
Abstract
Anacardic acid (AA), a major component of cashew nut shell liquid, has extensive bioactivities. However, little is known about its antiadipogenic properties or the mechanism that underpins them. The aim of this study was to investigate the effect of AA on 3T3-L1 preadipocyte differentiation and its mechanisms of action. AA inhibits lipid accumulation during adipogenesis in 3T3-L1 preadipocyte (IC50 = 25.45 μM). AA abrogates mRNA expressions of the genes implicated in lipogenesis and their transcription factors, especially Pparg and Cebpa. Furthermore, antibody microarray and Kyoto Encyclopedia of Genes and Genomes (KEGG) analysis results showed that the proteins implicated in the Akt signaling pathway were most likely altered by AA. Notably, upon AA treatment, heat shock protein 90 (Hsp90), a positive regulator of Akt, was decreased, resulting in Akt degradation. These findings indicate that AA, a natural product that acts as a Hsp90/Akt signaling inhibitor, may be a possible antiadipogenic agent.
INTRODUCTION
Obesity is a multifactorial, persistent relapsing condition marked by excess adiposity induced by a disparity of caloric consumption and expenditure. 1 It is mediated by a dynamic interaction of environmental causes, genetic predisposition, and human conduct. 2 Obesity's clinical burden is not limited to single organ malfunction-related morbidity and death, but rather includes dysfunctions of many tissues, implying that it is a multisystemic disorder. 3 Obese patients, in particular, are at a greater risk of diabetes, hypertension, cardiovascular disease, musculoskeletal conditions, and various cancers. 4 For this reason, obesity is closely related to an elevated risk of death, 5,6 and the current epidemic of obesity is an increasingly serious global problem.
Although lifestyle changes are strongly recommended as a first therapeutic option against obesity, such as calorie restriction and exercise intervention, pharmacotherapy, which includes orlistat, lorcaserin, and liraglutide, is also regarded as an alternative to lifestyle change in the treatment of obesity. 7 However, owing to the potentially dangerous side effects of antiobesity medications, 8 a variety of phytochemical products have been investigated to help treat obesity.
The phosphatidylinositol 3-kinase (PI3K)-Akt signaling pathway is specifically involved in the regulation of cell formation, cell cycle, proliferation, survival, and metabolism. 9 There are many polymerized targets that receive signals produced by the PI3K downstream cascade, but the serine/threonine kinase Akt is the most critical mediator in the signal transduction of the entire PI3K-Akt pathway. 10 The regulation of carcinogenesis through PI3K-Akt axis has been frequently reported, 9 but reports of the roles of PI3K-Akt in the other biological mechanisms, especially adipogenesis, are relatively limited.
Previous studies reported that the activation of the PI3K/Akt signaling mediates the adipogenic transcriptional cascade involved in adipogenesis. One of them indicated that the PI3K-AKT pathway is involved in the peroxisome proliferator-activated receptor γ (Pparγ)-CCAAT/enhancer-binding protein α (C/EBPα) implicated adipogenic conversion of mesenchymal stem cells. 11 Recently, Gun and his colleagues reported that the transcriptional activities of Pparγ and C/EBPα, which were two master transcription factors in adipogenesis, were accelerated through upregulation of the PI3K/Akt signaling pathway. 12 Based on these results, the PI3K-Akt pathway may be a good strategic target for the prevention and treatment of obesity.
Anacardic acid (AA) is a generic term for a group of closely related phenolic compounds made up primarily of salicylic acid with a 15-carbon alkyl side chain, which is the main component of natural cashew nut shell lipid obtained as a by-product of the cashew nut processing method. 12,13 Because of its multibiological functions, such as antitumor, antioxidant, gastroprotective, and antibiotic, 14 AA has gained a lot of attention as a possible lead for pharmaceutical agents. It is better known as a global inhibitor of histone acetyltransferase enzymes like p300/CBP, GCN5, and pCAF. 14 Through these mechanisms, AA mediates various cellular processes such as wound healing, osteoclastogenesis, apoptosis, angiogenesis, and autophagy. 15 –19 As above, although the evidence related to the therapeutic importance and potency of AA has been accumulating, their function in lipid metabolism and mechanisms of action are still little known.
