
Abstract
In this study, we describe the effects of Lactobacillus paracasei HY7015 (HY7015) on promoting mouse hair growth. Since our purpose was to increase hair growth through oral administration, medicinal yeast, at a suitable concentration for application in mice, was used as a positive control. First, experiments were conducted to determine the effect of HY7015 on proliferation of hair follicle dermal papilla cells (HFDPC), which are important contributors to hair growth. HY7015 stimulated HFDPC proliferation in vitro and increased their secretion of vascular endothelial growth factor and insulin-like growth factor-1. In mouse experiments, oral administration of HY7015 promoted hair growth and hair follicle maturation in the dorsal skin, as well as increasing growth factor levels in mouse serum. In summary, we demonstrate that L. paracasei HY7015 consumption can promote hair growth by stimulating HFDPC proliferation and growth factor secretion. Follow-up studies are warranted to determine the underlying mechanism, using various approaches, including investigation of changes in intestinal microbiota and alteration of gene and protein expression.
Introduction
Hair loss results from skin disorders and has significant impacts on the mental health of the patients affected. 1 Although there are no established effective treatments for hair loss, some therapies focused on prevention and mitigation are available. 2 Hair loss primarily occurs in middle-aged men; however, in recent years, it has also increased among women and children. 3,4 Various factors can contribute to hair loss, including mental stress, changes in hormone (particularly testosterone) levels, and environmental influences. 5 Hair loss can be classified as alopecia areata, anagen effluvium, androgenic alopecia, telogen effluvium, tinea capitis, trichorrhexis nodosa, and trichotillomania. Alopecia areata, also known as spot boldness, which manifests as partial or complete hair loss, can be caused by inflammation and mental health problems, and is best treated using anti-inflammatory drugs and rest. 6 Anagen is the hair growth phase, and lasts 2–6 years in humans; catagen is the transitional phase between anagen and telogen; and telogen, which lasts for ∼3–5 months, is a resting phase before hair falls out. Around 90% of total hair is anagen, 10% in telogen, and 1% in catagen, and disruption to this cycle can cause hair loss. Anagen effluvium occurs when anagen is abnormally shortened, resulting in an overall decrease in hair growth, whereas telogen effluvium is caused by abnormally rapid telogen conversion, resulting in increased hair loss. 7
Only two drugs for hair loss prevention are currently approved by the U.S. Food and Drug Administration: minoxidil and finasteride. Minoxidil prevents hair loss and promotes hair growth by influencing blood circulation, cell proliferation, and growth hormone stimulation in the hair follicle through KATP channel regulation 8 ; however, it has side effects, including hair loss and hypotension. 9 Finasteride is an inhibitor of the hair loss inducer, dihydrotestosterone (DHT). Physiologically, DHT is produced by conversion from testosterone by type II 5α-reductase, an enzyme inhibited by finasteride, which is an effective treatment for androgenic alopecia; however, it has side effects, including infertility and depression, and is not recommended for use in women or children. 10 Overall, both drugs are effective anti-hair loss agents, but there are major concerns about their side effects and they are only preventive agents. Therefore, further research to identify safe substances that can ameliorate hair loss without serious side effects is required and efforts are ongoing. For example, RE-ORGA, a herbal product, has been reported to prevent hair loss by directly inhibiting 5α-reductase activity. 11 In addition, a clinical research article reported the effects of probiotics from kimchi and cheonggukjang on androgenic alopecia. 12
Probiotics are defined by the World Health Organization (WHO) as “Live microorganisms which, when administered in adequate amounts, confer a health benefit on the host”. 13 Most probiotics are lactic acid bacteria of the genera Lactobacillus, Bifidobacterium, Lactococcus, and Streptococcus. Lactobacillus is major probiotic genus found in traditional fermented foods, such as fermented dairy products. This genus inhabits the gastrointestinal tracts of humans and animals, providing health benefits, 14 including important roles in improving host intestinal homeostasis. 15,16 Therefore, the anti-inflammatory effects of Lactobacillus species have been studied in the context of inflammatory bowel diseases, including ulcerative colitis, Crohn's disease, celiac disease, and microscopic colitis. 17,18 Furthermore, various potential functions of Lactobacillus spp. have been investigated, including studies of their effects on hair growth. 19,20 In a previous study, hydrolysates of Lactobacillus plantarum improved hair growth-related vascular regeneration by stimulating vascular endothelial growth factor (VEGF) production in C57BL/6 mice. 19 The effects of plant extracts fermented by L. plantarum or Lactobacillus rhamnosus on hair growth have also been evaluated by histological analysis and investigation of the expression of hair growth-related genes, including VEGF and keratinocyte growth factor. 20,21
In this study, we explored whether Lactobacillus paracasei HY7015 (HY7015), a probiotic bacterial strain, can prevent hair loss, by conducting hair growth experiments in mice. HY7015 was isolated from kimchi fermented with lactic acid bacteria. Lactic acid bacteria influence intestinal peristalsis and immune regulation, and have antioxidant activity 17,22,23 and oxidative stress and scalp inflammation cause hair loss 24,25 ; hence, supplementation with lactic acid bacteria was predicted to promote hair growth through antioxidant, anti-inflammatory, and VEGF secretion-inducing effects. 19 In addition, Lactobacillus bacteria have “Generally Recognized as Safe” (GRAS) status, and they have been used in food products for many years, and with consumption over long periods of time. 26 Based on its effects on dermal papilla cell proliferation, HY7015 was selected for further experiments in dermal papilla cells and animal models.
