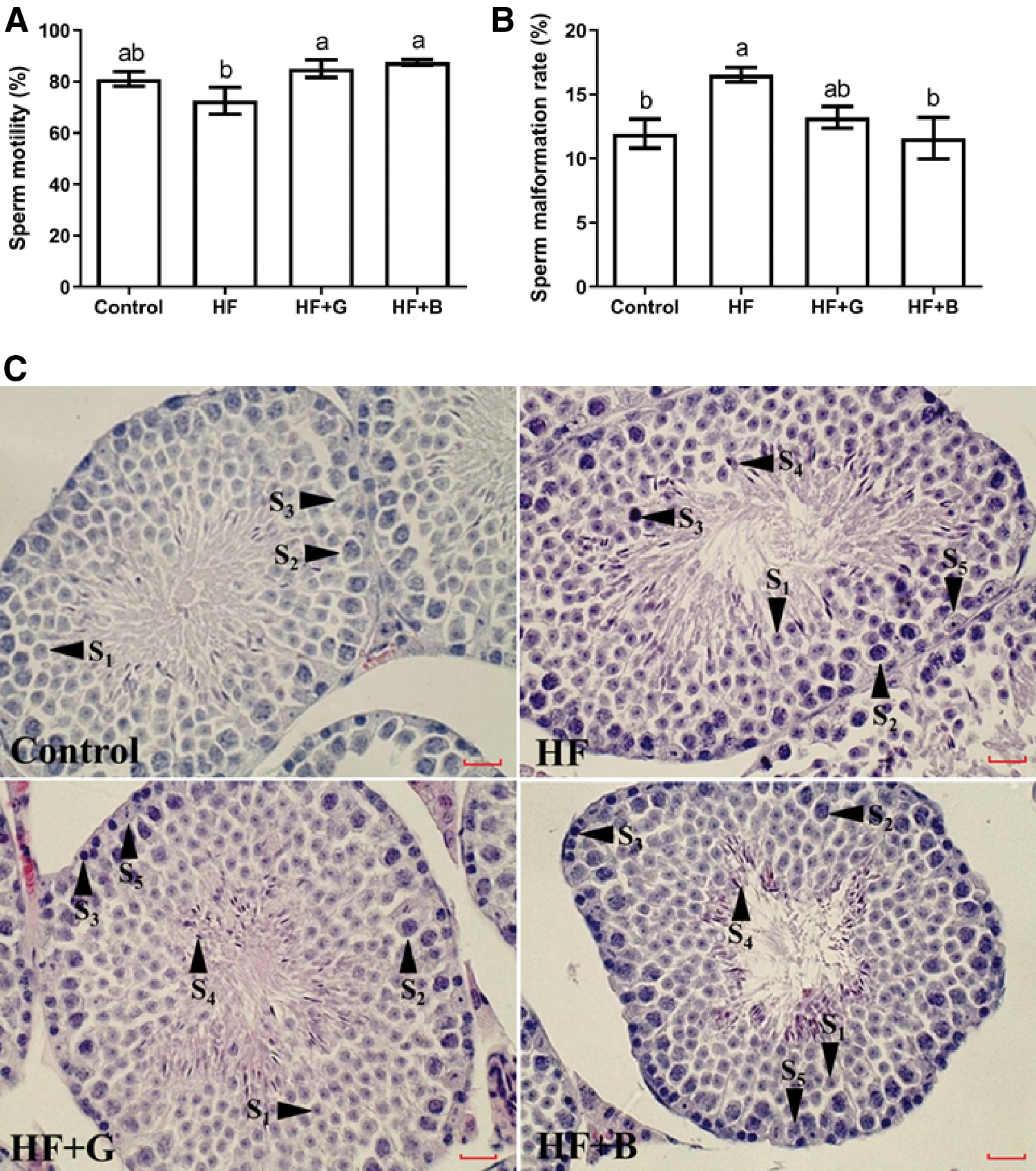
Effects of black and green tea supplements on sperm quality and testicular histology in a murine model of obesity (n = 6).
The histological differences in testis were explored. As shown in Fig. 2C, sperm cells (primordial spermatocytes, spermatogonia, primary and secondary spermatogonial cells, spermatogonial cells) at different stages were observed in all of the four groups, with normal morphology and no obvious lesions.
Effects of black tea on testosterone levels and expression of testosterone synthesis-related genes
Obesity can inhibit the formation of spermatozoa in the testis by reducing the testosterone levels and increasing the oxidation levels, leading to a spermatogenic disorder and an increased sperm malformation rate. Therefore, we examined the testosterone levels in serum and related genes during steroid production.