In this study, we investigated the antilipogenic activity of AA, as well as the underlying mechanism of action, notably, regulation of heat shock protein 90 (Hsp90)/Akt signaling pathway by AA.
MATERIALS AND METHODS
Cell culture and reagents
3T3-L1 fibroblast cells purchased from the American Type Culture Collection (ATCC, Manassas, VA, USA) were seeded into a six-well plate at 2105/well and grown in Dulbecco's modified Eagle's medium (DMEM; Welgene, Daegu, Korea) supplemented with 10% (v/v) calf serum (Gibco, New York, NY, USA) and 1% (v/v) pen (Gibco) in 5% carbon dioxide (CO2) humidified atmosphere at 37°C until reaching 100% confluence. Cells were induced 2 days after confluence by incubating them in DMEM (day 0 [D0]), and differentiation to adipocytes was induced by incubating in DMEM with 10% fetal bovine serum as a supplement with 0.5 mM isobutyl methylxanthine (Sigma-Aldrich, St. Louis, Mo, USA), 10 μg/mL insulin (Sigma-Aldrich), and 0.5 μM dexamethasone (Sigma-Aldrich; isobutyl methylxanthine, dexamethasone, and insulin-supplemented (MDI) medium). After 2 days from incubation in the MDI medium (D2), the MDI medium was subsequently replaced to DMEM supplemented with 10 μg/mL insulin every 2 days until D7. The differentiation process is briefly drawn in Figure 1A. AA was purchased from Sigma-Aldrich. AA was treated as the indicated conditions in D0, replaced with media change, and harvested to the experiments.

Effect of AA on 3T3-L1 differentiation.
Lipid content measurement
After the differentiation was completed, the cells were washed in phosphate-buffered saline (PBS) and fixed for 1 h at room temperature with 4% paraformaldehyde. The fixing solution was removed, and the cells were washed thrice with PBS before being incubated for 5 min with 60% isopropanol, followed by 10 min at room temperature staining with 0.1% Oil red O (ORO) staining solution (Sigma-Aldrich). Then, excessive ORO staining reagent was washed with distilled water. The images of stained wells in six-well plate were taken using an Olympus IX51 light microscope (Olympus, PA, USA). For quantification of lipid contents, isopropanol was added to each well of six-well plate, and the optical density (OD) values (510 nm) were measured using a microplate reader (SpectraMax 190; Molecular Devices, CA, USA).
Cytotoxicity
After the differentiation was completed with AA in the 3T3-L1 cells, 10 μL of WST-1 reagent (Roche, Branchburg, NJ, USA) was added to each well of the six-well plate, and the plate was incubated in 5% CO2 humidified atmosphere at 37°C for 3 h. After that, 100 μL of the mixture was transferred to each well of 96-well plate, and the OD value at 450 nm was measured using a microplate reader (Molecular Devices).
Quantitative real-time polymerase chain reaction analysis
Cells were differentiated with or without AA at the indicated concentrations; total RNA was extracted using an RNAiso reagent (Takara, Japan). In the iCycler iQ system, quantitative real-time polymerase chain reaction (qRT-PCR) was conducted using SYBR Green reagent (Thermo Fisher Scientific, Waltham, MA, USA; Bio-Rad, Hercules, CA, USA). The primers listed in Table 1 were used to conduct PCR in triplicate. The expression levels of target mRNAs were normalized to those of loading control, glyceraldehyde 3-phosphate dehydrogenase (GAPDH) mRNA, and the ΔΔCt methodology was used to measure the relative levels of mRNA expression.
