
Abstract
Group A Streptococci (GAS) or Streptococcus pyogenes is responsible for acute bacterial pharyngitis in children as well as adults. Streptococcal pharyngitis is initiated by successful attachment and colonization of the bacteria, followed by the establishment of the biofilm in various environments. In this study, we examined the antibacterial activities of in-house prepared aqueous and ethanolic extracts of 10 Atlantic Canada fruits in the context of minimum inhibitory concentration (MIC), minimum bactericidal concentration (MBC), time–kill kinetics, and adhesion inhibition properties against S. pyogenes. Per our findings, MIC and MBC for all the tested extracts ranged from 0.25 to 8 mg/mL and from 4 to 64 mg/mL, respectively. Accordingly, at 1⁄2 × MBC, cranberry and sumac extracts also lowered the attachment of GAS to the uncoated and fibronectin-coated substratum. Particularly, cranberry and sumac aqueous extracts were more effective against the adhesion of S. pyogenes ATCC 19615 to the fibronectin-coated surface than a clinical strain. In conclusion, ethanolic and aqueous extracts of cranberry and sumac could potentially be incorporated into natural health products designed for the amelioration of strep throat, yet a detailed understanding of its mode of action (e.g., biofilm inhibition and eradication) could pave its path to the field of antibacterial natural health product discovery, design, and development.
Introduction
Group A Streptococci (GAS) could cause infections ranging from non-life-threatening conditions such as mild skin infection or pharyngitis to life-threatening and severe conditions such as necrotizing fasciitis, rheumatic fever, and lethal streptococcal toxic shock syndrome. Pharyngitis is the most common type of GAS infection. Streptococcal pharyngitis results from the proliferation of GAS in the pharynx. 1 Virulence-associated factors enable Streptococcus pyogenes to attach to host tissues, evade the immune response, and spread by penetrating the host tissue layers. 2 –4 After successful attachment, bacteria aggregate, multiply, and form microcolonies that differentiate into exopolysaccharide-encased communities called mature biofilm. Manetti et al. have proposed the role of biofilm in the pathogenesis of GAS more than a decade ago. 5
Folklore medicinal plants have long been used, in the past and the present, for the treatment of S. pyogenes infections, including pharyngitis. 6,7 Recent literature has shown that phytochemicals, particularly polyphenols, exhibit good antibacterial activity against different strains of GAS. Phenolics and polyphenol-rich extracts of many plants, including maracuja (Passiflora foetida L.), white weed (Ageratum conyzoides L.), calabash tree (Crescentia cujete L.), bush banana (Uvaria chamae P. Beauv), ginger (Zingiber officinale Roscoe), bitter kola (Garcinia kola Heckel), and little gourd (Coccinia grandis L. Voigt), have shown anti-GAS activity. 6 –9 Berries are a good source of polyphenols, especially anthocyanins, which have been shown to possess noticeable antimicrobial activities. 9
In this study, we prepared aqueous and ethanolic extracts of 10 selected berries commonly found in Atlantic Canada for antibacterial screening. Specific objectives of this study were as follows: (i) screening the fruit extracts for their antimicrobial activity against S. pyogenes and (ii) investigating their mechanism of action in terms of anti-adhesive attributes.
Materials and Methods
Chemicals and reagents
Dimethyl sulfoxide, penicillin G sodium salt, sodium chloride, and Triton X-100 were purchased from Sigma–Aldrich Ltd. (Oakville, ON, Canada). MTT [3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide] was purchased from Life Technologies (Burlington, ON, Canada). Bovine serum albumin (BSA), ethanol, glycerol, human fibronectin, and phosphate-buffered saline (PBS) were purchased from Fisher Scientific (Ottawa, ON, Canada). Bacteriological agar and brain heart infusion (BHI) broth were purchased from Oxoid Ltd. (Nepean, ON, Canada).
Plant materials
The source of berries is given below (Global Positioning System [GPS] locations are only noted for locally collected berries). Crowberry (Empetrum nigrum L.) and squashberry [Viburnum edule (Michx.) Raf.] were provided by Dark Tickle Company (Saint Lunaire-Griquet, NL, Canada). Common buckthorn (Rhamnus cathartica L.) was harvested the first week of September 2014 at 40 Cox Road, Truro, NS, Canada (GPS location of 45°22′25.3″N 63°15′49.7″W). Mountain ash or rowanberry [Sorbus decora (Sarg.) C.K. Schneid.] was harvested the third week of September 2014 at 69 Pictou Road, Truro, NS, Canada (GPS location of 45°22′32.2″N 63°15′51.9″W). Sumac or staghorn (Rhus typhina L.) was harvested the fourth week of September 2014 at 221 Farnham Road, Truro, NS, Canada (GPS location of 45°23′00.4″N 63°15′52.1″W).