Serum testosterone levels in the HF group (4.60 ± 1.07 ng/mL) were significantly reduced compared with the control (8.12 ± 0.57 ng/mL), HF + G (6.56 ± 0.41 ng/mL), and HF + B groups (8.00 ± 0.29 ng/mL) (P < .01) (Fig. 3A). Testosterone levels in HF + G group (6.56 ± 0.41 ng/mL) were significantly lower than the control (8.12 ± 0.57 ng/mL) and HF + B groups (8.00 ± 0.29 ng/mL) (P < .05) (Fig. 3A). There was no difference between the HF + B and HF group testosterone levels (8.00 ± 0.29 vs. 8.12 ± 0.57 ng/mL, P > .05) (Fig. 3A).
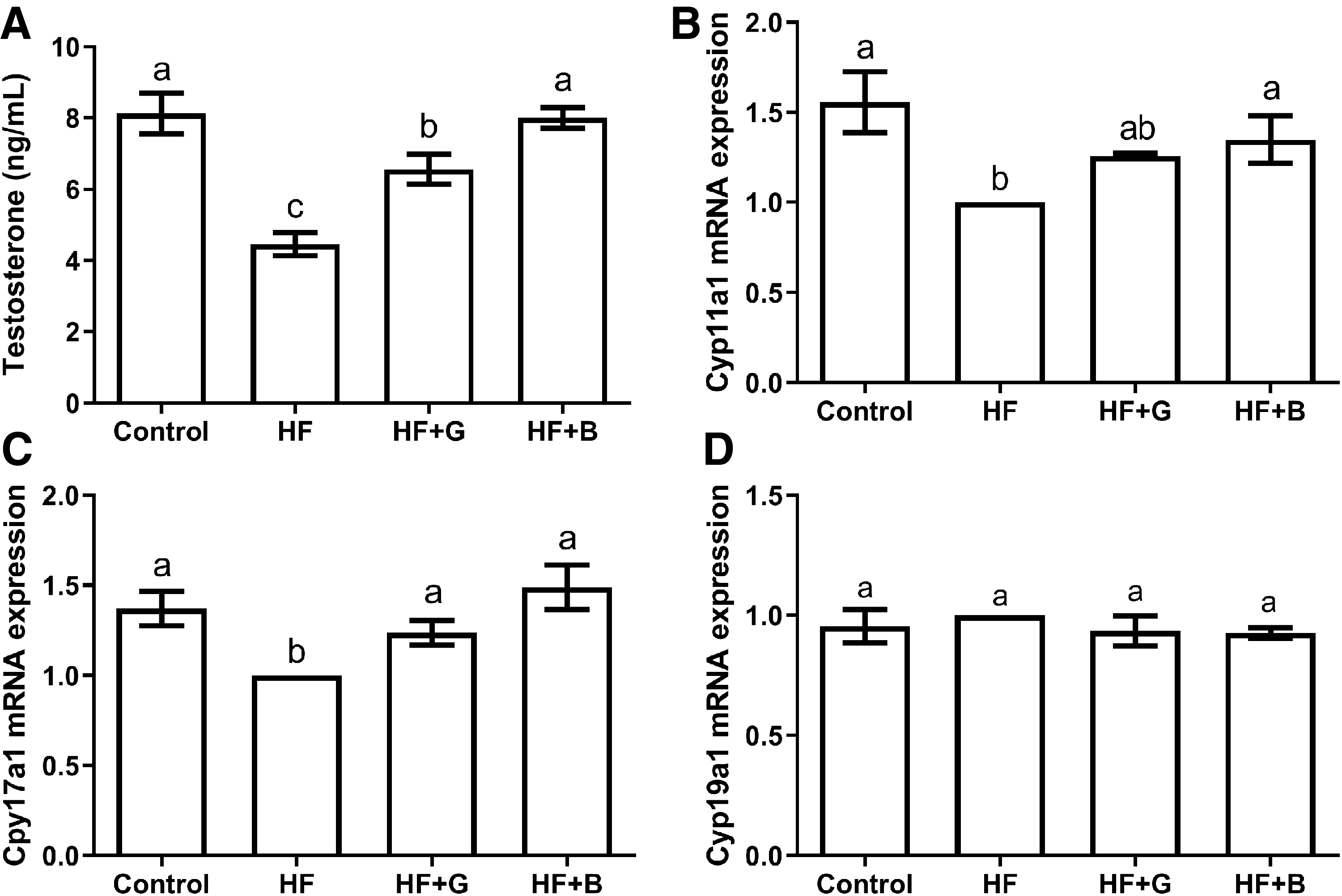
Effects of black and green tea supplements on testosterone levels and related gene expression of testosterone synthesis in a murine model of obesity (n = 6).