Primers for Mus musculus Used in This Study
Western blot analysis and antibodies
Following the treatment as indicated in each experiment, the 3T3-L1 cells were lysed with the radioimmunoprecipitation assay lysis buffer containing protease and phosphatase inhibitor cocktails (Roche). The lysate was centrifuged at 20,000 g for 30 min at 4°C, and the supernatant was collected. The 5 × Laemmli sample buffer was mixed with the supernatant solution and heated at 95°C for 5 min. Approximately 10 μg of the sample was loaded and separated on sodium dodecyl sulfate–polyacrylamide gel electrophoresis gel and then transferred to the nitrocellulose membrane (GE Healthcare, Madison, WI, USA). The membrane was incubated with 5% (v/v) nonfat skim milk for 30 min, followed by incubation with the indicated primary antibody overnight. The membrane was washed with Tris-buffered saline with 0.1% Tween®20 (TBST) solution (10 min, 3 times) and then incubated with the anti-rabbit or anti-mouse horseradish peroxidase (HRP)-conjugated secondary antibody (Bio-Rad). The blocked membranes were probed overnight at 4°C with the indicated antibodies, washed thrice with TBST, and sequentially incubated with the appropriate secondary anti-rabbit or anti-mouse HRP-conjugated antibody (Thermo Fisher Scientific) for 1 h. Finally, the proteins were detected with an enhanced chemiluminescence detection reagent (Thermo Fisher Scientific) using the imaging system (Vilber Lourmat, ZAC de Lamirault, France). The antibodies used in this study are as follows: Anti-HSP90 (Santa Cruz Technology, Santa Cruz, CA, USA); anti-phospho-Akt (Ser473; Cell Signaling Technology, Danvers, MA, USA); anti-Akt (Cell Signaling Technology); and anti-GAPDH (Cell Signaling Technology).
Full moon protein microarray
The signaling explorer antibody microarray was conducted by Full Moon BioSystems, Inc. (Sunnyvale, CA, USA). The whole cell lysates from 3T3-L1 cells in the absence or presence of either 20 or 40 μM of AA during the differentiation or untreated control were harvested using protein extraction buffer (Full Moon BioSystem, Inc.) and transferred on dry ice. The 1358 antibodies in the high-throughput enzyme-linked immunosorbent assay-based antibody array for quantitative protein expression profiling are designed to test for protein level changes in multiple cellular signaling pathways (
Statistical analyses
The data are provided as mean ± standard deviation (SD). GraphPad Prism was used for the statistical analyses (Version 7.04, GraphPad Software, Inc., CA, USA). One-way analysis of variance was used to assess statistical differences between mean values, followed by Bonferroni's multiple comparison test (P = .05).
RESULTS
AA blocks fat accumulation in 3T3-L1 adipocytes
Initially, to determine the antiadipogenic effect of AA on adipocyte growth, we treated the cells with 5, 10, 20, or 40 μM of AA at D0, D2, D4, and D6 with media changes during the 3T3-L1 preadipocyte differentiation (Fig. 1A), and then ORO staining was conducted. Fat accumulation in 3T3-L1 adipocytes was significantly suppressed in a dose-dependent manner (P < .05). The IC50 values for inhibitory effects of lipid accumulation by AA were 25.45 μM (Fig. 1B, C). To confirm the possibility that the cytotoxic effect of AA might decrease fat accumulation in 3T3-L1 cells, cell viability assay was conducted under the same conditions as with the ORO staining. As shown in Figure 1D, AA did not cause cytotoxicity even up to 40 μM of AA. These results demonstrated that AA suppresses lipid accumulation during 3T3-L1 differentiation without affecting cell viability.
