
Abstract
In Mexico, Cnidoscolus aconitifolius (chaya) has been used to treat cardiovascular diseases (CVD). Because CVD are the number one cause of mortality, chaya use has become a health strategy. The aim of this study was to evaluate the antithrombotic activity and identify the metabolites in the most active extract. Aqueous (Aq), ethanolic (EtOH), acetonic (An), ethyl acetate (AcOEt), diethyl ether (Et2O), and hexanic (Hx) extracts were obtained. Platelet aggregation, phospholipase A2, prothrombin time (PT), activated partial thromboplastin time (aPTT), and clot lysis were evaluated. Metabolites were identified by gas chromatography–mass spectrometry (GC-MS). EtOH showed the greatest inhibition of platelet aggregation and phospholipase A2. Ac had the greatest effect on PT and aPTT. AcOEt had the greatest effect on clot lysis. EtOH, with the highest potential, was analyzed by GC-MS; fatty acids and triterpenes were identified. Thus, EtOH showed greater antiplatelet activity and other extracts showed moderate activity. This is a preliminary antithrombotic study. Future research will allow the development of nutraceuticals or functional ingredients for the prevention and treatment of thrombosis.
Introduction
At present, the largest contributor to global mortality is cardiovascular diseases (CVD), which cause ∼17.9 million deaths per year. 1 In Mexico, 23% of deaths are caused by these pathologies. Some factors for CVD development are the high consumption of ultra-processed foods, excessive alcohol intake, and physical inactivity. 2
Thromboembolic diseases are the most common cardiovascular disorders. They are characterized by the formation of intravascular clots (thrombi), which cause partial or total obstruction of blood flow. Formation of the thrombus is caused by endothelial dysfunction, blood flow disorders, and hypercoagulable states. These factors cause constant activation of platelet aggregation and blood coagulation. 3 Because of the constant activation of these hemostatic systems, fibrinolysis, a mechanism responsible for dissolving blood clots, is decreased. 4 To treat thrombosis, platelet aggregation, and phospholipase A2 inhibitors, anticoagulants and fibrinolytics, such as aspirin, anagrelide, heparin, and streptokinase, respectively, have been developed. However, numerous side effects have been reported for these drugs, gastrointestinal bleeding and hyperplasminemia being the most frequent. 5 Therefore, the research for natural alternatives has become a strategy for the prevention and treatment of CVD.
In traditional Mayan medicine, Cnidoscolus aconitifolius (Mill.) I.M. Johnst., commonly known as chaya, has been used as a traditional remedy for the treatment of diabetes, inflammation, and gastrointestinal disorders. This plant has also been used to enhance blood circulation. 6,7 Several studies have indicated that this plant has secondary metabolites with antithrombotic activity, such as ferulic acid, kaempferol, β-amyrin, and lupeol acetate. 8 –11 In the scientific literature it has been reported that chaya extracts exhibit a cardioprotective effect in CD1 mice induced to ischemia/reperfusion, an anticoagulant effect in diabetic rats induced with streptozotocin and a fibrinolytic effect in an in vitro model. 6,12,13 However, no studies focused on the antithrombotic activity of chaya. This study aims to evaluate the antithrombotic activity of chaya leaf extracts and to identify the major metabolites present in those extracts with greater biological activity.
Materials and Methods
Reagents and chemicals
Ethanol and acetone were purchased from Sigma-Aldrich (St. Louis, MO, USA). Ethyl acetate was purchased from Fermont (Monterrey, Nuevo León, Mexico). Diethyl ether and hexane were purchased from Merck Millipore (Burlington, MA, USA). All solvents were chemical grade. To determine the coagulation times prothrombin time (PT) and activated partial thromboplastin time (aPPT), the respective kits of the Stago Diagnostic brand (Parsippany-Troy Hills, NJ, USA) were used.
Plant material
C. aconitifolius leaves were collected in Timucuy, Yucatan State, Mexico (20° 48′ 43.4″ N, 89° 30′ 49.0″ W) in October 2017. The plant was identified by Dr. José Luis Tapia Muñoz and a specimen was deposited at Centro de Investigación Científica de Yucatán Herbarium (voucher registration number 69489).
Plant extraction
The chaya leaves were dried for 24 h in a convection oven, with air circulation, at 40°C, and pulverized in a Foss Cyclotec mill (ICSA, Valencia, Spain). The samples were stored in polyethylene bags at 24–26°C in darkness until the extraction was carried out. The samples were macerated separately with water, ethanol, acetone, ethyl acetate, diethyl ether, and hexane at a ratio of 1 : 10 (w/v) at 24–26°C for 48 h with constant stirring using a Thermolyne Big Bill orbital shaker (LabWrench, Toronto, Ontario, Canada) at 3 × g to produce Aq, EtOH, An, AcOEt, Et2O, and Hx extracts, respectively. The extracts were filtered using No. 50 filter paper. Finally, all extracts were concentrated under reduced pressure, using an RV10C rotary evaporator (IKA, Staufen, Breisgau, Germany). The resulting extracts were frozen and lyophilized in a FreeZone 2.5 L Benchtop Freeze Dryer (Labconco, Kansas City, MO, USA).
Blood collection
The study was conducted in accordance with the principles outlined in the Declaration of Helsinki. Blood samples were taken from donors with prior authorization. All the participants were healthy and they reported not having used any medicine 14 days before the blood sample was taken. Venous blood was removed from the antecubital vein using the vacutainer technique. Blood was placed in 4.5 mL vacutainer tubes with 0.109 M sodium citrate for later use in antithrombotic assays.