Cultivated berries were collected in the following order with their location (if applicable): blackcurrant (Ribes nigrum L. cv. Titania), provided by a Prince Edward Island commercial grower, wild-type blueberry (Vaccinium corymbosum L.), university experimental plot (Debert, NS, Canada), cranberry (Vaccinium macrocarpon Aiton), commercial farm of Cranberry Acres Ltd. (Berwick, NS, Canada), honeysuckle (Lonicera caerulea L., variety of Borealis), commercial farm of LaHave Natural Farms (Blockhouse, NS, Canada), and partridgeberry (Vaccinium vitis-idaea L., wild type) (Southern Labrador, NL, Canada). All berries were stored at −20°C until further use.
Bacterial strains and growth conditions
S. pyogenes ATCC 19615, ATCC 49399, and a clinical strain were included in the study. S. pyogenes ATCC 19615 and ATCC 49399, purchased from American Type Culture Collection (ATCC, Manassas, VA, USA), were isolated from the pharynx of a child following an episode of sore throat and human throat, correspondingly. The clinical strain was kindly provided by Dr. R. J. Davidson's Bacteriology Laboratory, Division of Microbiology, Pathology, and Laboratory Medicine, Nova Scotia Health Authority, Halifax, NS, Canada (isolated from a positive strep throat patient). Bacteria were stocked in BHI broth containing 20% glycerol at −50°C. Bacteria were routinely grown in BHI broth and were statically incubated overnight at 37°C.
Preparation of bacterial cultures
BHI broth (10 mL) was inoculated with 10 μL of bacterial stock. After overnight incubation at 37°C, the culture was vortexed, and its optical density (OD) was read at 600 nm in a microplate reader (Epoch™; BioTek, Winooski, VT, USA). Cells from 1.5 mL of the overnight culture were pelleted by centrifugation (13,000 g for 10 min) and resuspended in BHI broth to the concentration of 9 log colony-forming units (CFU) per milliliter. Separate experiments showed that one OD600 nm unit equals 9 log CFU/mL. Serial 10-fold dilutions were made from the bacterial suspension, and the number of bacterial cells was routinely confirmed by plating onto BHI agar.
Preparation of phytochemical-rich extracts
The extracts were prepared according to a previously described method with modifications. 10 In this study, two types of extracts were prepared using two green solvents (ethanol or deionized water). One hundred grams of berries was weighed (in triplicates), subsequently homogenized, and blended (with 100 mL of 95% ethanol or deionized water for 5 min using ethanol-resistant commercial blenders) to smooth consistency. The homogenized berries were filtered through Whatman No. 5 filter papers. The residue consecutively was re-extracted with another 100 mL of solvent and filtered. Filtrates were combined (to achieve a homogeneous product) and stored at 4°C.
The ethanolic extracts were partially dried using a rotary evaporator (Model Rota Chill LM6; Heidolph, Elk Grove Village, IL, USA) under reduced pressure at 20°C to obtain the crude extract. The extracts were later lyophilized (Dura-Stop MP; FTS Systems, Philadelphia, PA, USA) under 3600 mT vacuum, first 36 h at −20°C and then for 14 h at 20°C. The final products were stored at −80°C until further analysis. All extracts were prepared as described above except sumac, which was extracted with a berry:solvent ratio of 1:4 because of low moisture content of the fruit. The filter sterilization method was used to remove the contaminating microbes (if any) from the extracts. Decontamination was performed after the dissolution of the crude extracts with the appropriate solvent before each assay.
Determination of minimum inhibitory concentration and minimum bactericidal concentration of the extracts against S. pyogenes
Minimum inhibitory concentration (MIC) and minimum bactericidal concentration (MBC) were determined using the microbroth dilution method as described by the Clinical Laboratory Standards Institute (CLSI). 11 Stock solutions of the extracts/compound were prepared using sterile distilled water. To 100 μL of the diluted extract, 100 μL of bacteria was added (∼5.7 log CFU/mL). The 96-well plate was incubated at 37°C for 24 h. Sterile distilled water was used as solvent control. Wells without extract (bacteria only) served as growth (population) control.
The background color was subtracted from the readings (wells with extracts and without bacterial inoculum). Readings were taken at 600 nm with a microplate reader. MIC is defined as the lowest concentration of the extract inhibiting bacterial growth. Samples (30 μL) from wells with no visible growth were plated onto BHI agar plates to determine MBC values (no colony growth on the plate). This value is defined as the lowest concentrations at which 99.9% of the bacteria are killed.