Materials and Methods
Reagents and equipment
Reagents and equipment used in this study included L, paracasei HY7015 (HY7015; Korea Yakult Co., Ltd.; Seoul, Korea), Mouse VEGF (MMV00), Mouse/Rat IGF-I/IGF-1 (MG100), and Human VEGF (DVE00) Quantikine ELISA kits (R&D Systems; Minneapolis, MN, USA).
Cell culture and proliferation assay
Human hair follicle dermal papilla cells (HFDPC) were purchased from Cell Applications (San Diego, USA). Cryopreserved cells were cultivated using a HFDPC medium and supplement kit (Cell Applications) in T-75 flasks at 37°C in a humidified 5% CO2 atmosphere. When cells reached 80% confluence, they were detached using trypsin and subcultured. To assess cell proliferation, 5.0 × 103 cells were seeded in each well of 24-well plates. Attached cells were washed with phosphate-buffered saline, treated by adding medium containing 2% fetal bovine serum (FBS) alone or supplemented with 50 μg/mL medicinal yeast (MY; positive control) or HY7015 (1 × 106 CFU/mL), and were incubated for 24 or 48 h before cell proliferation measured using the 3-[4,5-dimethylthiazol-2-yl]-2,5 diphenyl tetrazolium bromide (MTT) method. 27 In brief, culture medium was replaced with medium containing 500 μg/mL MTT solution and incubated for 4 h. Resulting MTT crystals were dissolved in dimethyl sulfoxide (DMSO) and absorbance measured at 570 nm.
Measurement of HFDPC growth factor secretion
HFDPC were cultured in 24-well plates (5.0 × 103 cells/well) for 24 h. After cells attached, they were treated with medium containing 2% FBS alone or supplemented with 50 μg/mL MY or HY7015 (1 × 106 CFU/mL). Medium containing 10% FBS was used as a positive control. After culturing for 48 h, growth factor secretion assays of culture supernatants were conducted using Human VEGF and Human IGF-1 Quantikine ELISA kits.
In vivo assays
Experimental procedures were approved by the Ethics Review Committee of the Korea Yakult Company Limited R&D center, Korea (AEC-2020-00001-Y). Female C57BL/6 mice (6 weeks old) were purchased from Dooyeol Biotech (Seoul, Korea). Mice were divided into the following groups (n = 8 each): normal controls (Con), mice fed 200 mg/kg MY (MY), and mice fed HY7015 (1 × 108 CFU/day mouse) (HY7015). At 7 weeks old, mice were anesthetized and their dorsal hair removed using animal clippers. After depilation, mice in the HY7015 and MY groups were fed with probiotics or medicinal yeast, respectively, mixed with normal feed for 7 weeks. After treatment was completed, mice were sacrificed and dorsal skin and blood samples collected. Blood was allowed to stand at room temperature for 30 min and then centrifuged at 3000 g to separate serum. Collected serum and tissues were stored at −80°C until use.
Growth factor measurement in mouse serum
Frozen serum samples were thawed for growth factor measurement using mouse VEGF and IGF-1 Quantikine ELISA kits.
Preparation and analysis of dorsal skin samples
Dorsal skin samples were fixed in 10% formalin solution and used to prepare hematoxylin and eosin (H&E)-stained slides by KPC (Gwangju-si, Korea). Dorsal skin thickness was measured under an Olympus CK2 microscope (Tokyo, Japan) at 100 × magnification and images analyzed using Image J software (National Institutes of Health, Bethesda, MD, USA) to determine the number of hair follicles per unit area ( × 104/μm2)
Statistical analysis
Data are shown as the mean ± standard deviation (SD) of independent experiments. Statistical comparisons between groups were performed by analysis of variance (ANOVA), followed by Tukey's test, using SPSS version 12.0 (SPSS, Inc., Chicago, USA); P < .05 was regarded as significant.