AA suppresses mRNA expression of PPARγ, C/EBPα, and liver X receptor α transcription factors and their target genes implicated in adipogenesis in 3T3-L1 cells
Since we found that the lipid accumulation in 3T3-L1 adipocytes was significantly abrogated at 20 and 40 μM of AA compared to the control (without AA treatment) group, the concentrations were used to examine the effects of AA. Next, we performed qRT-PCR assay to evaluate whether the antiadipogenic effect of AA is implicated in suppression of lipogenesis-related gene expression. We categorized genes related to lipid metabolism into major functional variants such as transcription, fatty acid synthesis, lipid transport, and triglyceride (TG) synthesis and then measured how AA affected their mRNA expression. As shown in Figure 2A, the mRNA expression of PPAR (encoded in Pparγ), C/EBP (Cebpa), and liver X receptor (LXR; Nr1h3) was significantly reduced in a dose-dependent manner following AA therapy. The expression of fatty acid synthase (Fasn), which is involved in the mechanism of fatty acid synthesis, was almost completely obstructed by AA, but ATP-citrate synthase (Acly) which is another major gene responsible for the process was not affected (Fig. 2B). The expressions of gene related to the lipid transport process were examined, and AA suppressed mRNA expression of Cidec encoding fat-specific protein 27 (FSP27); however, mRNA expression of CD36 (encoded in Cd36) and fatty acid-binding protein 4 (Fabp4) was also not altered following AA treatment (Fig. 2C). In the process of the TG synthesis, 1-acylglycerol-3-phosphate O-acyltransferase 2 (Agpat2), monoacylglycerol acyltransferase (Mogat1), and diacylglycerol acyltransferase 2 (Dgat2) mRNA expressions were effectively suppressed by AA; however, it was shown that AA did not affect the mRNA expression of other Agpat family (Agpat1, 3, and 4) and Dgat1 (Fig. 2D). Collectively, it was suggested that AA inhibits mRNA expression of lipogenesis-related genes and their transcription factors, PPARγ, C/EBPα, and LXRα.
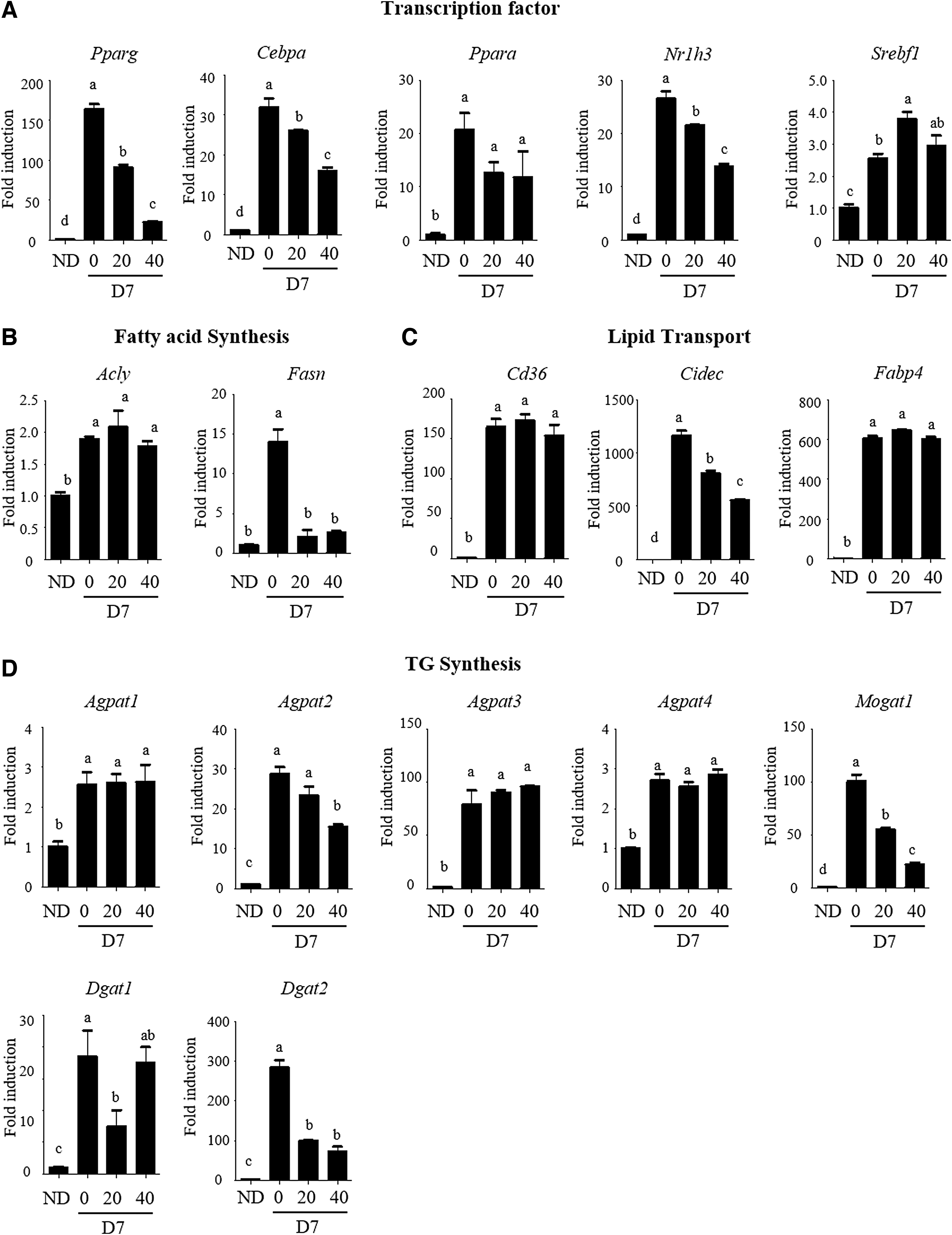
Effect of AA on mRNA expression of the genes involved in adipogenesis. The mRNA levels of the genes regulating gene transcription
AA affects the PI3K-Akt signaling pathway
