
Abstract
Previously, we reported that the administration of a mixture of Humulus japonicus (MH) increased the longitudinal bone growth rate in Sprague Dawley rats. In this study, we investigated the effects of the dietary administration of MH on longitudinal bone growth in growth hormone (GH)-deficient hypophysectomized male and female rats to determine whether the effect of MH was similar to that of GH. We measured the nose-to-anus and nose-to-tail length gain, femur and tibia lengths, growth plate zones, and expression of insulin-like growth factor-1 (IGF-1) and IGF-binding protein-3 (IGFBP-3) after the dietary administration of MH or the injection of GH into hypophysectomized rats for 4 weeks. Results demonstrated that the dietary administration of MH had no effect on longitudinal bone growth, whereas the injection of GH increased the nose-to-tail length gain and femur and tibia lengths in hypophysectomized rats. In addition, MH did not affect the growth plate, bone mineralization, and expression of IGF-1 and IGFBP-3. These findings indicate that MH does not exert a GH-like effect and that the effects of MH and GH on longitudinal bone growth involve different pathways.
INTRODUCTION
Although several biochemical regulators contribute to longitudinal bone growth, growth hormone (GH) is the key peptide regulator that is responsible for this process. The GH exerts its function via an endocrine–paracrine–autocrine system and is secreted by the anterior pituitary gland in response to the GH-releasing hormone and hypophyseal-portal somatostatin. 1 The released GH binds to the GH receptor (GHR) in numerous cell types, leading to the production of insulin-like growth factor-1 (IGF-1). 2 Circulating IGF-1 binds to IGF-binding protein-3 (IGFBP-3) and acts as a regulator that contributes to longitudinal bone growth, a process that involves chondrogenesis. 3,4 Therefore, longitudinal bone growth failure is linked to decreased serum levels of IGF-1 and IGFBP-3, which are highly correlated with GH levels. 3 –6
The growth plate, populated by chondrocytes, is located between the epiphysis and metaphysis of long bones and consists of three distinct zones: the resting zone, proliferating zone, and hypertrophic zone. 7 These zones reflect the gradual transition of chondrocytes through the different stages of differentiation. The resting zone acts as a reserve of precursor cells for proliferating chondrocytes. The proliferating zone serves as a scaffold for bone elongation, whereas the hypertrophic zone is the site of the cells differentiating into hypertrophic chondrocytes. 7,8 Hypertrophic chondrocytes are specialized cells that are essential for bone growth via vascular invasion and recruitment of osteogenic cells, resulting in the replacement of cartilage. 9,10
Humulus japonicus, a commonly grown plant in Korea and China, belongs to the family Cannabaceae. Previous studies have shown that H. japonicus has antioxidant, anti-inflammatory, anticancer, and antibacterial properties and bioactive components, including terpenes, lupulons, phenols, and flavonoids. 11 –13 In a previous study, we demonstrated that the administration of a mixture of H. japonicus (MH; 300 mg/kg b.w.) increased the nose-to-tail length gain, lengths of femur and tibia, and growth plate height in Sprague Dawley (SD) rats. In addition, we found that MH stimulated the proliferation of chondrocytes within the growth plate, increased the serum levels of IGF-1 and IGFBP-3, and increased the expression of IGF-1 and IGFBP-3 mRNA in the liver and bone. 14 However, the mechanisms underlying these phenomena have not been elucidated. In this study, we investigated whether MH could enhance longitudinal bone growth via a mechanism similar to that of GH. To decipher the mechanism, we treated GH-deficient hypophysectomized rats with MH extract and investigated the effects on longitudinal bone growth.
MATERIALS AND METHODS
Extract preparation and animal experiments
The MH was prepared by using the method of Kim et al. 14 Hypophysectomized SD rats (3 weeks old, male and female) were purchased from Japan SLC, Inc. The experimental protocol was approved by the Animal Care and Use Review Committee of Kyung Hee University (KHGASP-19-312). The rats were housed in an environmentally controlled facility (22 ± 2°C, 50–60% humidity, and 12/12-h light/dark cycle) and fed an AIN 93G diet for an acclimation period of 7 days. The animals were randomly divided into five groups: sham-operated and on normal diet (Sham group); hypophysectomized and on normal diet (Control group); hypophysectomized and on normal diet, with intraperitoneal injection of 0.37 mg recombinant GH/kg b.w./day (GH group); hypophysectomized and on normal diet supplemented with 50 mg MH/kg b.w. (MH50 group); and hypophysectomized and on normal diet supplemented with 300 mg MH/kg b.w. (MH300 group). After 4 weeks, the rats were euthanized via cervical dislocation.