Antithrombotic activity in vitro
The dry plant material obtained from each extract was dissolved at final concentrations of 0.1, 1.0, and 10.0 mg/mL with 0.1% DMSO in physiological solution. Then, the in vitro antithrombotic activity of each solution was evaluated through platelet aggregation inhibition, PT, aPTT, and clot lysis assays.
Platelet aggregation inhibition
This assay measures the change in optical density of platelet-rich plasma (PRP) after adding an aggregating agent. As platelets aggregate, the passage of light through the plasma increases; in this way, the platelet aggregometer detects the increase in transmittance. Inhibition of platelet aggregation was determined according to the method proposed by Miyashita et al.
14
To obtain PRP, the vacutainer tubes with blood were centrifuged using a Solbat device (Proquisur, San Bartolome Xicomulco, CDMX, Mexico) at 136 × g for 15 min; the supernatant was removed and placed in a siliconized tube. The residual phase was centrifuged at 2050 × g for 15 min to obtain the platelet-poor plasma (PPP). Then, 250 μL of PPP were added to a glass cell and placed in the receptacle of a PAP-8E aggregometer (Sentinel Diagnostics, Milan, Italy) to adjust the 100% light transmission. The PRP was adjusted to 300 × 103 platelets per microliter using a KX-21 hematic cytometer (Sysmex, Kobe, Japan) and physiological solution as diluent. The PRP was readjusted to 200 × 103 platelets/μL by mixing 150 μL of 300 × 103 platelets/μL PRP with 75 μL of the respective extract in a glass cell with a magnetized bar in the aggregometer receptacle. The PRP was incubated for 3 min and after that time, 2 μL of an aqueous solution of 10 μM adenosine diphosphate was added, waiting 6 min before measurement. Graphs of the transmittance were produced by the computer program Start PAP-8E V2.1.0 (BioData Corp.). The percentage inhibition of platelet aggregation was calculated with the formula:
where:
%AS is the percentage of platelet aggregation of the sample.
%AC is the percentage of platelet aggregation of the negative control.
The negative control was a mixture of PRP with 0.1% DMSO in physiological solution (v/v) and the positive control was a mixture of PRP with 0.1, 1, and 10 mg/mL commercial aspirin in 0.1% DMSO in physiological solution (v/v).
Phospholipase A2 inhibition
This test measures phospholipase A2 action using Bothrops asper poison and erythrocyte solution. The enzyme causes the de-esterification of arachidonic acid from erythrocyte membranes. Thus, the blood cells release hemoglobin, which is quantified by spectrophotometry. Phospholipase A2 inhibition was determined according to the method proposed by Reddi et al.
15
Poison was obtained from a B. asper snake belonging to the Tsaab Kaan Wildlife Conservation Management Unit (registration SEMARNAT-UMA-IN-0183-YUC-10). Poison was lyophilized and suspended in 335,000 μg/mL phosphate buffer. Human erythrocytes were washed three times with phosphate buffer, suspended at 1.2 × 106 cells/μL and mixed with 1% (v/v) egg albumin. The resulting mixture was the substrate for the snake poison. Next, 100 μL of erythrocytes, 100 μL of 100 mg/mL venom, and 50 μL of the respective extract were mixed. The mixture was incubated for 30 min at 37°C, centrifuged at 1720 × g for 5 min, and the supernatant was placed in a 96-well plate. The absorbance was determined at 540 nm with an EL×800 plate reader (BioTek, Winooski, VT, USA). The percentage inhibition was calculated with the formula:
where Abs sample is the supernatant absorbance when the corresponding extract was added And Abs poison is the supernatant absorbance when the poison was added. The negative control was the erythrocyte and venom mixture with 0.1% DMSO (v/v).
Blood coagulation inhibition
The PT and aPTT tests were carried out following the methodology provided with the manufacturer's kit. The addition of prothrombin reagent to the blood plasma activates the extrinsic coagulation pathway through the formation of FT–FVII complex. To evaluate the effect of chaya extracts on the extrinsic coagulation pathway, 50 μL of PPP were mixed with 50 μL of the respective dissolved extract, and this mixture was incubated at 37°C for 1 min. Next, 100 μL of prothrombin reagent, consisting of tissue factor, was added and the time required to form a clot was measured with a Start coagulometer (Diagnostica Stago, Parsippany-Troy Hills, NJ, USA).
Adding the partially activated thromboplastin reagent to the blood plasma will activate the intrinsic coagulation pathway. To evaluate the effect of chaya extracts on the intrinsic coagulation pathway, 50 μL of PPP were mixed with 50 μL of the respective extract at the respective concentration (0.1, 1, or 10 mg/mL) and 100 μL of partially activated thromboplastin reagent was added. This mixture was incubated for 2 min. Finally, 100 μL of 25 mM calcium chloride was added, incubated for 15 sec, and the coagulation time was recorded as aPTT with a Stago coagulometer. The results were expressed in seconds.
For PT and aPTT assays, the negative control was plasma with 0.1% DMSO in physiological solution (v/v) and the positive control was plasma suspended in a vacutainer tube with 75 USP heparin.
Clot lysis
The activation of plasminogen to plasmin (active form) causes lysis of a clot by breaking down the fibrinogen and fibrin contained in the clot. This assay measures the extracts' ability to dissolve previously formed blood clots. The clot lysis test was performed using the method proposed by Prasad et al.