To better delineate the signaling consequences of AA exposure in the observed antilipogenic effects, we utilized a large-scale proteomics analysis using an antibody array (Full Moon Microarray) to identify proteins differentially controlled in the AA treated-3T3-L1 adipocytes. From the result of Full Moon Microarray which consists of duplicated 4 blocks containing antibodies against total and phosphor proteins, including 1358 proteins in more than 30 different cell signaling pathways, we selected the proteins which showed 1.25 times increase in induction compared to control (IND/NONE >1.25) and 0.8 times decrease in 40 μM of AA treatment compared to induction (AA40/IND <0.8), or IND/NONE <0.8 and AA40/IND >1.25, and then a total of 32 proteins was identified (Fig. 3A). Using gene ontology enrichment analysis in terms of KEGG pathway, 20 we analyzed 19 commonly upregulated- or 13 downregulated proteins following AA treatment during 3T3-L1 adipocyte differentiation, and then as shown in Figure 3B, the findings revealed that the proteins were highly enriched in the modulation of the PI3K-Akt signaling pathway. Following AA treatment, many proteins that modulate or are modulated by the PI3K-Akt signaling axis, such as Hsp90 or nitric oxide synthase 3, were affected. Overall, AA regulated 3T3-L1 adipocyte differentiation by modulating the pathway.

AA modulates the proteins involved in PI3K-Akt signaling pathway.
AA inhibits Hsp90/Akt signaling axis
Hsp90 is a molecular chaperone and positive regulator of Akt that directly binds to Akt and maintains its activity. 21 Consistent with the antibody array result (Fig. 3A), Hsp90 was substantially upregulated in differentiated 3T3-L1 cells and decreased in a dose-dependent manner after AA treatment (Fig. 4A). Hsp90 inhibition results in Akt dephosphorylation and destabilization, followed by degradation. 22 As shown in Figure 4A, Akt and its phosphorylation level were also decreased by AA. To evaluate the possibility that AA can decrease Akt protein at the transcriptional level, cycloheximide (CHX), a protein synthesis inhibitor, was used in the presence or absence of AA in the differentiated 3T3-L1 cells. Blockage of protein synthesis showed that AA did not transcriptionally affect Akt level in 3T3-L1 cells (Fig. 4B). Consistent with the result, AA treatment did not affect Akt mRNA expression in 3T3-L1 cells (Fig. 4C). Thus, we evaluated whether AA decreases the level of Hsp90, an upstream positive regulator of Akt at the transcriptional or protein level. AA did not decrease Hsp90 protein levels in the presence of CHX (Fig. 4B), and AA specifically inhibited Hsp90b1 expression in the HSP90 gene family, Hsp90aa1, Hsp90ab1, and Hsp90b1, in 3T3-L1 cells (Fig. 4D). These results demonstrated that AA suppresses Hsp90-Akt signaling axis through inhibition of Hsp90 mRNA expression in 3T3-L1 adipocytes.