Antimicrobial efficacy testing (time–kill kinetics)
Standardized bacterial cultures were incubated with 1⁄2 × MIC to 8 × MIC concentrations of the extracts/compound at 37°C. At selected time points, 100 μL of the cultures was 10-fold diluted with sterile saline water and plated onto BHI agar (three replicates). Solvent control was included in the assay. BHI broth media with and without bacterial culture were set as positive and negative bacterial growth controls, respectively.
Adherence inhibition assays
Anti-adherence activity of selected extracts against attachment of S. pyogenes to uncoated surfaces
The assay was conducted as described by others with slight modifications. 12 The uncoated 96-well polystyrene microplate was incubated at 37°C for 1 h with the bacterial inoculum (approximate bacterial density of 5.7–6.0 log CFU/mL) and different concentrations of the test material (equal or less than MBC). After 1 h of treatment, loosely attached bacteria were discarded, and adherent bacteria were recovered with 100 μL of BHI (vigorous pipetting technique).
Negative and positive controls (BHI broth with and without bacterial culture) were also designed for the assay. Only two solvent controls parallel to the two highest concentrations of each test material were tested. Solvent controls were as follows: for cranberry aqueous extract (8–16 mg/mL) 6.25–12.5% of H2O, for sumac aqueous extract (4–8 mg/mL) 3.13–6.25% of H2O, and for penicillin G (8–16 ng/mL) 0.01–0.02% of H2O. Subsequently, serial dilutions (10-fold) of the adherent bacteria were plated onto BHI agar plates for CFU/mL enumeration.
Anti-adherence activity of selected extracts against attachment of S. pyogenes to protein-coated surfaces
The assay was conducted as described in the preceding section. Briefly, wells of polystyrene microplates were coated with fibronectin (in PBS, 10 μg/mL). To each well, 100 μL of the fibronectin solution was added, and then, the plate was incubated at room temperature for 2 h. Following incubation, the content was discarded and gently washed with 150 μL of PBS.
To block the fibronectin-coated wells, BSA (in PBS, 1%) in the volume of 120 μL was added and incubated at room temperature for 1 h (for accurate anti-adhesive activity measurements, BSA was used to block the spots not occupied by fibronectin, thus decreasing the nonspecific bindings). Fibronectin-coated and BSA-blocked wells were washed with 150 μL of PBS three times. The rest of the procedure was like the one mentioned in the preceding protocol. Percent adherence of the bacteria to the proteins (BSA and fibronectin) relative to the initial inoculum was calculated.
The background adhesion was also taken into account by blocking the wells with BSA only (no fibronectin coating). Subsequently, wells were treated in the same manner to quantify the adherence of bacteria to BSA via nonspecific binding proteins. Moreover, this demonstrated the sole adherence of bacteria to fibronectin through fibronectin-binding proteins only versus nonspecific binding through other adhesins.
Anti-adherence activity of selected extracts against attachment of S. pyogenes to epithelial cells
The assay was conducted according to a previously described method with slight modification. 13,14 HTEpiC (human tonsil epithelial cells) cells (isolated from human normal tonsil tissue, purchased from ScienCell Research Laboratories [catalog No. 3220]) were maintained as per the manufacturer's instructions. The cells were seeded in the wells of 96-well plates (cell density of 25,000 cells per well and the volume of 250 μL) and incubated for 24 h at 37°C, 5% carbon dioxide, and 95% air. Cells were then washed with 250 μL of PBS twice.
Equal volumes of test compound at different concentrations (2–4 mg/mL for cranberry aqueous extract and 8–16 ng/mL for penicillin G) and bacterial inoculum (approximate density of 6.4–6.5 log CFU/mL for S. pyogenes ATCC 19615 and clinical strain) were mixed and then incubated at room temperature for 10 min. Concentrations tested in the assay were selected based on the results of the cell cytotoxicity assay. Bacterial cultures were prepared as described earlier with slight modifications. Bacteria were diluted with antibiotic-free cell culture media. Two hundred microliters of the mixture was added to each well and incubated for 1 h. The ratio of the tonsil cells to the bacteria was ∼1:100.
Weakly adhered bacteria were discarded, and 100 μL of trypsin (1% in PBS) was added and incubated for 10 min at room temperature. One hundred microliters of Triton X-100 (0.001% in PBS) was added to each well and incubated at room temperature for a couple of minutes. BHI broth with and without bacterial culture was considered positive and negative controls. Solvent controls were as follows: 0.01–0.02% of H2O for 8–16 ng/mL of penicillin and 1.56–3.13% of H2O for 2–4 mg/mL of cranberry aqueous extract. Later, serial dilutions (10-fold) of the adherent bacteria were plated onto BHI agar plates for CFU/mL enumeration. Percent adherence of the bacterial cells to the tonsil cells relative to the positive control (control: 100%) was calculated.