The expression level of the Cyp11a1 gene in the HF group was significantly lower than the control (P < .01) and HF + B groups (P < .05) (Fig. 3B). Furthermore, there was no difference in Cyp11a1 gene expression found between the HF + G and HF groups (P > .05) (Fig. 3B). The expression of Cyp17a1 gene in the HF testicular tissue was significantly lower than the control, HF + B (P < .01), and HF + G groups (P < .05) (Fig. 3C), and the differences among the HF+G, HF+B, and control groups were not significant (P > .05) (Fig. 3C). There were no significant differences in the expression levels of the Cyp19a1 gene among the four groups (P > .05) (Fig. 3D).
Effects of black tea on oxidative damage
Obesity can reduce the activity of free radical scavenging enzymes in cells, produce an excessive generation of reactive oxygen species (ROS) or imbalance the oxidation/antioxidant system, and lead to increased cell oxidation levels. We analyzed the antioxidant system changes in testis tissue. SOD levels in the HF group (25.43 ± 3.33 U/mgprot) were significantly lower than the control (38.62 ± 4.95 U/mgprot) and HF + B groups (39.88 ± 2.32 U/mgprot) (P < .05) (Fig. 4A). Moreover, there were no significant differences among the control, HF+G, and HF + B groups in SOD levels (38.62 ± 4.95, 36.90 ± 5.14, vs. 39.88 ± 2.32 U/mgprot, P > .05) (Fig. 4A).

Effects of black and green tea supplements on antioxidants in a murine model of obesity (n = 6).
The MDA level in the testis tissues of HF mice (1.53 ± 0.43 nmol/mgprot) was significantly higher than that of the control (0.47 ± 0.02 nmol/mgprot), HF + G (0.46 ± 0.10 nmol/mgprot), and HF + B groups (0.45 ± 0.05 nmol/mgprot) (P < .01). Also, there were no differences among the control, HF+G, and HF + B groups (0.47 ± 0.02, 0.46 ± 0.10, vs. 0.45 ± 0.05 nmol/mgprot, P > .05) (Fig. 4B).
The testis tissue GSH levels in the HF group (6.75 ± 2.10 U/mgprot) were significantly lower than in the control (13.40 ± 0.89 U/mgprot), HF + G (16.05 ± 1.67 U/mgprot), and HF + B groups (20.24 ± 2.36 U/mgprot) (P < .05) (Fig. 4C). The GSH levels in the HF + B group were significantly higher than in the control group (20.24 ± 2.36 vs. 13.40 ± 0.89 U/mgprot, P < .05), while no differences were found in the HF + G group (20.24 ± 2.36 vs. 16.05 ± 1.67 U/mgprot, P > .05) (Fig. 4C).
In terms of injury repair, the testicular tissue Xrcc1 gene expression in the HF group was significantly higher than that of the control (P < .01), HF + G (P < .05) and HF + B groups (P < .01) (Fig. 5A), and there were no significant differences among HF+B, HF + G and control groups (P > .05) (Fig. 5A). Compared with the control group, Ogg1 gene expression in the HF testicular tissue was significantly decreased (P < .05) (Fig. 5B). There were no differences found among the HF+B, HF + G and control groups in terms of Ogg1 gene expression (P > .05) (Fig. 5B). The expression of Apex1 gene in the testicular tissue from the HF + B (P < .01) and HF groups (P < .05) was significantly lower than the control group (Fig. 5C). The Apex1 gene expression in HF + G group was significantly higher than the HF + B group (P < .05), while no difference was observed between the control and HF groups (P > .05) (Fig. 5C).