16
Venous blood (0.5 mL) was placed in preweighed microtubes and incubated for 45 min at 37°C in an Agl-12B water bath (GeoLab, Torreon, Coahuila, Mexico). After that, the tubes were centrifuged for 5 min at 168 × g in a Legend Micro 21R microcentrifuge (ThermoFisher Scientific, Waltham, MA, USA). The serum was removed, and the tube was weighed again. Subsequently, 100 μL of each extract at the respective concentration was added to an Eppendorf tube with a previously weighed clot. The tubes were incubated at 37°C for 90 min. Then, they were centrifuged for 5 min at 168 × g. The liberated liquid was removed, and the tubes were weighed again. The difference in weight before and after lysis of the clot was expressed as the percentage of clot lysis with the formula:
where P bl is the clot weight before lysis and P al is the clot weight after lysis. The negative control was the clot with 0.1% DMSO (v/v).
Identification of major metabolites in chaya extracts by gas chromatography–mass spectrometry
To identify the major metabolites, 0.3 μL of chaya extract was injected in splitless mode at 280°C. The separation was realized with a DB-5MS capillary column (30 m × 250 μm ID; 0.25 μm film thickness; 5% diphenylsiloxane and 95% dimethylsiloxane). The mobile phase was 1 mL/min helium. The oven temperature program initiated at 90°C. Then, the temperature was raised at 35°C/min up to 210°C and held for 2 min; then, the temperature was raised at 10°C/min to reach 270°C and held for 2 min; finally, the temperature was increased at 5°C/min to 300°C and held for 4 min.
The injector temperature was 280°C, whereas the transfer line and ion source of the mass spectrometer were set at 230°C and 1501°C, respectively. The ionization mode was electronic impact (70 eV). Ionic fragments were detected in scan mode (50–865 m/z). Spectral interpretation was carried out using the database NIST/EPA/NIH Mass Spectral Library Version 2.0.
The information detailed by the software included the name of each compound, along with various parameters that indicate the similarity between the sample spectrum and the database spectrum. The coincidence factor (CF) specified the degree of similarity of the unknown spectrum with the database spectra. Meanwhile, the inverse coincidence factor (ICF) referred to this similarity, ignoring the m/z unknown spectrum signals that did not match with the library spectra. Values >900 indicated a high match and values between 800 and 900 were used as good matches.
Statistical analysis
All experiments were performed in triplicate. The results for inhibition of platelet aggregation and phospholipase A2, PT, aPTT, and clot lysis tests were analyzed with a two-way ANOVA multifactorial model, to determine if there was significant statistical difference in the biological activity of the different dissolved extracts at the concentrations evaluated. Statistical assumptions were verified and P < .05 was considered as the level of significance. The data were processed using Statgraphics Centurion XVI® software.
Results
Platelet aggregation inhibition
The negative control inhibited platelet aggregation by 0.0%, indicating that the solvent used to solubilize the lyophilized extracts did not interfere in the biological process studied. At 0.1 mg/mL, the extracts showed no effect. At 1 mg/mL, the biological activity varied between 13% and 34%, corresponding to Hx and EtOH, respectively. The percentage inhibition of extracts at this concentration increased as follows: Hx<An<Aq = Et2O = AcOEt<EtOH, forming four groups according to their biological effect, which, as given in Figure 1, were significantly different from each other (P < .05). At 10 mg/mL, the biological activity varied between 0% and 100%, corresponding to Et2O and EtOH, respectively. The percentage inhibition of extracts at this concentration increased in the order: Et2O<AcOEt<Hx<An<Aq = EtOH, forming five groups according to their biological effect, which were significantly different from each other (P < .05). At this concentration, AcOEt had lowest percentage inhibition, because at 1 mg/mL, metabolites present in the said extract possibly saturated platelet enzymes. Therefore, increasing the concentration of the AcOEt did not influence the percentage inhibition of platelet aggregation. When evaluating the positive control, which consisted of a commercial solution of aspirin, at 0.1 mg/mL it demonstrated no inhibition of platelet aggregation. However, at 1 and 10 mg/mL it was shown to inhibit 100% of the biological activity. This shows that Aq and EtOH at 10 mg/mL have an inhibitory effect on platelet aggregation similar to that of the commercial drug aspirin.

Platelet aggregation inhibition of the different extracts at their respective concentrations. a–h Different lowercase letters indicate significant statistical difference between treatments (P < .05). n = 3. AcOEt, ethyl acetate; An, acetonic; Aq, aqueous; EtOH, ethanolic; Et2O, diethyl ether; Hx, hexane.
Phospholipase A2 inhibition
The negative control inhibited phospholipase A2 by 0.0%, indicating that the solvent used to solubilize the lyophilized extracts did not interfere in the biological process studied. At 0.1 and 1 mg/mL, no extract inhibited the poison's catalytic action. The negative percentages of the activity of the extracts at this concentration may be owing to the fact that some metabolite could be permeabilizing the erythrocyte membrane. This caused a greater release of hemoglobin compared with the negative control, which was expressed as negative percentages of biological activity. At 10 mg/mL, the biological activity varied between −22% and 44%, corresponding to Hx and EtOH, respectively. The percentage inhibition of extracts at this concentration increased as follows: Hx<An<AcOEt<Aq<Et2O<EtOH, forming six groups according to their biological effect, which, as given in Figure 2, were significantly different from each other (P < .05). For the positive control, the inhibition reported for quercetin was taken as a reference. 17 This flavonoid has been shown to inhibit 50% of phospholipase A2 activity at 0.813 μg/mL, which is lower than that reported for chaya extracts. This could be attributed to quercetin interacting directly with phospholipase A2, contrary to the complex mixture of metabolites present in the extracts.