Micro-computerized tomography and analysis
Femur and tibia length, bone mineral density (BMD), trabecular number, bone volume/total volume, trabecular thickness, and trabecular separation were measured by using a Skyscan 1172® X-ray micro-computerized tomography scanning system (Bruker, Belgium).
Hematoxylin and eosin staining and immunohistochemical analysis
Tibiae were dissected out, fixed in 4% paraformaldehyde, and decalcified in 10% ethylenediamine tetraacetic acid. The samples were embedded in paraffin, cut into 5-μm-thick sections, and subjected to hematoxylin and eosin (H&E) staining or immunohistochemical analysis for IGF-1, IGFBP-3, and bone morphogenetic protein-2 (BMP-2).
Measurement of serum IGF-1 and IGFBP-3 levels
Serum IGF-1 and IGFBP-3 were determined by using IGF-1 and IGFBP-3 ELISA kits (R&D Systems, Inc., Minneapolis, USA).
mRNA extraction and reverse transcription-polymerase chain reaction
The mRNA from rat liver tissues was extracted by using an RNeasy Mini Kit (QIAGEN, Maryland, USA). complementary DNA (cDNA) synthesis was performed by using an iScript™ cDNA Synthesis Kit (Bio-Rad, Hercules, CA, USA) according to the manufacturer's direction. Real-time polymerase chain reaction was carried out by using a SYBR Green PCR Master Mix (Bio-Rad) with 1 μL synthetized cDNA. Next, we amplified the cDNA with 40 cycles of denaturation (95°C for 15 sec), annealing (58°C for 30 sec), and extension (72°C for 45 sec) by using the following primers: GAPDH forward primer 5′-TGG CCT CCA AGC AGT AAG AAA C-3′, reverse primer 5′-CAG CAA CTG AGG GCC TCT CT-3′; IGF-1 forward primer 5′-GCT ATG GCT CCA GCA TTC G-3′, reverse primer 5′-TCC GGA AGC AAC ACT CAT CC-3′; IGFBP-3 forward primer 5′-GGA AAG ACG ACG TGC ATT G-3′, reverse primer 5′-GCG TAT TTG AGC TCC ACG TT-3′. Data analysis was performed by using 7500 System SDS software version 1.3.1 (Applied Biosystems, Foster City, CA, USA).
Statistical analysis
Data are shown as means ± standard deviation. The data were statistically analyzed by one-way analysis of variance or Student's t-test in SPSS statistical procedures for Windows (SPSS PASW Statistic 23.0; SPSS, Inc., Chicago, IL, USA), and Duncan's multiple range test was used to determine the significance of differences in the data obtained. A P value less than .05 was considered significant.
RESULTS
MH did not affect longitudinal bone growth in hypophysectomized rats
We observed a significant decrease in the weight gain and nose-to-anus and nose-to-tail length gain in the hypophysectomized rats (Control) compared with those in the sham control (Sham) rats. At the same time, weight gain and nose-to-anus and nose-to-tail length gain were significantly increased in the GH group compared with those in the Control group. However, the dietary administration of 50 mg MH/kg b.w. and 300 mg MH/kg b.w. in the hypophysectomized rats did not affect the weight gain and the nose-to-anus and nose-to-tail length gains (P < .05) (Table 1). Further, on comparison with the Control group, there were no significant differences in the femur and tibia lengths of the hypophysectomized rats treated with 50 mg MH/kg b.w. and 300 mg MH/kg b.w., whereas there was a significant increase in the same parameters in the GH group (P < .05) (Fig. 1 and Table 2).