Cell viability assay
Cells were seeded in the 96-well plate with a density of ∼5 × 103 cells per well. Cranberry aqueous extracts at a concentration ranging from 2 to 16 mg/mL were added to the wells (in triplicates). The plate was incubated for 1 h at 37°C. Thereafter, 100 μL of media was added to the wells following the aspiration of their content. Ten microliters of freshly prepared MTT (5 mg/mL dissolved in PBS) was added to each well, and then, the plate was incubated for 2–4 h at 37°C in the dark. After purple color development, absorbance was read at 540 nm. Percent cell viability was calculated by the following formula:
where A is the absorbance read at 540 nm.
Statistical analysis
Experiments were performed as three independent studies, each in triplicate. Student's t-test (Excel, Microsoft Office 365) was used for statistical analysis of MIC values assessing the significant differences between the solvent control and the extract/compound treatments (P ≤ .05). Statistical significance of means (solvent control, positive control, and the treatment) was assessed by one-way analysis of variance; Tukey's multiple comparisons (P ≤ .05) using Minitab 16.1. Correlation analysis was performed using the same version of Minitab.
Results and Discussion
MIC and MBC of extracts against S. pyogenes
The results of MIC analysis showed that 9 of the 10 berry extracts tested were active against S. pyogenes strains, with sumac being the most active one (Table 1). The MBC for tested extracts ranged from 4 to 64 mg/mL (Table 1). The aqueous and ethanolic extracts of the berries were equally potent in killing the tested S. pyogenes strains. All the cultivated berries, excluding blueberry (MBC of ≥64 mg/mL), were relatively equally effective against S. pyogenes (MBC of 8–32 mg/mL). Among the wild-grown berries, sumac berry extracts were the most potent (MBC of 4–8 mg/mL).
Minimum Inhibitory Concentration and Minimum Bactericidal Concentration of Aqueous and Ethanolic Extracts of Selected Atlantic Canada Fruits Against Streptococcus pyogenes by Microbroth Dilution Method
MIC values for penicillin G (antibiotic control) are 2, 4, and 2 ng/mL for S. pyogenes ATCC 19615, ATCC 49399, and clinical strain, respectively. Student's t-test was used to determine the significant differences between the control and the extract/compound treatment (P ≤ .05). MBC for penicillin G (antibiotic control) are 16, 64, and 16 ng/mL for S. pyogenes ATCC 19615, ATCC 49399, and clinical strain, respectively.
19615, S. pyogenes ATCC 19615; 49399, S. pyogenes ATCC 49399; ATCC, American Type Culture Collection; Clinical, a clinical strain of S. pyogenes isolated from a patient; MBC, minimum bactericidal concentration; MIC, minimum inhibitory concentration.
In a study by Nasar-Abbas and Halkman, Sicilian sumac (Rhus coriaria L.) extracts showed inhibitory effects against Gram-positive and Gram-negative bacteria. 15 Per their report, MIC against Gram-positives, such as Bacillus species and Listeria monocytogenes, ranged from 0.25 mg/mL to 32 mg/mL (w/v) and 0.67 mg/mL (w/v), respectively; whereas the MIC against Gram-negatives were 0.42–0.67 mg/mL (w/v). 15 Another study has reported the antibacterial activity of R. coriaria against Gram-positives and Gram-negatives with MIC ranging from 0.5 to 20 mg/mL. 16
The potency of berry extracts against S. pyogenes ATCC 19615, the reference strain, could not properly be compared with previous studies since few have examined the anti-streptococcal effect of phytochemicals, particularly ethanolic and aqueous extracted crude extracts, against the strain. Conclusively, polyphenols, triterpenoids, and/or tannins have been tested and shown effectively against our strain of interest. Ethanolic extract of Diospyros lotus with high content of triterpenoids and tannins have MIC value of 5–10 mg/mL against S. pyogenes ATCC 19615. 17
Moreover, algae-derived phlorotannin was bactericidal against S. pyogenes ATCC 19615 at a concentration of 0.4 mg/mL. 18 Commercial phloretin, a dihydrochalcone, has also demonstrated bacteriostatic effect against the same strain of S. pyogenes at a concentration of 46.8 μg/mL. 19 Accordingly, phenolics were also effective against the same strain with MIC of ≤6.25 μg/mL and MBC of 100 μg/mL. 8 These observations are indicative of phytochemicals' prospective in the discovery and design of antibacterial compounds against S. pyogenes.