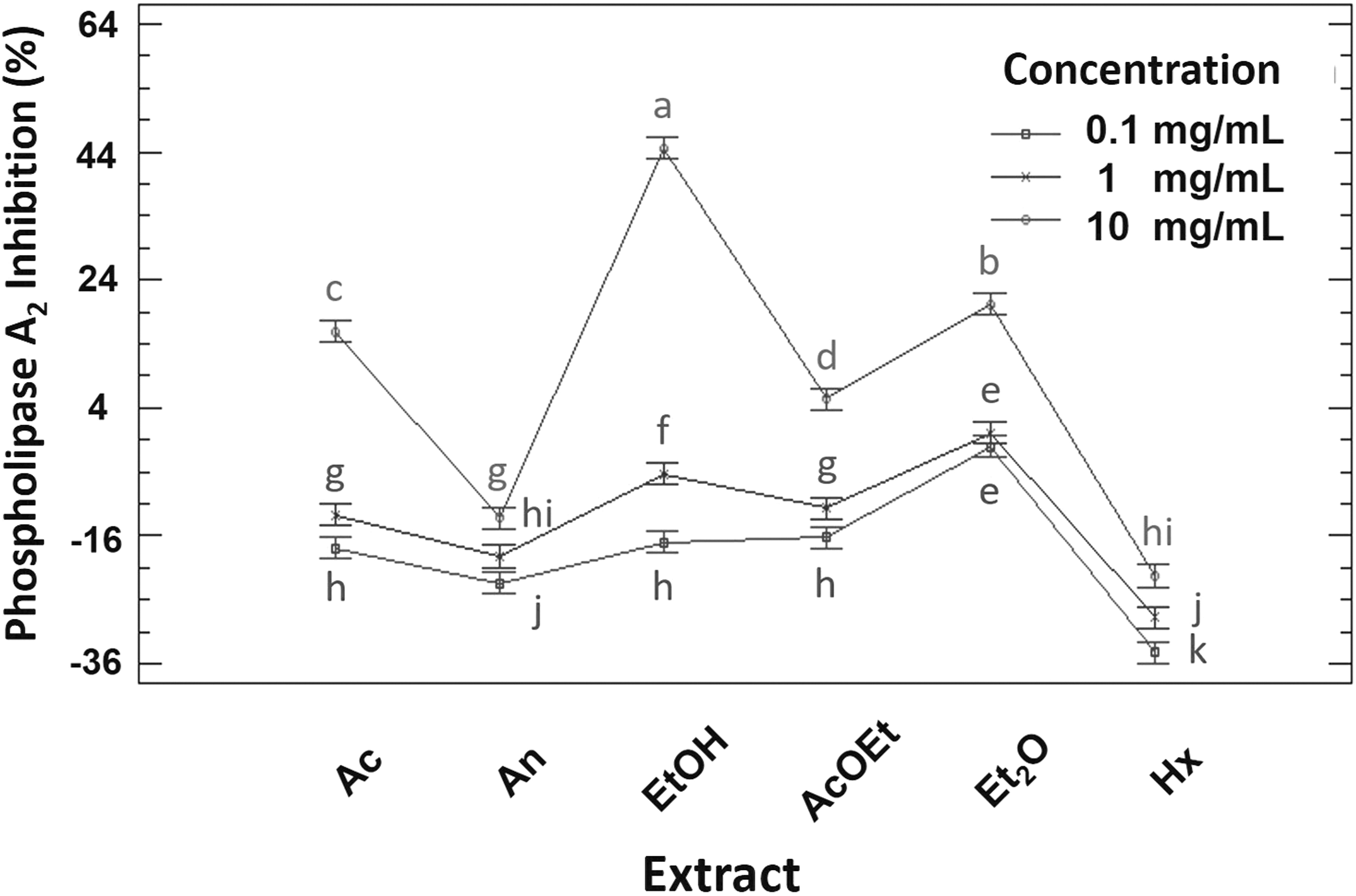
Phospholipase A2 inhibition of the different extracts at their respective concentrations. a–k Different lowercase letters indicate significant statistical difference between treatments (P < .05). n = 3.
Blood coagulation inhibition
For PT and aPTT, the negative control was evaluated; the coagulation times were 13.1 + 0.3 and 27.9 ± 0.7 sec, respectively, these times being normal in healthy humans. This indicates that the solvent used to dilute the extracts did not interfere in the biological effect studied. For PT, no extracts prolonged blood clotting at 0.1 and 1.0 mg/mL (Fig. 3). In comparison with the negative control and other extracts, Aq and An at 10 mg/mL prolonged the coagulation times by 5.8 ± 0.5 and 1.8 ± 0.36 sec, respectively (P < .05).