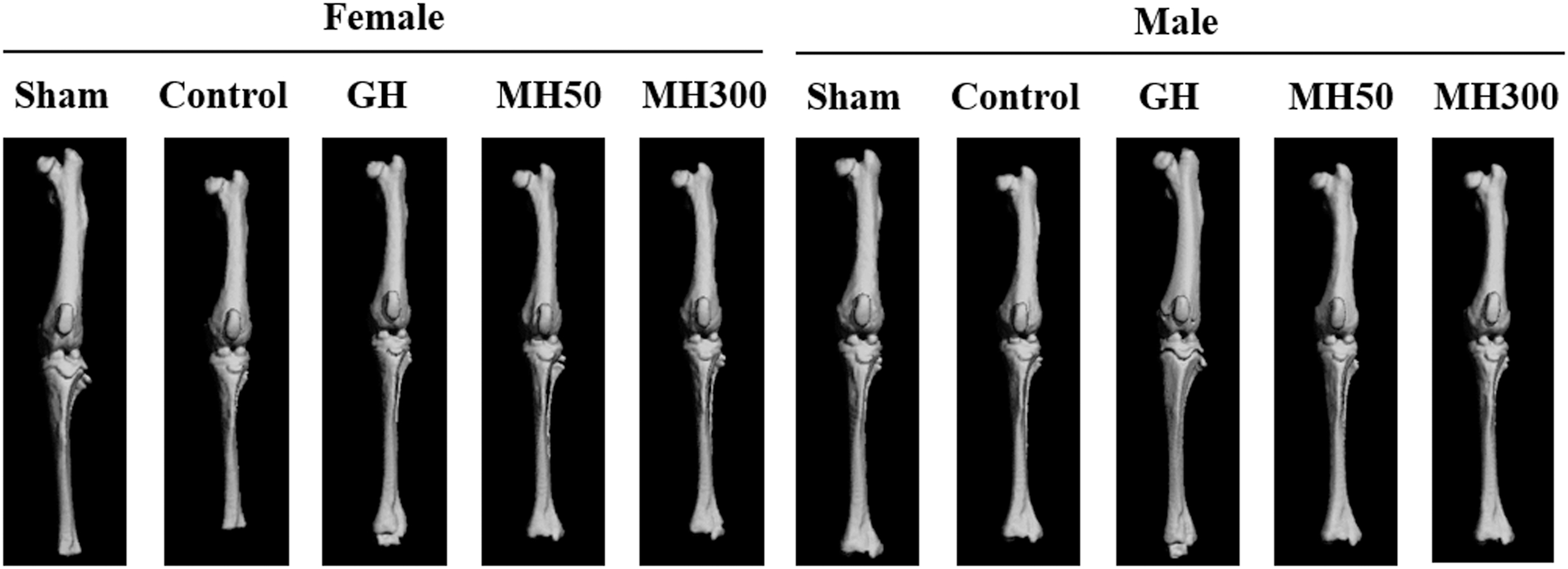
Effect of the dietary administration of MH on the femur and tibia lengths of hypophysectomized rats. Sham: sham-operated and normal diet; Control: hypophysectomy and normal diet; GH: hypophysectomy and normal diet with an intraperitoneal injection of 0.37 mg recombinant GH/kg b.w./day; MH50: hypophysectomy and normal diet supplemented with 50 mg MH/kg b.w.; MH300: hypophysectomy and normal diet supplemented with 300 mg MH/kg b.w. GH, growth hormone; MH, mixture of Humulus japonicus.
Effect of the Dietary Administration of Humulus japonicus Mixture on Weight Gain and Nose-to-Anus and Nose-to-Tail Length Gain in Hypophysectomized Rats
FER = [weight (g)/food intake (g)] × 100.
Sham: sham-operated and normal diet; Control: hypophysectomy and normal diet; GH: hypophysectomy and normal diet with an intraperitoneal injection of 0.37 mg recombinant GH/kg b.w./day; MH50: hypophysectomy and normal diet supplemented with 50 mg MH/kg b.w.; MH300: hypophysectomy and normal diet supplemented with 300 mg MH/kg b.w. Statistical values are presented as the means ± standard deviation. Different letters indicate a significant difference (P < .05) determined by using Duncan's multiple-range test.