The killing rate of S. pyogenes ATCC 19615 by cranberry and sumac aqueous and ethanolic extracts was determined (Fig. 1). Cranberry aqueous extract at a concentration of 8 mg/mL (8 × MIC) following 24 h exposure lowered the bacterial population by ∼1.6 log CFU/mL below the initial inoculum (5.1 log CFU/mL), accounting for about 30% kill rate. At 24 h of incubation, the population in control doubled (∼9.8 log CFU/mL). A similar trend was recorded for cranberry ethanolic extract (Fig. 1). The sumac aqueous extract at 4 mg/mL drastically decreased the bacterial numbers to ∼0.9 log CFU/mL below the initial inoculum in a 10-min sampling time. At 24 h, viable cell numbers markedly dropped to below detection. Ethanolic extract of sumac at the highest concentration tested (2 mg/mL), at the 10-min sampling time, killed about 7% of the initial inoculum.
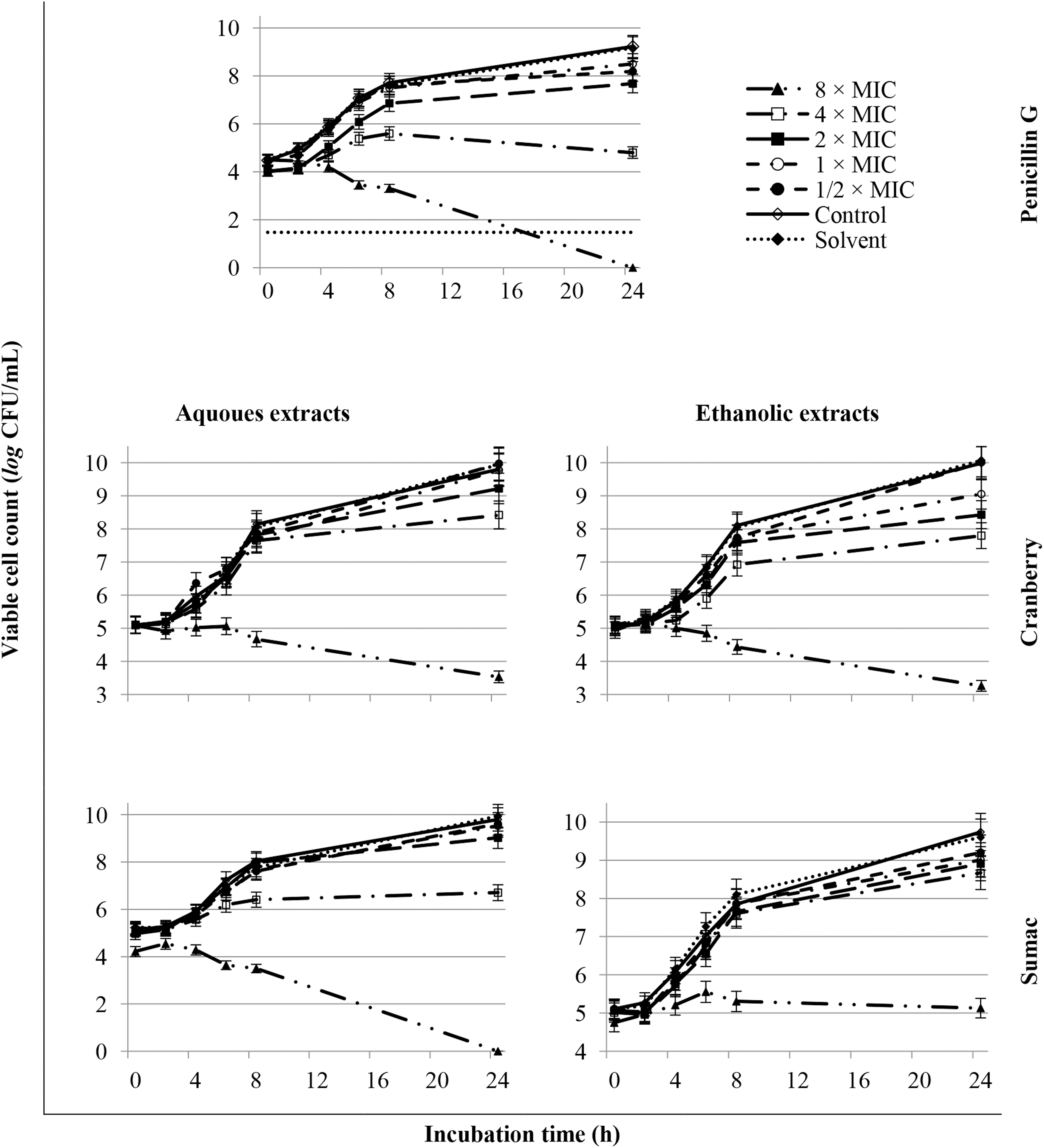
Time–kill kinetics for Streptococcus pyogenes ATCC 19615 in the presence of different extracts/compound. Wells without extracts/compound are considered control (positive control). Solvent (solvent control) means the highest concentration of distilled sterile water used to dissolve the extract (6.25% for cranberry aqueous and ethanolic extracts, 3.13% for sumac berry aqueous extract, 1.56% for sumac berry ethanolic extract, and 0.02% for penicillin G). The error bars represent the standard deviation of triplicate measurements. Limit of quantification is represented as dotted line. ATCC, American Type Culture Collection.