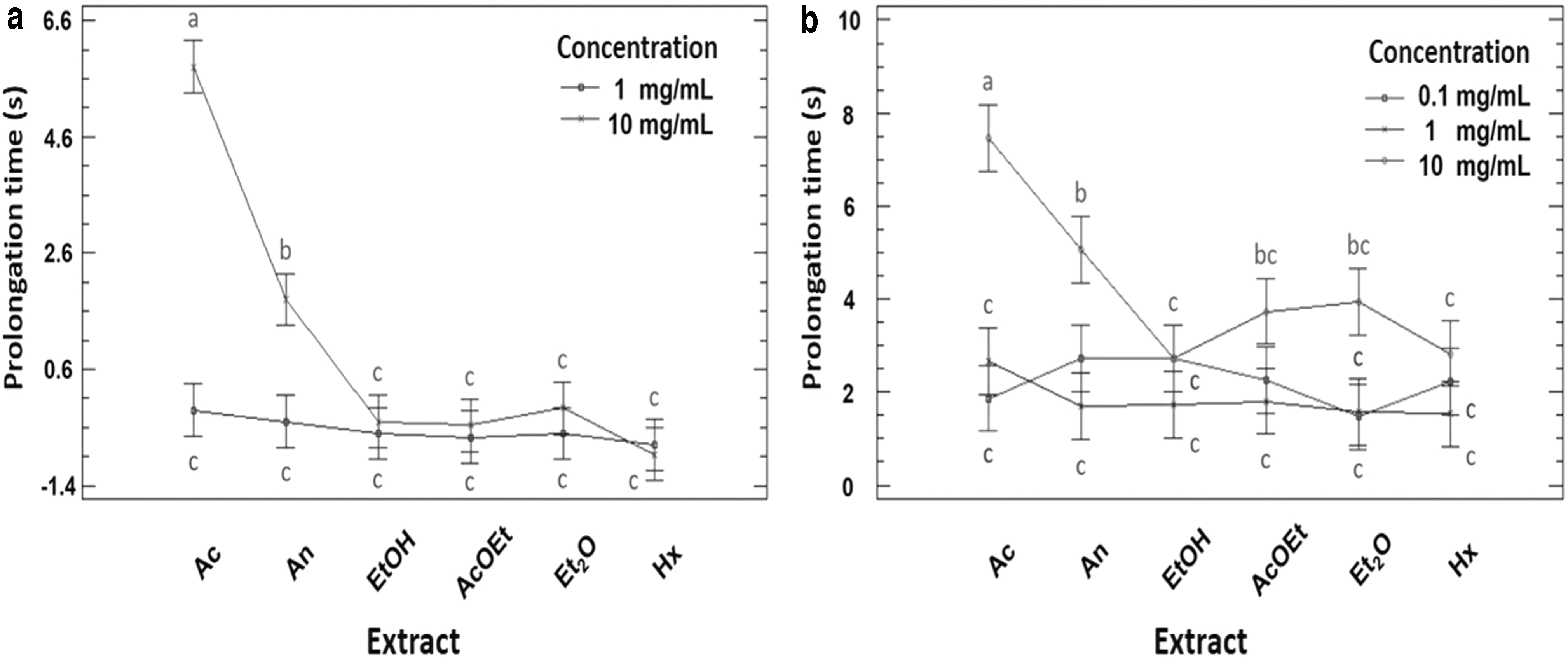
Blood coagulation inhibition owing to
For aPTT, there was no statistically significant difference (P < .05) in the anticoagulant effect of all extracts at 0.1 and 1.0 mg/mL; they prolonged coagulation between 1.5 and 2.7 sec. At 10 mg/mL, EtOH, AcOEt, Et2O, and Hx did not present statistically significant differences in comparison with the extracts at 0.1 and 1.0 mg/mL (Fig. 3). However, at this concentration, Aq and An extracts were significantly different from each other and from the rest of the extracts (P < .05), prolonging coagulation by 7.46 ± 0.80 and 5.06 ± 1.15 sec, respectively.
To evaluate the positive control, the blood was suspended in a 75 USP heparin tube. This plasma had a clotting time of 70.4 ± 0.3 and 96.9 ± 0.5 sec for PT and aPTT, respectively, indicating inhibition of the factors.