Results
Effect of HY7015 on HFDPC proliferation
To suppress HFDPC proliferation, samples were cultured in low-serum (2% FBS) medium. After 24 h, proliferation of positive control cells treated with MY was significantly increased (118.19% ± 6.04%; P < .05) relative to untreated controls. Treatment with HY7015 also increased proliferation (111.12% ± 0.64%); however, the difference was not significant (P > .05). After 48 h, cell proliferation was significantly increased in both groups; MY (159.24% ± 9.27%; P < .01) and HY (123.18% ± 0.79%; P < .05) (Fig. 1).
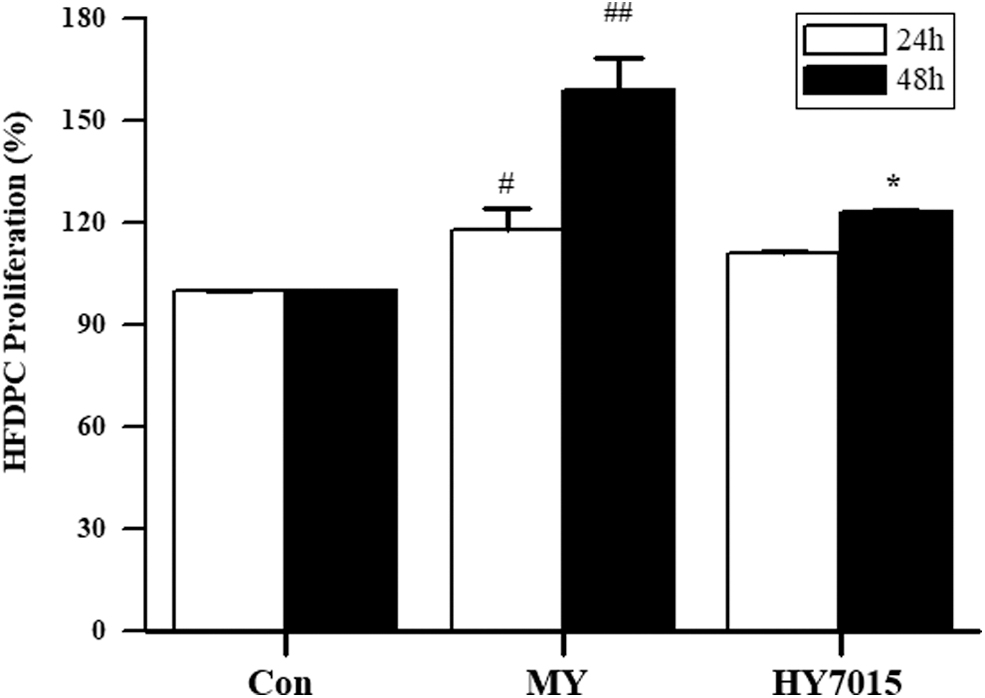
HY7015 promotes HFDPC proliferation. HFDPC were untreated (Con), or treated with L. paracasei HY7015 (1 × 106 CFU/mL) (HY7015) or medicinal yeast (50 μg/mL) (MY) for 24 or 48 h. Data are presented as mean ± SD. Significant differences are indicated by # P < .05, ## P < .01 (control group vs. MY group) and *P < .05 (control group vs. HY7015 group). HFDPC, hair follicle dermal papilla cells; SD, standard deviation.
HY7015 stimulates increased VEGF and IGF-1 secretion by HFDPC
Next, we evaluated whether treatment with HY7015 influenced VEGF and insulin-like growth factor-1 (IGF-1) secretion by HFDPC by measuring VEGF and IGF-1 levels in culture supernatants after treatment for 48 h. VEGF levels were 64.16 ± 1.02, 156.20 ± 21.32, and 161.88 ± 5.24 pg/mL in the Con, MY, and HY7015 groups, respectively (Fig. 2A), representing significant increases of 2.43-fold (P < .05) and 2.52-fold (P < .01) in the MY and HY7015 groups, respectively, relative to the Con group. There was no significant difference between the HY7015 and MY groups (Fig. 2A).