Penicillin at 16 ng/mL after 6 h exposure caused ∼3.6 log CFU/mL reduction compared with the positive control, and at 24 h period viable S. pyogenes ATCC 19615 bacterial numbers were lower than quantification limits. Similar to the present finding, the reported MIC of penicillin G for GAS varies from 0.003 to 0.12 μg/mL, depending on the test strain. 20
In vitro adherence inhibition activity of the selected extracts
In this static in vitro assay, anti-adhesive attributes of selected material were tested on the attachment of GAS. According to our observations, as presented in Table 2 and Figure 2, phytochemical-rich sumac and cranberry extracts had significant bioactivity against the attachment of our test bacteria to various coated and uncoated surfaces. In general, anti-adherence effects of sumac extracts were rather superior to cranberry extracts. In a dose-dependent manner, cranberry and sumac aqueous extracts were equally effective on the adherence of both strains to the bare surface (Table 2).
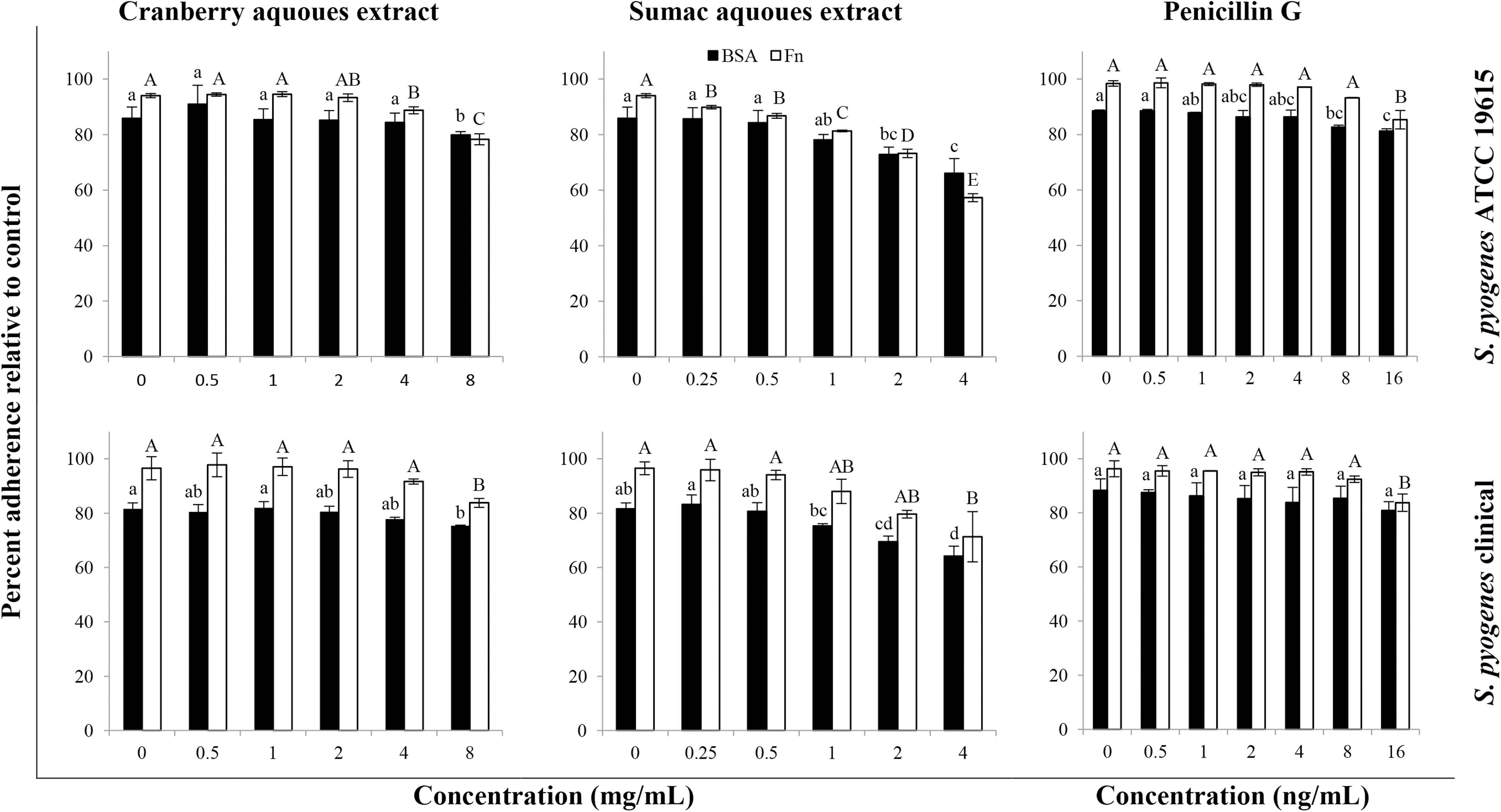
Inhibitory effects of cranberry and sumac aqueous extracts/compound on percent adherence of S. pyogenes to protein-coated surfaces after 1 h of treatment. Data are presented as percent adherence relative to the initial inoculum. ABCDColumns and abcdcolumns with different superscript letters on bars as obtained by Tukey's multiple comparison test are used to indicate statistical significance (P < .05) of different concentrations of compound, the solvent, and positive control. The error bars represent the standard deviation of at least two independent experiments. Black- and white-colored bars, respectively, represent anti-adhesion attributes of the test material on BSA and fibronectin-coated surfaces. BSA, bovine serum albumin; Fn, fibronectin.
Inhibitory Effects of Cranberry and Sumac Aqueous Extracts/Compound on the Adherence of Streptococcus pyogenes to Uncoated Surfaces After 1 h of Treatment
Each data point represents log CFU/mL of adherent bacterial cells after 1 h of treatment. Viable adherent cell number of solvent control and positive control was not significantly different.
Columns with different superscript letters as obtained by Tukey's multiple comparison test are used to indicate statistical significance (P < .05) of different concentrations of compound, the solvent, and positive control.