FER, food efficiency rate; GH, growth hormone.
Effect of the Dietary Administration of Humulus japonicus Mixture on Femur and Tibia Length in Hypophysectomized Rats
Sham: sham-operated and normal diet; Control: hypophysectomy and normal diet; GH: hypophysectomy and normal diet with an intraperitoneal injection of 0.37 mg recombinant GH/kg b.w./day; MH50: hypophysectomy and normal diet supplemented with 50 mg MH/kg b.w.; MH300: hypophysectomy and normal diet supplemented with 300 mg MH/kg b.w. Statistical values are presented as the means ± standard deviation. Different letters indicate a significant difference (P < .05) determined by using Duncan's multiple-range test.
MH did not affect bone mineralization in hypophysectomized rats
Next, we investigated whether the dietary administration of MH affected bone mineralization in the hypophysectomized rats. The injection of GH in the hypophysectomized rats increased BMD, bone volume/total tissue volume, and trabecular number, thickness, and separation compared with those in the Control group (P < .05). However, the dietary administration of 50 mg MH/kg b.w. and 300 mg MH/kg b.w. did not have any effect on the BMD, bone volume/total tissue volume, and trabecular number, thickness, and separation in the hypophysectomized rats (Fig. 2 and Table 3).

Effect of the dietary administration of MH on the structure of the tibial trabecula and tibial cortex in hypophysectomized rats. Sham: sham-operated and normal diet; Control: hypophysectomy and normal diet; GH: hypophysectomy and normal diet with an intraperitoneal injection of 0.37 mg recombinant GH/kg b.w./day; MH50: hypophysectomy and normal diet supplemented with 50 mg MH/kg b.w.; MH300: hypophysectomy and normal diet supplemented with 300 mg MH/kg b.w.
Effect of the Dietary Administration of Humulus japonicus Mixture on Bone Mineralization in Hypophysectomized Rats
Sham: sham-operated and normal diet; Control: hypophysectomy and normal diet; GH: hypophysectomy and normal diet with an intraperitoneal injection of 0.37 mg recombinant GH/kg b.w./day; MH50: hypophysectomy and normal diet supplemented with 50 mg MH/kg b.w.; MH300: hypophysectomy and normal diet supplemented with 300 mg MH/kg b.w.; Statistical values are presented as the means ± standard deviation. Different letters indicate a significant difference (P < .05) determined by using Duncan's multiple-range test.
BMD, bone mineral density.
MH did not affect the growth plate in hypophysectomized rats
Further, we evaluated the effect of MH administration on the growth plates by using tissue sections stained with H&E (Fig. 3). Significant differences in the resting, proliferating, and hypertrophic zones of the growth plates were observed in the hypophysectomized rats injected with GH compared with those in the Control group (Table 4). However, in the MH50 and MH300 groups, there were no significant differences observed in the resting, proliferative, and hypertrophic zones of the growth plates (P < .05).