Clot lysis
The negative control (0.1% DMSO, v/v) showed 3.3% ± 0.0% clot lysis (Fig. 4). This reveals that the DMSO used to dissolve the extracts did not cause lysis of the clot. All the extracts showed a dose-dependent behavior for lysis of the clot, with a statistically significant difference between 1 and 10 mg/mL (Fig. 4). The exception was EtOH and Et2O, which did not show a statistically significant difference (P < .05) at the three concentrations. These extracts possibly reached their greatest biological effect at 0.1 mg/mL, achieving the maximum conversion of plasminogen to plasmin. Thus, the increment of concentration did not increase the biological activity. The homogeneous group constituted by AcOEt, An, and Hx at 10 mg/mL had the greatest fibrinolytic effect, varying from 18.4% to 24%. In contrast, streptokinase has been reported to have a lysing capacity of 76% at 30,000 IU/mL, the equivalent of 62.7 mg/mL. 16
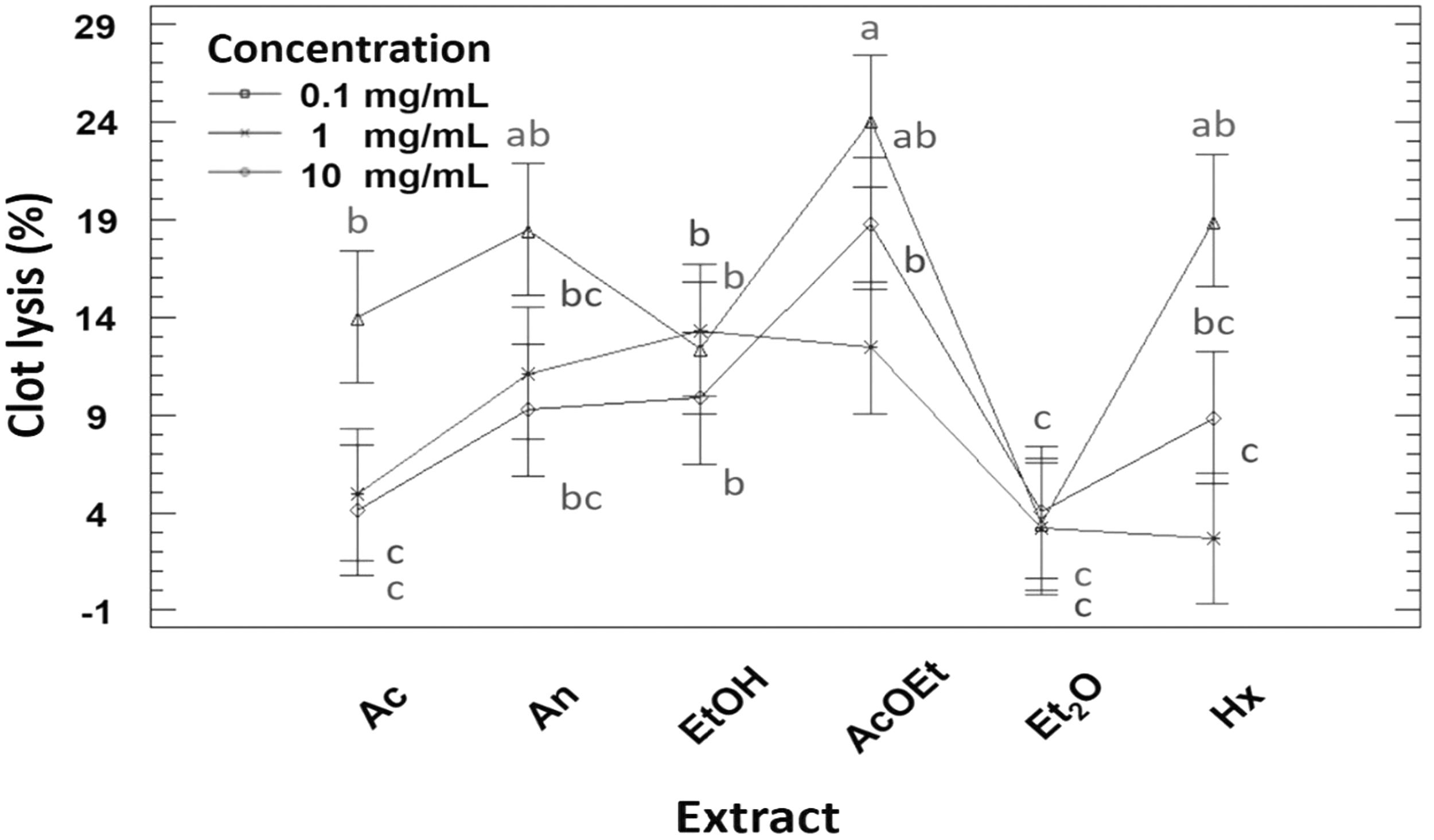
Clot lysis percentage of the different extracts at their respective concentrations. a–cDifferent lowercase letters indicate significant statistical difference between treatments (P < .05). n = 3.
Identification of major metabolites present in chaya extracts by GC-MS
The extracts with the highest biological activity were Aq and EtOH. Only EtOH was analyzed by gas chromatography–mass spectrometry (GC-MS) owing to the compatibility of the extract with the column stationary phase. Ten compounds were identified in EtOH. Table 1 shows the retention time (t R), the name of the metabolite, the coincidence factor, the inverse coincidence factor (CF/ICF), and the main ionic fragments for each compound. All experimental spectra showed high or good coincidence with the database spectra.
Major Metabolites Identified by Gas Chromatography-Mass Spectrometry in Ethanolic Extract
CF, coincidence factor; ICF, inverse coincidence factor.
In the spectrum of the peak with t R of 6.71 min, (1) a series of fragments that varied in 14 U from ion 73 to 227 m/z was observed. This fragmentation is characteristic of the loss of methylene groups [-CH2-] from aliphatic fatty acids. The molecular ion was observed at 256 m/z and the fragment at 239 m/z is related to the cleavage at the α-carbon of the carboxylic acid. The above data confirm the identity of palmitic acid. 18
The peak with t R of 7.10 min (2) reported a base ion at 88 m/z and a series of fragments that varied by 14 U from ion 101 to 255 m/z. This corresponds to the McLafferty rearrangement and the loss of methylene groups from long-chain ethyl esters, respectively. Fragments 284 and 239 m/z correspond to the molecular ion and the loss of the ethoxyl group [-O-CH2-CH3], respectively, confirming the identity of ethyl palmitate. 19
The mass spectrum of the peak with t R of 8.07 min (3) presented a base ion at 71 m/z, a molecular ion at 296 m/z, and some fragments such as 81, 95 and 123. Thes data correspond to the characteristic fragmentation of phytol. 20
The spectrum of the peak with t R of 8.32 min (4) presented a base ion at 79 m/z and a series of fragments that varied by 14 U from ion 93 to 163 m/z. Such fragments are characteristic of triene-interrupted methylenes. Fragments 108 and 222 m/z correspond to ω and α ions, which indicate a terminal n-3 double bond and a first double bond at position 8 from the carboxylic carbon, respectively. The above fragments, together with the molecular ion at 278 m/z confirm the identity of α-linolenic acid. 21
The mass spectrum of the compound with t R of 14.55 min (5) presented a base ion at 69 m/z and various fragments at 81, 95, 121, 137, and 149 m/z. The molecular ion was detected at 410 m/z. The previous data corresponded to squalene. 22
The mass spectrum of the compound with t R of 18.17 min (6) presented molecular and base ion at 430 and 165 m/z and various ions at 205, 177, and 121 m/z. These fragments have been reported by Bruinen et al., 23 who mentioned that it belongs to the specific fragmentation of α-tocopherol.