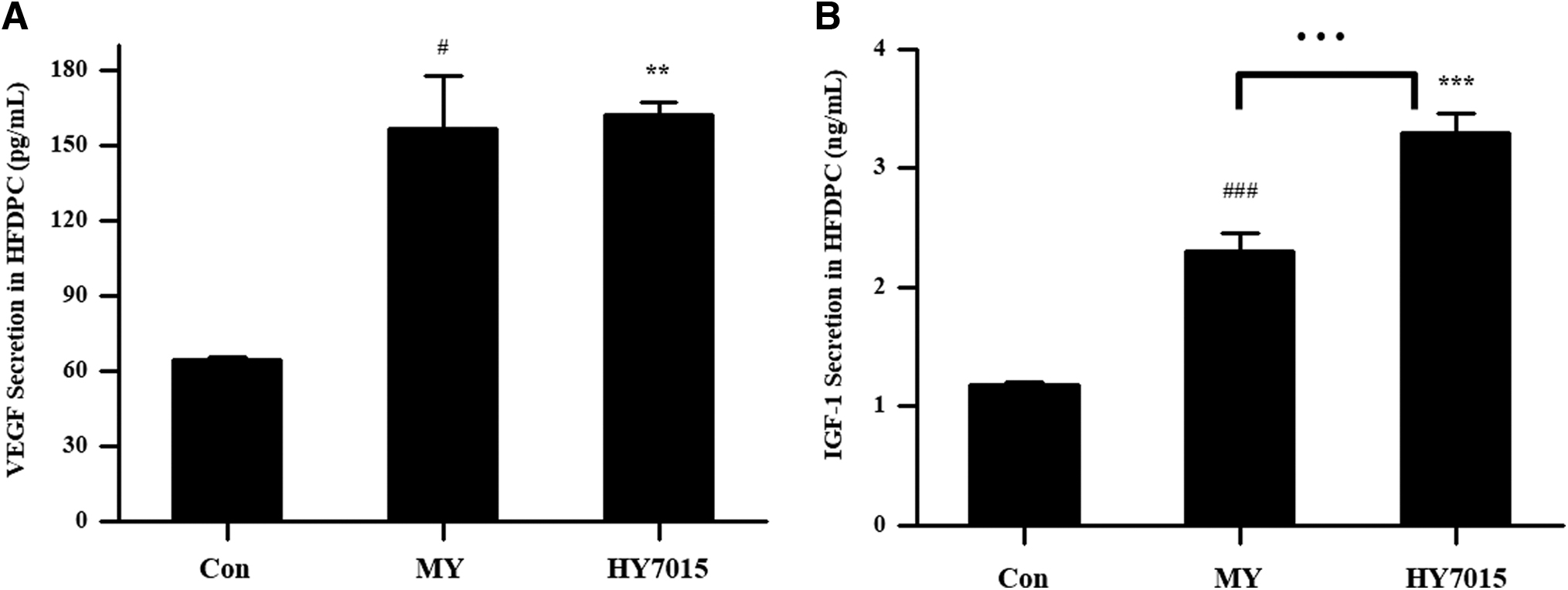
HY7015 increases HFDPC growth factor secretion. HFDPC were untreated (Con), or treated with L. paracasei HY7015 (1 × 106 CFU/mL) (HY7015) or medicinal yeast (50 μg/mL) (MY) for 48 h.
Similarly, secretion of IGF-1 was 2.30 ± 0.15 ng/mL in the MY group and 3.30 ± 0.16 ng/mL in the HY7015 group, which were significant increases relative to the Con group (1.18 ± 0.02 ng/mL; both P < .001) (Fig. 2B). Furthermore, HY7015-treated cells secreted significantly more IGF-1 than those in the MY group (P < .001) (Fig. 2B).
Promotion of mouse hair growth by oral HY7015 intake
The effects of HY7015 on mouse hair growth were assessed using a telogenic mouse model. C57BL/6 mice enter telogen at 7 weeks old, after cessation of hair growth 28 and entry into catagen, and hair growth stops for ∼3 weeks. During this period, we assessed whether ingestion of HY7015 promoted hair growth. Lyophilized live probiotics were mixed with mouse feed and administered to the mice and observation of mouse hair demonstrated promotion of hair growth 7 weeks after treatment (Fig. 3A).

Oral administration of HY7015 promotes hair growth in a telogenic mouse model. Shaved 7-week-old mice consumed nonsupplemented feed (Con), or feed supplemented with L. paracasei HY7015 (108 CFU/day) (HY7015) or medicinal yeast (200 mg/kg) (MY) daily for 7 weeks.