BQL, below lower limit of quantification; CFU, colony-forming units; NT, not tested.
The anti-adherence activity of sumac and cranberry aqueous extracts on the fibronectin-binding activity of strains was not constantly dose-dependent. Fibronectin-binding activity of the tested strains of S. pyogenes was equally sensitive to the inhibitory effects of extracts. Adhesion of GAS to BSA-coated substrate at 8–16 mg/mL of sumac and cranberry aqueous extracts was reduced by >2 to 3 log CFU/mL compared with the control. Nonetheless, effects varied from one to another type of fruit, type of extract, type of test strain, and/or type of substratum. Therefore, for this type of assay, many experimental factors shall be considered.
In general, GAS adhesion is mediated by capsule, pilus-like structures, and/or components of the cell wall, such as M protein, lipoteichoic acid, and fibronectin-binding proteins. 5 To date, more than 17 GAS adhesins have been identified. 3 GAS adhesion to the upper respiratory tract is initiated by receptor–adhesin interactions. 3,21 It is noteworthy that bacterial attachment to different substratum differs greatly among strains and is generally tissue-dependent. 22 Some GAS strains are capable of binding to abiotic polystyrene surfaces, whereas others need a matrix or a protein-coated surface for successful attachment and adhesion. 22,23
For instance, isolates from asymptomatic GAS carriers are most likely to carry the fibronectin-binding protein F2 gene, and these isolates tend to have higher adherence capabilities than isolates from pharyngitis subjects. 22 Our test strains showed similar adhesion capacity to both uncoated and BSA-coated substrates (Table 2 and Fig. 2). To the same extent, in equal experimental conditions, more of S. pyogenes attached to fibronectin-coated surface than to the uncoated surface (∼8–15%) (Table 2 and Fig. 2).
Generally, cranberry aqueous extract had a dose-dependent anti-adhesive effect on the attachment of ATCC 19615 to uncoated polystyrene surfaces. At the same concentration (8 mg/mL), cranberry aqueous extract did not show a significant effect on the adhesion of clinical strain of S. pyogenes to uncoated polystyrene surfaces, but at 16 mg/mL, significantly reduced the viable cell count (Table 2). At 8 mg/mL, cranberry aqueous extract reduced the adhesion of ATCC 19615 to polystyrene to ∼77% compared with the control (P < .05) (Table 2).