The mass spectrum of the compound with t R of 19.70 min (7) presented the molecular and base ion at 412 and 55 m/z, respectively. In addition, it presented various ions at 394, 351, 255 and 213 m/z, which, as reported by Suttiarporn et al. 24 belong to the tetracyclic triterpene stigmasterol.
The mass spectrum of the peak with t R of 20.93 min (8) presented a base ion at 218 m/z, characteristic of triterpenes with oleanan or ursan skeleton. The molecular ion at 426 m/z was observed, which suggested a molecular formula C30H50O. Because the 203 m/z ion is more than twice the intensity of the 189 m/z ion, according to Retro Diels-Alder fragmentation, this metabolite was identified as β-amyrin. 25
The mass spectra of the compounds with t R of 22.39 (9) and 23.10 min (10) presented as molecular ion 468 m/z that indicated a triterpene with a COOCH3 group at carbon 3. The first compound presented a base ion at 218, characteristic of amyrin esters, whereas the second, presented a base ion at 189, characteristic of lupane-type compounds. Therefore, both compounds were identified as amyrin acetate and lupeol acetate. 26
Discussion
Due to the side effects of antithrombotic drugs, natural alternatives have become a viable option to treat these complications. In traditional medicine, chaya has been used to improve blood circulation. There are studies where it has been reported that chaya has beneficial effects on the cardiovascular system. However, studies focused on the antithrombotic potential of chaya have not been reported. 6,12,13
Platelet–platelet interaction has the purpose of producing a platelet plug that will prevent the loss of blood fluid; however, factors such as the accumulation of lipids in the blood vessels constantly activate platelet aggregation, which can cause the formation of thrombi. 3 Thus, it is of interest to evaluate the antiplatelet activity of agents of natural origin. In the platelet aggregation inhibition assay, EtOH and Aq at 10 mg/mL demonstrated an antiplatelet effect similar to that of commercial aspirin solution, suggesting that these extracts possess metabolites with great antiplatelet potential. It has been reported that extracts from vegetables such as onion (Allium cepa L.), garlic (A. sativum L.), leek (A. ampeloprasum L.), and ginger (Zingiber officinale R.) at 1.5 mg/mL have an inhibitory effect on platelet aggregation, with values ranging from 0% to 17% against ADP agonist. 27 In the same way, it has been reported that a hexanic extract of olive tree leaves at 640 μg/mL showed 32% platelet inhibition against the ADP agent at 10 μM. 28 In this study, EtOH and Aq at 1 mg/mL showed 34.00% ± 2.65% and 26.67% ± 1.53% platelet inhibition, indicating that chaya extracts have a greater biological effect than onion, garlic, leek, and ginger but a lesser effect in comparison with hexanic olive leaves. This biological activity could be attributed to the synergy of the chaya metabolites. Among the metabolites present in chaya leaves and reported by Ramos-Gomez et al., 11 it has been shown that triterpene β-amyrin and the phenolic compounds quercetin and ferulic acid inhibit platelet aggregation. 9,29
Phospholipase A2 plays an important role in thrombosis, because it catalyzes the release of arachidonic acid from the platelet membrane, to be converted into thromboxane A2, a thrombotic agent. 3 The ethanolic extract at 10 mg/mL showed the greatest phospholipase A2 inhibition (44.65% ± 1.30%). Several studies have indicated that there is a correlation between the presence of phenolic compounds in plant extracts and the inhibition of phospholipase A2. EtOH had the greatest inhibitory effect because this solvent extracts a greater amount of phenolic compounds in comparison with acetone and water. 30 Finally, triterpenic compounds with oleanan-type skeleton, which were identified in this study as presented in Table 1, have been reported to inhibit the phospholipase A2 enzyme. 31
The inhibition of coagulation factors has been proposed as a strategy to combat thromboembolic diseases. The anticoagulant activity of chaya extracts was investigated by PT and aPTT assays. The first is used to evaluate the inhibition of coagulation factors II (tissue factor), V and VII, which are involved in the extrinsic pathway. The second is used to evaluate the inhibition of factors VIII, IX, X, XII, and prekallikrein, which are involved in the intrinsic pathway. 32 As for platelet aggregation inhibition, Aq and EtOH were the extracts that had the greatest anticoagulant effect in the PT and aPTT tests. Comparing the anticoagulant effect of chaya extract with that of other vegetables, it has been reported that beans (Phaseolus vulgaris L.), squash (Cucurbita moschata D.), broccoli (Brassica oleracea var. italic), carrots [Daucus carota subsp. sativus (Hoffm.) Arcang.], cauliflower (B. oleracea var. botrytis L.), and spinach (Spinacia oleracea L.) extracts do not inhibit blood clotting in PT and aPTT tests. 33 Among the metabolites identified in chaya leaves, β-amyrin has been shown to inhibit tissue factor expression, resulting in inhibition of the extrinsic coagulation pathway. 34 Similarly, β-sitosterol, a triterpene identified in chaya leaves, has been reported to prolong blood coagulation in vitro in human PPP by 28 and 62 sec, for PT and aPTT, respectively. 35 The presence of these metabolites in chaya extracts could be influencing their anticoagulant activity.