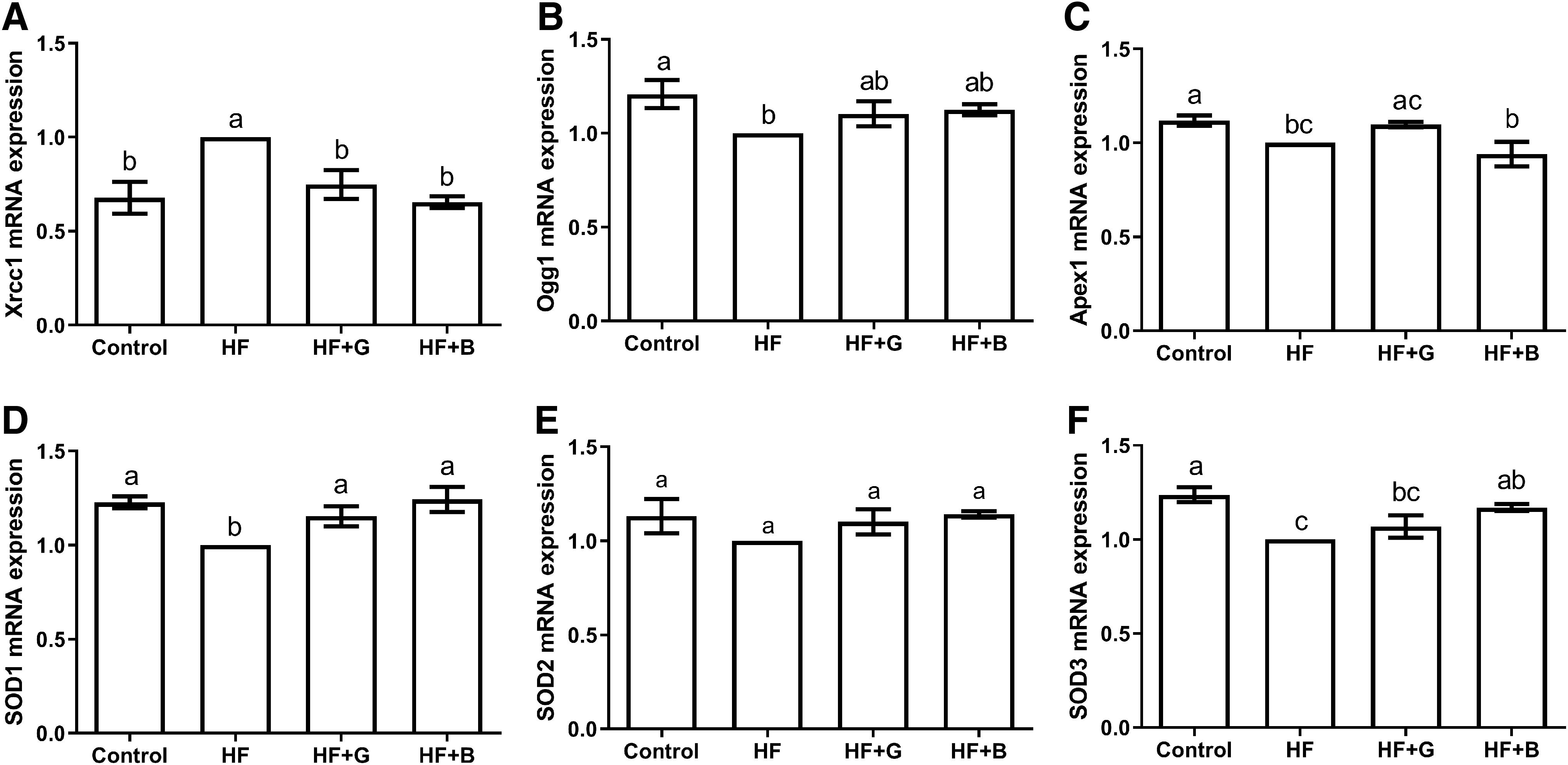
Effects of black and green tea supplements on the injury repair-related gene expression in a murine model of obesity (n = 6). The relative abundance of
In terms of oxidative stress, the expression level of SOD1 gene in the HF testicular tissue was significantly lower than that of the control, HF + B (P < .01), and HF + G groups (P < .05). We checked the raw data, recalculated and compared the expression levels of SOD1 and SOD2 genes in the testicular tissues, and found that there were no significant differences among the control, HF + G, and HF + B groups (P > .05) (Fig. 5D, E). The expression level of SOD3 gene in HF testicular tissue was significantly lower than the control group (P < .01), the HF + B group significantly higher than the HF group (P < .05), but was not different from the HF + G group (P > .05).