Comparisons of hair growth rates in hair removal areas showed 11.31% ± 11.38% growth in the control group, compared with 79.36% ± 15.79% and 72.89% ± 23.21% in the MY and HY7015 groups, respectively, which were both significantly higher than controls (both P < .001) (Fig. 3B). Hence, HY7015 can promote hair growth in a telogenic mouse model.
Weekly assessment of hair growth in each group revealed rates of 6.22% ± 6.59% and 11.31% ± 11.38% in the control group in the 6th and 7th weeks, respectively (Fig. 3C). By contrast, in the MY group, hair growth was observed from the 5th week of (19.33% ± 17.10%), with growth of 46.19% ± 19.92% (P < .05) and 79.36% ± 15.79% (P < .001) in the 6th and 7th weeks, respectively (Fig. 3C). Ingestion of HY7015 led to similar hair growth to that observed in the MY group, which was observed from the 4th week. Significantly increased hair growth was observed in the HY7015 group relative to controls, with rates of 1.78% ± 5.04% (P > .05), 27.78% ± 31.15% (P < .05), 42.22% ± 39.63% (P < .05), and 72.89% ± 23.21% (P < .001) at 4, 5, 6, and 7 weeks, respectively (Fig. 3C). These data confirm that ingestion of HY7015 and MY significantly promoted hair growth in telogenic mice.
Effect of HY7015 treatment on hair follicles and dorsal skin thickness
Next, we stained cross-sectional and longitudinal sections of mouse skin samples with H&E and evaluated the numbers and shapes of hair follicles (Fig. 4A). Compared with controls (3.51 ± 7.95 × 104 hair follicles/μm2), significantly more hair follicles were detected in the positive control MY (24.11 ± 17.24 × 104 hair follicles/μm2; P < .01) and HY7015 (25.28 ± 16.87 × 104 hair follicles/μm2; P < .01) groups (Fig. 4B). Hence, the effects of HY7015 treatment on mouse hair growth were consistent with those on hair follicle numbers.
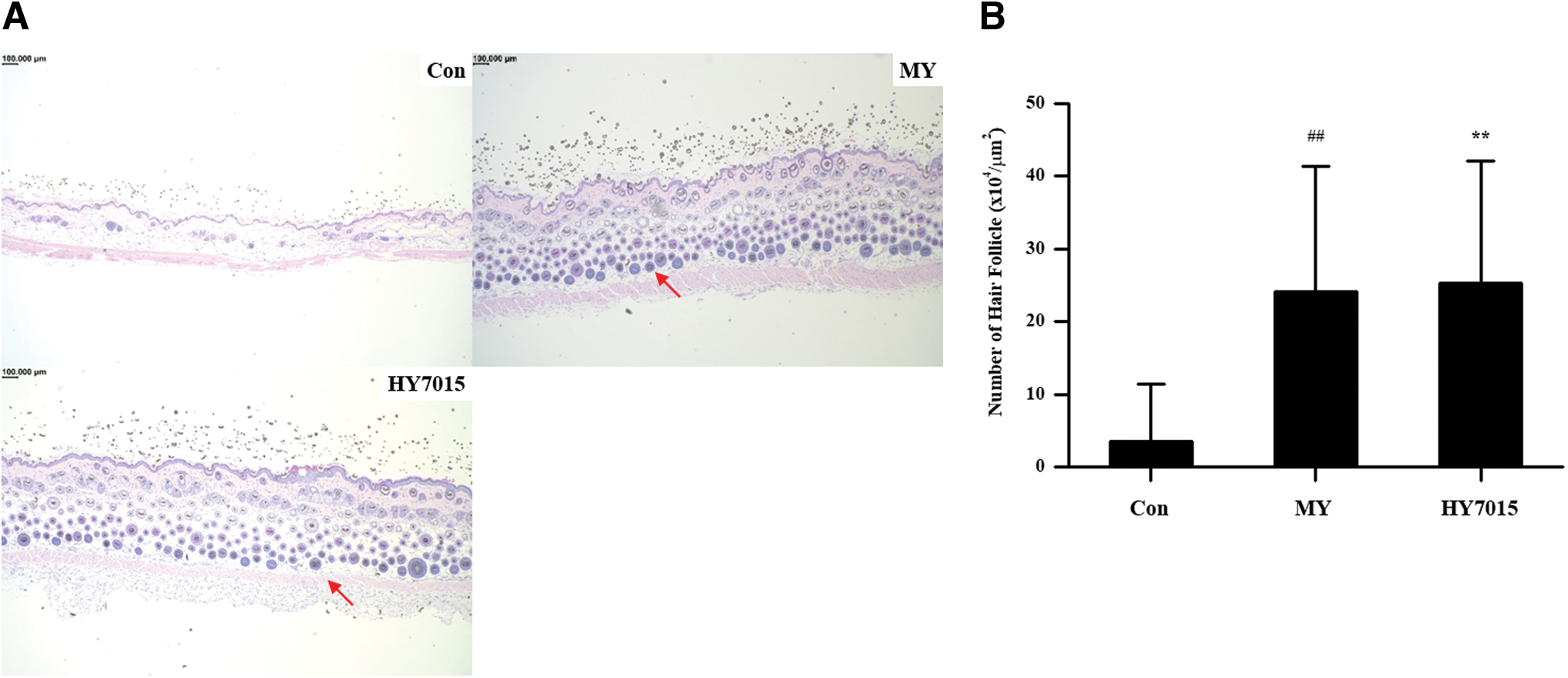
Oral administration of HY7015 stimulates hair follicles in telogenic mice.