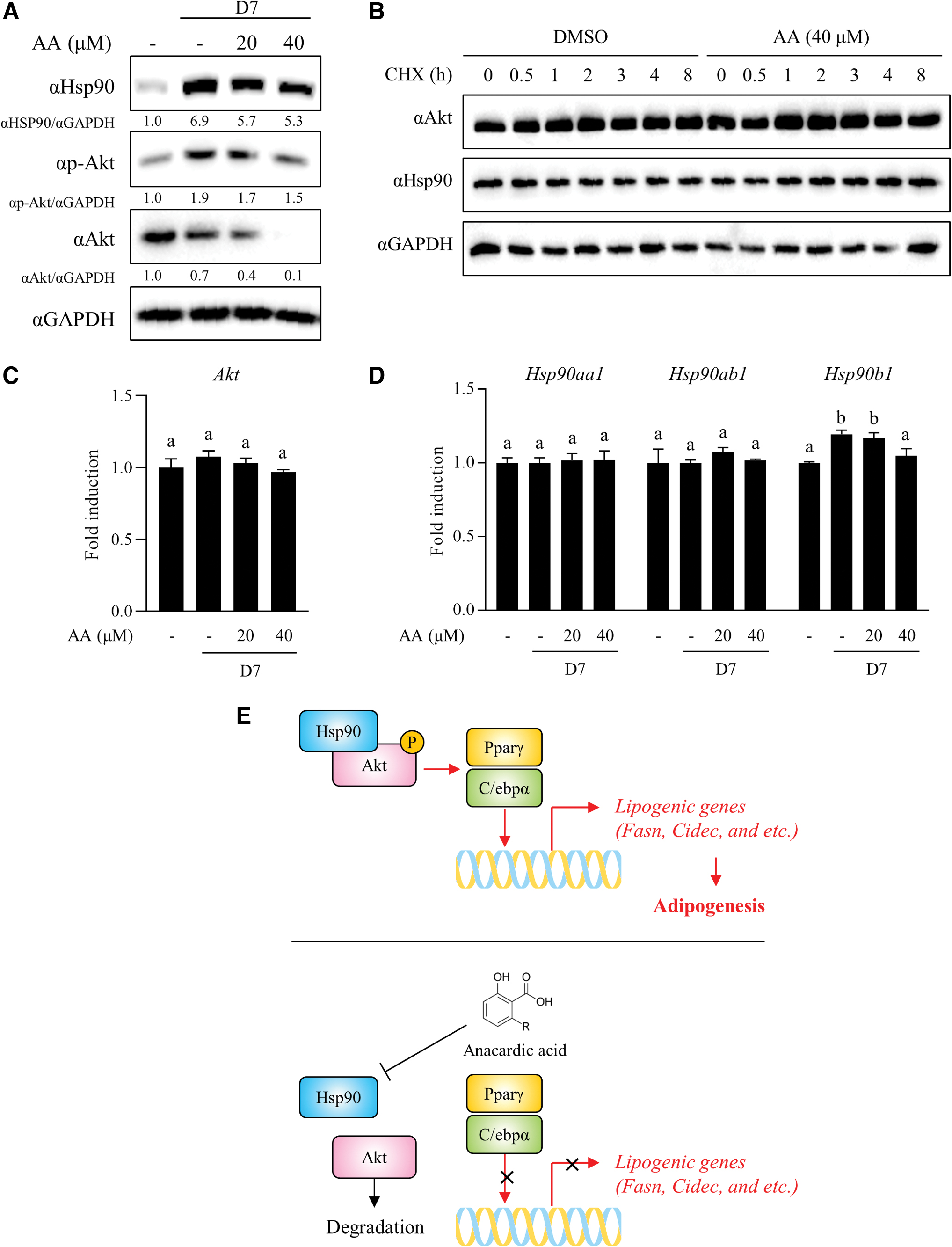
AA inhibited Hsp90/Akt signaling axis.