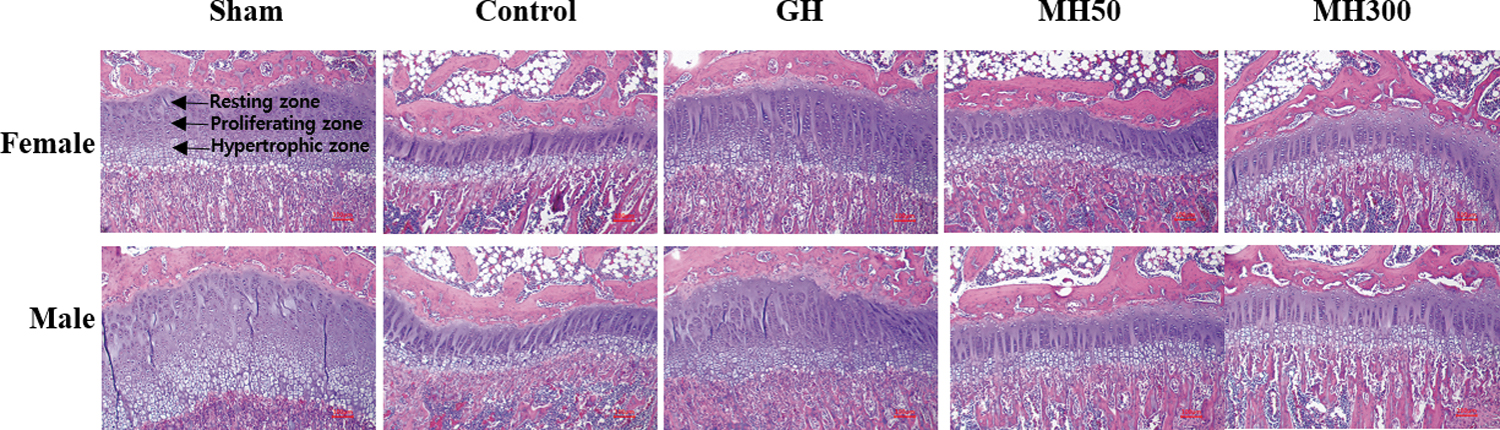
Effect of the dietary administration of MH on the growth plate of hypophysectomized rats. Sham: sham-operated and normal diet; Control: hypophysectomy and normal diet; GH: hypophysectomy and normal diet with an intraperitoneal injection of 0.37 mg recombinant GH/kg b.w./day; MH50: hypophysectomy and normal diet supplemented with 50 mg MH/kg b.w.; MH300: hypophysectomy and normal diet supplemented with 300 mg MH/kg b.w.
Effect of the Dietary Administration of Humulus japonicus Mixture on the Growth Plate in Hypophysectomized Rats
Sham: sham-operated and normal diet; Control: hypophysectomy and normal diet; GH: hypophysectomy and normal diet with an intraperitoneal injection of 0.37 mg recombinant GH/kg b.w./day; MH50: hypophysectomy and normal diet supplemented with 50 mg MH/kg b.w.; MH300: hypophysectomy and normal diet supplemented with 300 mg MH/kg b.w. Statistical values are presented as the means ± standard deviation. Different letters indicate a significant difference (P < .05) determined by using Duncan's multiple-range test.
MH did not affect IGF-1 and IGFBP-3 levels in hypophysectomized rats
To evaluate whether the dietary administration of MH affected the expression of IGF-1, IGFBP-3, and BMP-2 in the bone tissues of hypophysectomized rats, we performed immunohistochemical analysis. The expression levels of IGF-1, IGFBP-3, and BMP-2 in the bone tissues of the hypophysectomized rats injected with GH were increased compared with those in the Control group. However, there were no marked differences in the expression levels of IGF-1, IGFBP-3, and BMP-2 in the hypophysectomized rats administered 50 mg MH/kg b.w. and 300 mg MH/kg b.w., when compared with those in the Control group (Fig. 4).

Effect of the dietary administration of MH on IGF-1, IGFBP-3, and BMP-2 expression in the bone tissues of hypophysectomized rats. Sham: sham-operated and normal diet; Control: hypophysectomy and normal diet; GH: hypophysectomy and normal diet with an intraperitoneal injection of 0.37 mg recombinant GH/kg b.w./day; MH50: hypophysectomy and normal diet supplemented with 50 mg MH/kg b.w.; MH300: hypophysectomy and normal diet supplemented with 300 mg MH/kg b.w. BMP-2, bone morphogenetic protein-2; IGF-1, insulin-like growth factor-1; IGFBP-3, IGF-1 binds to IGF-binding protein-3.
Serum levels of IGF-1 and IGFBP-3 and the expression of their corresponding mRNA in liver tissues were significantly increased in the GH group but they showed no significant difference in the MH50 and MH300 groups (P < .05) compared with those in the Control group (Fig. 5).

Effect of the dietary administration of MH on serum levels of IGF-1
DISCUSSION
In a previous study, we investigated the effects of MH on longitudinal bone growth in SD rats to evaluate its potential as a food supplement. We observed that the administration of 300 mg MH/kg/b.w. increased femur and tibia bone length, chondrocyte proliferation, growth plate height, as well as bone mineralization in SD rats, suggesting that MH could effectively increase the longitudinal bone growth rate. 14 However, whether MH increased the longitudinal bone growth rate through a GH-like effect had not been elucidated. Therefore, in this study, we measured the nose-to-anus and nose-to-tail length gain after the injection of GH or the administration of MH in GH-deficient hypophysectomized rats.