Hair growth is accompanied by an increase in epidermal thickness and numbers of hair follicles. 29 Therefore, effects on hair growth can be predicted by measuring dermal layer thickness. Mouse epidermal skin thickness was measured using H&E stained samples. No growing hair follicles (in anagen) were detected in the control group by microscopic observation; however, such follicles were present in the MY and HY7015 groups (Fig. 5A), indicating hair growth promotion in treated mice. Measurement of epidermal thickness using Image J software demonstrated mean ± SD values of 28.20 ± 9.45, 55.00 ± 23.59, and 68.74 ± 28.14 μm in the control, MY, and HY7015 groups, respectively (Fig. 5B), representing significant increases in both the MY (P < .05) and HY7015 (P < .001) groups; there was no significance between the MY and HY7015 groups (P > .05; Fig. 5B). Taken together, these data indicate that HY7015 ingestion contributes to hair growth by increasing hair follicle numbers and dermal layer thickness.
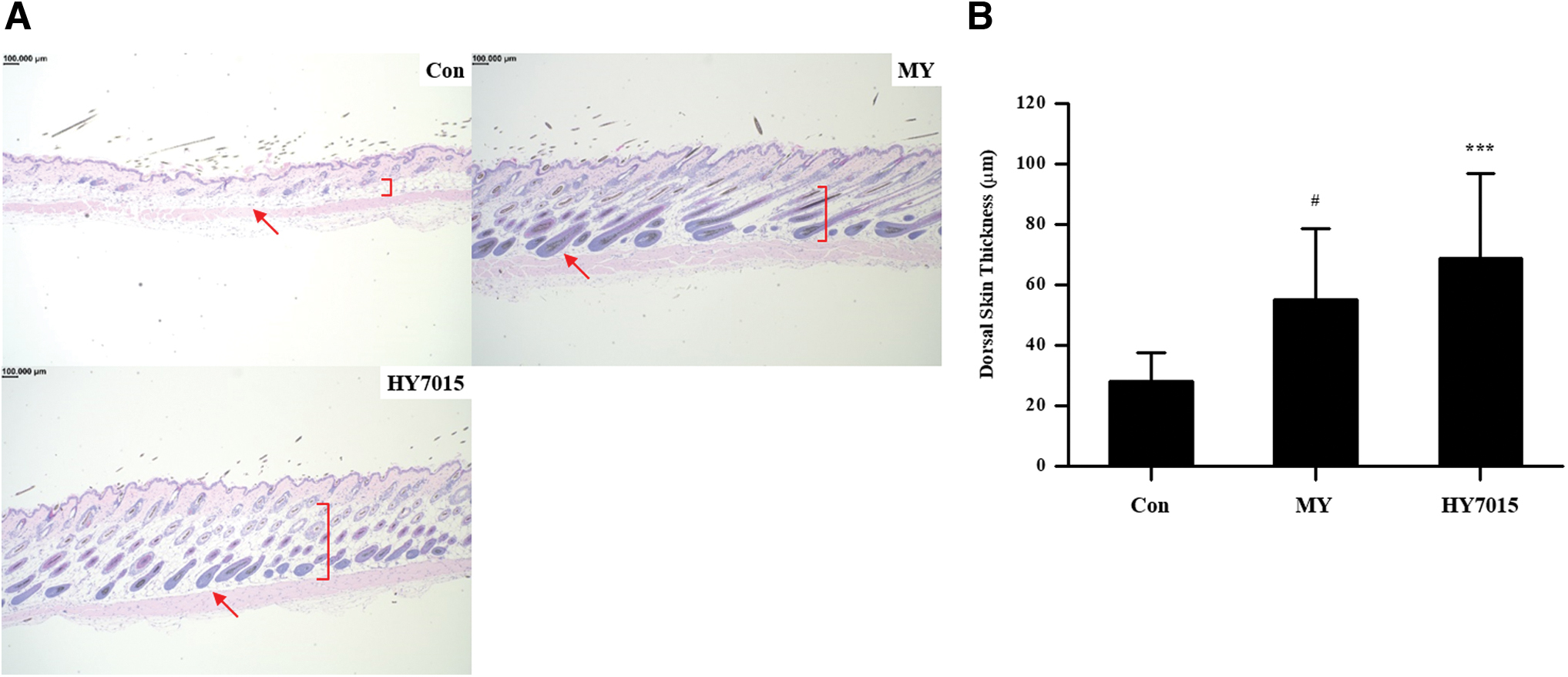
HY7015 treatment increases mouse dorsal skin thickness.