Discussion
For half a century, unhealthy eating habits and lifestyles have led to rising obesity rates. In men, obesity may cause a gradual decline in sperm quality and thus reduce fertility. 20,21 Tea has been listed as an important weight loss drink. 12 It can reduce the levels of cholesterol, triglyceride, and adiponectin in the serum of obese mice, thereby alleviating obesity. 22 However, it is unclear whether tea can reduce the reproductive damage caused by obesity in male animals. In this study, it was found that HF could increase the BW of male mice, as well as the rate of sperm malformation, affect the expression of insulin and leptin, and the production of androgens. The addition of tea into the diet can reduce the rate of sperm malformation, increase the expression of insulin and leptin, and partially reverse male reproductive dysfunction.
Obesity can inhibit spermatogenesis in the testes by reducing testosterone levels and increasing oxidation levels, leading to spermatogenesis disorder and increased sperm malformation rate. 23 –25 In this study, it was found that the HF-induced sperm malformation rate in male mice significantly increased, which could be ameliorated effectively by the addition of green tea and black tea. The testicular tissue from obese mice did not exhibit changes in spermatogenesis, this may be due to the fact that an 18 weeks of HF may not have been long enough to negatively influence spermatogenesis. 26
Male hormones in the testis can affect sperm production, increasing the rate of sperm malformation in human. 27,28 In this study, it was also found that obesity induced by HF also reduced the production of testosterone in male mice, and both green tea and black tea could increase the serum testosterone level to different degrees. Volle et al. also found that obesity resulted in lower sperm quality, sperm motility, and testosterone levels and higher estradiol levels, while weight loss significantly improved these parameters. 29 Cyp11a1 and Cyp17a1 are genes that express rate-limiting enzymes in the process of cholesterol synthesis into testosterone, while Cyp19a1 is a gene that expresses P450 aromatase. 7,30
In the current study, compared with the control group, we discovered that the expression levels of Cyp11a1 and Cyp17a1 genes in the high-fat group were reduced, indicating that the HF-induced obese mice decreased serum testosterone level by inhibiting the synthesis of testosterone. Muntasser et al. reported that the addition of black tea to the diet could improve the reproductive system efficiency through precluding the decreased expression of Cyp11a1 and Cyp17a1 in adult male rats exposed to the oxidative stress induced by streptozotocin. 31 Based on the gene expression profile microarray analysis, Wu et al. discovered that polyphenols and polysaccharides in black tea could regulate the expression of genes related to steroid hormone secretion, and so confirmed that black tea promoted the synthesis of testosterone and improved reproductive disorders; polyphenols and polysaccharides in black tea could also reduce BW, Lee index, visceral fat weight, and fat cell size, and improve biochemical indicators and increase fecal fatty acid content to achieve weight loss, but the specific mechanisms need to be further studied. 32
Obesity can reduce the activity of free radical scavenging enzymes in cells, produce excessive generation of ROS or imbalance of the oxidation/antioxidant system, and lead to increased cell oxidation levels. 33 –35 ROS production by sperm primarily comes from the nicotinamide adenine redox enzyme system in mitochondria. 36 High levels of ROS not only can damage the sperm cell membrane and reduce sperm motility, resulting in decreased sperm and egg binding ability, 37 but also alter sperm DNA, passing defective DNA from parents to offspring. 38
Our experimental results showed that in testicular tissue of the male obese mice induced by HF, SOD and GSH levels were lowered, the content of MDA increased, the expression of oxidative damage Xrcc1 gene upregulated, Apex1, Ogg1, SOD1, and SOD3 genes reduced, and SOD2 was unchanged, which suggested that the HF-induced obese mice caused the testicular oxidation/antioxidant system imbalance, in turn leading to testicular oxidative damage; besides that, the addition of black and green tea upregulated the gene expression of SOD1, SOD3, and Xrcc1, and improved the levels of SOD, GSH, and MDA, which indicated that black tea could remove ROS in the testis by increasing the levels of SOD and GSH, and thereby ameliorating the oxidative damage of testis.
These results are similar to those that have been well documented, 39 and the protective mechanism of tea against oxidative damage have been associated with the ability to scavenge oxygen free radicals and inhibit the lipid peroxidation of EGCG (tea polyphenols), catechins, and tea polysaccharides in tea. 40
In conclusion, our study demonstrates that tea has the potential to alleviate the adverse effects of obesity on male reproduction. Mechanism studies have shown that black tea can reduce the rate of sperm malformation, by affecting the change of androgen levels and oxidative stress. Green tea can also regulate hormone levels, but its overall effect is weaker than black tea. This study provides a research basis for the benefit of tea in male reproductive health.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the Open Fund of State Key Laboratory of Tea Plant Biology and Utilization (Grant no. SKLTOF20150203) and the National Natural Science Foundation of China (Grant no. 31972728).