DISCUSSION
In addition to lifestyle changes, pharmacological approaches are also recommended to support the treatment of obesity. Antiobesity medications such as Contrave, orlistat, and Qsymia have been developed and approved by the FDA; however, potential side effects can occur during long-term use. Regulation of obesity by natural product-derived chemicals has been expected to have advantageous effects in terms of their safety. 23 In the present study, we find that AA, a key component of cashew nut shell liquid, exerts an antiadipogenic effect by inhibiting the HSP90/Akt signaling pathway.
PPARγ and C/EBPα are a part of delicate network of transcription factors, which regulate the adipocyte differentiation. 24 Especially, PPARγ is almost exclusively predominant in adipocytes 25 and triggers and facilitates the processes with C/EBPα. 26,27 Our results demonstrated that AA suppressed Pparγ and Cebpa mRNA expression in differentiated 3T3-L1 cells. Several studies have shown that PPARγ positively regulates Mogat1, 28 Fasn, 29 and Fsp27 30 expressions, and combined knockdown of Pparγ and Cebpa substantially reduced Cidec expression. 27 Consistent with the previous studies, AA inhibited Mogat1, Fasn, and Cidec mRNA expression, demonstrating that AA inhibits the expression of adipogenesis-related genes and their transcription factors at the mRNA level.
Two studies have reported that blockage of Hsp90 inhibits 3T3-L1 differentiation and fat accumulation. 31,32 Antibody array and subsequent pathway analysis results revealed that AA suppressed the protein expression involved in the PI3K-Akt signaling pathway; notably, Hsp90, a positive regulator of Akt activity, 21,23 was inhibited by AA treatment during 3T3-L1 differentiation. In addition, downregulation of Hsp90, Akt, and its phosphorylation were validated by western blotting. The PI3K-Akt pathway is an upstream pathway that controls adipogenic transcription factors Ppar and C/EBP and accelerates adipogenesis when activated. 11,12 Interestingly, AA did not directly affect the mRNA level and protein stability of Akt, and the expression of Hsp90b1 gene encoding HSP90β isoform was specifically inhibited by AA without modulating HSP90 protein stability in 3T3-L1 cells. HSP90β but not HSP90α directly binds to the amino acid residues 229–309 of Akt (pivotal residues for Akt kinase activity) and inhibits Akt activity in vivo. 21 In addition, HSP90β isoform suppresses intestinal epithelial cell apoptosis through stabilizing Akt. 33 These studies supported our suggestion that AA suppresses adipogenesis through Hsp90β/Akt/Pparγ and C/EBPα signaling axis. However, it is needed to further confirm the mechanisms of action involved in the specific inhibitory effect of HSP90β at the transcriptional level of transcription factors and epigenetic machinery.
In conclusion, we discovered that AA has an antilipogenic effect and revealed the underlying mechanism in 3T3-L1 adipocytes. AA inhibited the mRNA expression of lipogenic genes and transcription factors, most specifically Ppar and C/ebp. AA also inhibited the PI3K-Akt signaling pathway, which is an upstream pathway of Ppar and C/ebp, by reducing Hsp90, followed by Akt inactivation and degradation (Fig. 4E). Therefore, AA could be a novel Hsp90 modulator as a potential antiobesity agent derived from natural products.
Footnotes
AUTHOR DISCLOSURE STATEMENT
No competing financial interests exist.
FUNDING INFORMATION
This work was supported by the Main Research Program (Grant No. E-0210601) of the Korea Food Research Institute (KFRI), funded by the Ministry of Science, ICT & Future Planning.