HY7015 treatment stimulates increased growth factor secretion in mouse serum
Finally, levels of growth factors that contribute to hair growth were measured in mouse serum, to assess the effects of HY7015 treatment in vivo. VEGF, a proangiogenesis factor, showed results consistent with those observed in HFDPC; measurements were 31.98 ± 11.39, 60.96 ± 5.11, and 58.02 ± 11.76 pg/mL in the control, MY, and HY7015 groups, respectively (Fig. 6A). Compared with the control group, levels were significantly increased in both the MY and HY7015 groups (both P < .001) (Fig. 6A).
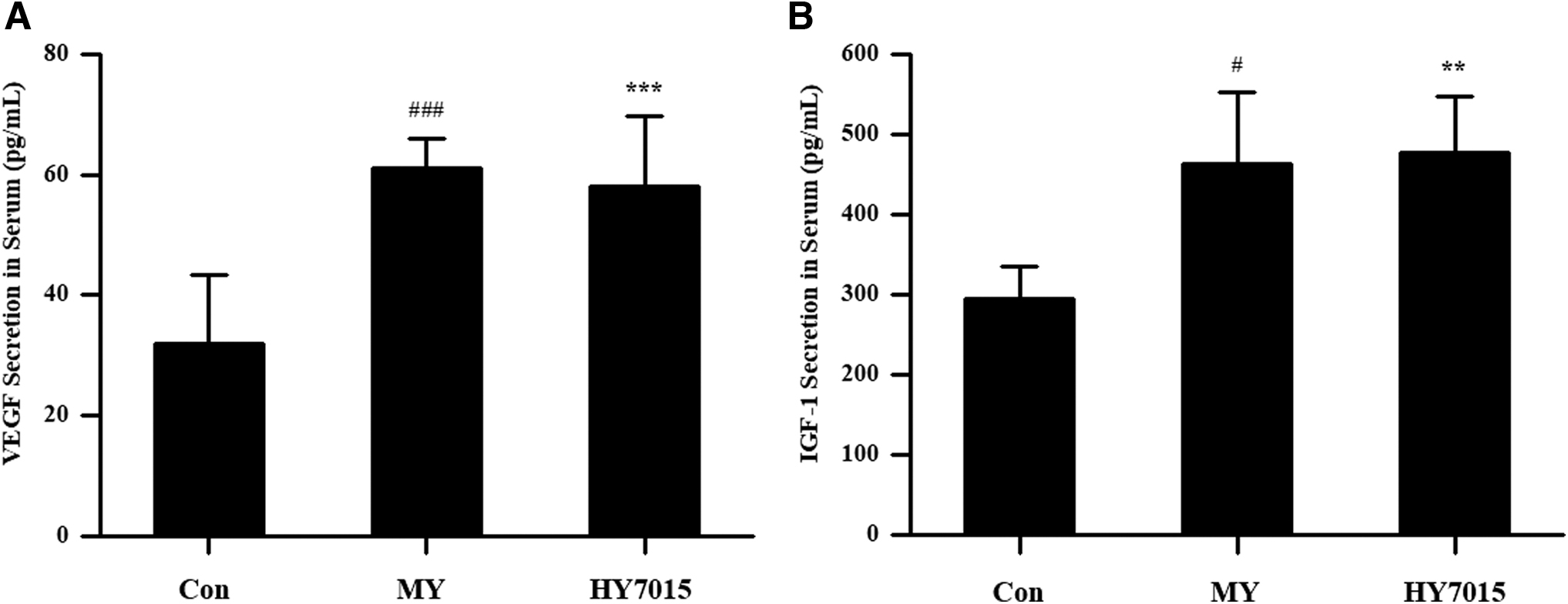
Oral administration of HY7015 stimulates growth factor secretion in mouse serum.