The hypophysectomized rat model was used to investigate the involvement of GH and the effectiveness of a potential GH replacement. Our results demonstrated that the hypophysectomized rats exhibited a significantly decreased food efficiency rate (FER), nose-to-anus and nose-to-tail length gain, and femur and tibia lengths, compared with the sham control rats; however, the hypophysectomized rats that received the GH injection had significantly higher FER, nose-to-anus and nose-to-tail length gain, as well as femur and tibia lengths. These results indicated that the GH deficiency induced by hypophysectomy suppressed longitudinal bone growth, whereas GH replacement restored the longitudinal bone growth. Previous studies have also reported increased longitudinal bone growth after GH replacement in hypophysectomized rats. 14 In contrast, the dietary administration of MH in the hypophysectomized rats did not affect the FER, nose-to-anus and nose-to-tail length gain, and femur and tibia lengths. These results suggest that increased FER is one of the key factors mediating the effects of GH during longitudinal bone growth. Collectively, our current and previous findings indicate that MH does not act in a manner similar to GH, even though it can increase the longitudinal bone growth rate under certain conditions.
To evaluate the effect of dietary MH administration on the growth plate chondrocytes of the hypophysectomized rats, we performed H&E staining and quantified the differentiation zones of the growth plate, which is the area of the bone where longitudinal endochondral growth occurs. Several growth factors and hormones are known to act directly or indirectly on the growth plate. Further, the longitudinal bone growth rate is determined by the rate of chondrocyte proliferation and cartilage matrix secretion in the growth plate. 15 –17 We observed that the GH injection caused an increase in the resting, proliferating, and hypertrophic zones of the growth plate in the hypophysectomized rats; however, there were no significant differences in the three zones of the growth plate in the MH50 and MH300 groups compared with those in the Control group. In addition, the dietary administration of MH did not affect bone mineralization, BMD, bone volume/total tissue volume, and trabecular number, thickness, or separation in the hypophysectomized rats. Collectively, these results indicated that the dietary administration of MH did not affect the growth plates of the hypophysectomized rats.
Circulating pituitary GH binds to the GHR on the cell membrane and stimulates the production of IGF-1, which, in turn, contributes to chondrogenesis by binding to IGFBP-3. 18,19 We found that the injection of GH increased the expression and serum levels of IGF-1 and IGFBP-3 in the liver tissues of the hypophysectomized rats, whereas the dietary administration of MH did not. Previously, we demonstrated that the effect of MH on longitudinal bone growth in SD rats was mediated by increased JAK2/STAT5-induced IGF-1 production. 14 Our current results suggest that the effects of MH and GH on longitudinal bone growth are mediated via different pathways.
Kim et al. 20 reported that HT042, a herbal formula, promoted longitudinal bone growth and stimulated chondrocyte proliferation in adolescent SD rats. However, in another study, Kim et al. 21 demonstrated that HT042 did not affect longitudinal bone growth in spontaneous dwarf rats—a homozygous GH-deficient rat strain. The authors proposed that HT042 did not act in a manner similar to GH, even though it could increase the longitudinal bone growth rate in normal rats. Based on our previous and current findings, we suggest that MH increases the longitudinal bone growth rate but does not have a GH-like effect.
In conclusion, the dietary administration of MH had no effect on longitudinal bone growth in hypophysectomized rats, whereas the administration of GH resulted in increased nose-to-tail length gain and femur and tibia lengths. Further, the administration of MH did not affect the growth plate, bone mineralization, and expression of IGF-1 and IGFBP-3. These findings indicate that MH does not have a GH-like effect and that the effects of MH and GH on longitudinal bone growth are mediated via different pathways.
Footnotes
AUTHOR DISCLOSURE STATEMENT
No competing financial interests exist.
FUNDING INFORMATION
No funding was received.