A reduction in fibrinolysis owing to a highly thrombotic environment can cause pathological problems. 4 Thrombolytic drugs activate plasminogen to plasmin, which lyses a clot by breaking down the fibrin contained in the clot. Commercial thrombolytic drugs, such as streptokinase, act nonselectively, activating plasminogen both in clots and in surrounding circulation, thus causing problems such as uncontrolled bleeding. The use of plants as fibrinolytic agents could be an alternative, so the fibrinolytic activity of chaya leaf extracts was investigated by a clot lysis assay. It has been reported that a pentacyclic triterpene compound of the isohopane type was able to positively regulate the ratio of the enzymes involved in the fibrinolytic system, such as tissue plasminogen activator (t-PA) and its inhibitor (PAI-1). 36 In this study, various triterpenic compounds with a pentacyclic skeleton were identified in the ethanolic extract, which could be participating in fibrinolytic activity. In addition, the triterpene β-sitosterol, with steroid type skeleton has been shown to cause lysis of 7.7 mg of a goat blood clot, compared with streptokinase, which demonstrated lysis of 15.5 mg. 35 In this study, the steroidal triterpene compound stigmasterol was identified, a metabolite that could also be participating in fibrinolytic activity. In contrast with our results, another study reported that extracts obtained from onion (A. cepa L.), spinach (S. oleracea L.), cauliflower (B. oleracea var. botrytis L.), and tomato (Solanum lycopersicum L.), among others, were not able to produce fibrinolysis in a euglobulin lysis time assay. 33 This indicates that those plant extracts did not activate the plasminogen among the metabolites identified in chaya leaves.
Because the ethanolic extract had the highest biological activity in most tests, it was analyzed by GC-MS. This analysis reported the presence of palmitic acid, ethyl palmitate, and linolenic acid in the extract. Palmitic acid has been shown to inhibit the activity of phospholipase A2, 37 whereas ethyl palmitate has been reported to decrease the expression of the enzyme cyclooxygenase 2. 38 On the contrary, the consumption of unsaturated fatty acids has been associated with a decrease in the incidence of myocardial infarction and stroke. 39
Phytol, squalene, and vitamin E were identified in the ethanolic extract. No antithrombotic effects have been reported for phytol, whereas squalene has been reported to stabilize cardiac tissue membrane in a model of rats induced to heart attack by the drug isoprenaline. 40 Vitamin E has been demonstrated to have antiplatelet effects through four mechanisms: (1) protein kinase C inhibition, (2) γ-carboxylase inhibition, (3) arachidonic acid pathway inhibition, and (4) the reduction of platelet adhesion. 41 The above shows the antiplatelet activity reported in this study. Finally, the ethanolic extract presented stigmasterol, β-amyrin, β-amyrin acetate, and lupeol acetate. In general, these triterpenes have been reported to inhibit the catalytic action of cyclooxygenase 2 by hydrophobic interaction. 42 Furthermore, the pentacyclic triterpene β-amyrin has been shown to inhibit platelet aggregation in human blood plasma against the ADP agonist agent, demonstrating its antagonistic effect on the platelet receptors P2Y1 and P2Y12. 9 In this way, the effects of chaya reported in traditional Mayan medicine can be attributed to the secondary metabolites it contains, such as the terpenes reported here; this study being a first approach to the antithrombotic potential of this plant. It would be interesting to analyze the chemical composition of the ethanol extract using techniques that allow the identification of nonvolatile polar compounds, such as liquid chromatography–mass spectrometry.
In conclusion, in this study the antithrombotic activity of different chaya extracts was evaluated. Chaya extracts were able to inhibit ADP-induced platelet aggregation, Aq and EtOH being those with an effect similar to that reported for aspirin. In relation to phospholipase A2 inhibition, again EtOH was the one that presented the greatest enzymatic inhibition. EtOH and Aq prolonged blood coagulation because of both coagulation pathways. For fibrinolytic activity, AcOEt demonstrated the greatest potential. EtOH was analyzed by GC-MS because it showed high inhibition in most of the tests. This extract was found to contain various unsaturated and saturated fatty acids with a protective effect against CVD. It also contains vitamin E, a metabolite that has been shown to inhibit platelet aggregation by various routes. Finally, EtOH contains various triterpenes, which have been demonstrated to have anticoagulant effects and act as inhibitors of platelet aggregation. Thus, chaya extracts, especially EtOH, presented high antithrombotic potential, which may be associated with the metabolites they contain. This is a preliminary study on the antithrombotic potential of chaya. Thus, future research is necessary to elucidate bioactive compounds with antithrombotic potential and to develop nutraceuticals or functional ingredients for the prevention and treatment of thrombosis.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
The authors thank CONACYT for providing scholarship 632866 to J.P.Q.-M. and Kellogg's foundation for project P3037075.