Measurement of IGF-1 showed similar results to that of VEGF, with significantly increased levels in both the MY (462.82 ± 90.01 pg/mL; P < .05) and HY7015 (477.10 ± 70.25 pg/mL; P < .01) groups compared with controls (294.95 ± 39.15 pg/mL) (Fig. 6B).
Discussion
Hair follicles and hair are important organs in mammals that function to maintain body temperature and as a physical barrier to the external environment. 30 Hair loss is recognized as a problem and societal focus has changed from existing to living well, including increased interest in the cosmetic aspects of hair and facial features. Furthermore, hair loss can cause mental health problems.
In this study, we found that HY7015 can promote HFDPC proliferation (Fig. 1), particularly after 48 h of treatment. As dermal papilla cells are key to hair growth, substances inducing their proliferation can be predicted to promote hair growth. 31 Furthermore, HY7015 treatment for 48 h strongly promoted VEGF secretion (Fig. 2A). VEGF is a powerful regulator of physiological and pathological angiogenesis, as well as contributing to follicle formation during hair growth; hence VEGF induction by HY7015 is predicted to stimulate hair growth. 32,33 The IGF-1 growth factor can also control oxidative stress caused by hydrogen peroxide in keratinocytes. 34 Since increased IGF-1 contributes to hair growth, 35 promotion of its secretion by dermal papilla cells is highly likely to stimulate hair growth. Furthermore, treatment of HFDPC with HY7015 promoted IGF-1 secretion even more strongly than the positive control, MY (Fig. 2B). Together, the results of our cell culture experiments predict that HY7015 can stimulate hair growth by promoting dermal papilla cell proliferation and growth factor secretion.
The effects of HY7015 ingestion in promoting hair growth in vivo were evaluated using a telogenic mouse model. In C57BL/6 mice, hair begins to grow after birth and hair maturation is completed at 7 weeks of age; therefore, these animals can serve as a mouse model for assessment of hair growth effects in telogen. 32 In previous studies, substances used to promote hair growth have generally been applied to dorsal skin to facilitate drug delivery; however, we administered HY7015 by mixing small portions of lyophilized live bacteria with feed for 7 weeks. MY, a substance that can prevent hair loss, was also supplied at 200 mg/kg mixed in feed as a positive control. Hair growth was observed each week, and hair growth rates calculated relative to the total epilation area. Growth was significantly higher in the MY group than in controls (P < .001, Fig. 3B) and HY7015 had a similar effect on hair growth as did MY. Furthermore, hair growth was rapidly induced in the MY and HY7015 groups, with significant growth promotion relative to controls from 5 weeks after ingestion (Fig. 3C).
H&E staining was performed to determine whether HY7015 affected hair follicles directly. Since hair growth occurs in the hair follicle, when hair growth increases, the size or number of hair follicles also rises. 8 Numbers of hair follicles were significantly higher in the HY7015 group relative to controls and were similar to those in the positive control group (MY; Fig. 4B). In addition, observation of the proportions of hair follicles in anagen phase revealed a low proportion in the control group that increased in mice fed HY7015 (Fig. 5A). When hair follicles are in anagen, the epidermal layer thickens 36 ; therefore, we measured epidermal layer thickness and found that it was significantly increased after HY7015 intake (Fig. 5B). Overall, H&E staining of dorsal skin showed that HY7015 intake increased the number of hair follicles and their degree of maturation, alongside hair growth.
The effects of HY7015 on growth factors in vivo were assessed by analysis of mouse serum. VEGF levels were significantly increased in the HY7015 group relative to the control group, with similar results for IGF-1 (Fig. 6), confirming that HY7015 can stimulate growth factor secretion in vivo, as well as in cultured dermal papilla cells.
Overall, our data show that ingestion of HY7015 can stimulate hair growth by promoting growth factor secretion and dermal papilla cell proliferation, both in vitro and in mouse experiments. This study highlights the potential for further expansion of lactic acid bacteria functional applications and could stimulate further research into foods that have no side effects and can safely stimulate hair growth; however, since this study focused on promotion of hair growth, rather than the processes involved, further research on the underlying mechanisms is warranted. Probiotics can regulate biomolecules, such as those involved in antioxidant and anti-inflammatory pathways, as well as acting as regulators of the intestinal flora. Therefore, research into relationships between the intestinal flora and hair growth will be key to understanding probiotic hair growth agents.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this study.