In the presence of 8 mg/mL of cranberry aqueous extract, the clinical and ATCC strains of S. pyogenes adhered 2% and 9% less, respectively, to the uncoated polystyrene surface than the control after 1 h period (Table 2), whereas the same concentration of extract did not affect the planktonic ATCC 19615 bacteria after 2 h of incubation (Fig. 1). This could be because cranberry contains considerable amounts of polyphenols, particularly flavonoids, which have been reported for their anti-streptococcal adhesive effects by a handful of studies, and the topic has been previously reviewed by the authors. 7 Polyphenol-rich tea extract at concentrations as low as 1–4 mg/mL prevented the attachment of Streptococcus mutans and Actinomyces viscosus to a significant degree. 24
In the present study, the anti-adhesive effect of cranberry aqueous extract was dose-dependent against the attachment of S. pyogenes ATCC 19615 to the bare substratum, but not the clinical strain (Table 2). Cranberry aqueous extract at 8 mg/mL had a higher adherence inhibition efficacy on fibronectin-binding proteins (13–16%) than on the nonspecific proteins (7–8% inhibition only) (Table 2 and Fig. 2). In a study, cranberry extracted high-molecular-weight nondialyzable material exhibited adhesion reduction activity in a dose-dependent manner at concentrations of 66–1330 μg/mL against the attachment of Streptococcus sobrinus to hydroxyapatite beads coated with fructosyltransferase and glucosyltransferase. 25
The sumac aqueous extract exhibited different anti-adherence effects than the cranberry aqueous extract. The sumac extract (4 mg/mL) reduced the adherence of S. pyogenes strains to the uncoated polystyrene surface by 23–32% after 1 h of incubation (Table 2), whereas the same concentration of extract killed ∼11% of the planktonic bacteria after 2 h of incubation (Fig. 1). The sumac aqueous extract at 4 mg/mL reduced the adhesion of S. pyogenes (clinical and ATCC 19615) to BSA by ∼20% compared with the control. However, the same concentration prevented more (up to 32%) of the adhesion of the bacteria to the uncoated substrate (Table 2).
The efficacy of sumac extract may be explained by its most abundant phytochemicals (caffeic acid, gallic acid, ellagic acid, and methyl gallate). 25 –27 For example, gallic acid and epigallocatechin gallate (ester of epigallocatechin and gallic acid) in green and oolong tea have been reported to be responsible for their inhibitory activities on the attachment of S. mutans and other oral bacteria to collagen, tooth surfaces, and gingival cell line. Similar to the cranberry aqueous extract, the sumac aqueous extract inhibited more fibronectin-binding adherence (25% and 32% for clinical and ATCC strains, respectively) than nonspecific adherence to BSA (19–20%). Sumac aqueous extract noticeably prevented more clinical isolate adhesion than that of the ATCC strain.
Penicillin had negligible effects on the adherence of S. pyogenes strains to uncoated polystyrene substrate (Table 2). The adhesion of S. pyogenes to the fibronectin-coated surface was more influenced by 16 ng/mL of penicillin (12–13% less attachment relative to the positive control) than the nonspecific proteins (∼8% attachment reduction) (Fig. 2). Penicillin G had a dose-independent effect on the fibronectin-binding proteins of S. pyogenes. It has previously been shown that inhibitory concentrations of penicillin could stimulate the loss of lipoteichoic acid, consequently reducing the adherence of Streptococcus sanguinis to host tissue surfaces. 28
Significant reduction in the adhesion of GAS strains regardless of their degree of virulence (noninvasive, highly invasive, and low invasive) to laminin were observed after overnight exposure of the bacteria to 1⁄2 × MIC of penicillin G. 22 Comparable to our study, noninvasive, highly invasive, and low invasive isolates of S. pyogenes adhered more to the laminin-coated plates than to the uncoated plates. 22 GAS adhesion to uncoated and coated substratum was decreased to about ≤60%, with treatments of 1⁄2 × MBC to MBC concentrations of the sumac and cranberry extracts. To better evaluate the anti-adherence effects of fruit extracts, we, in a cell study, examined the cranberry aqueous extract's attribute on the attachment of pretreated bacteria to the tonsil cells.
Cranberry aqueous extract at 2–4 mg/mL concentrations did not influence the adherence of S. pyogenes to the tonsil cells (Table 3). It is of note that higher concentrations of the material could not be tested due to its cytotoxic effects on the tonsil cells (Fig. 3). Lack of activity, however, may be related to the inappropriateness of the cell line as substrate in oral streptococci adhesion studies. For instance, buccal epithelial cells have been advised inappropriate for such studies due to the occurrence of nonspecific adhesion.
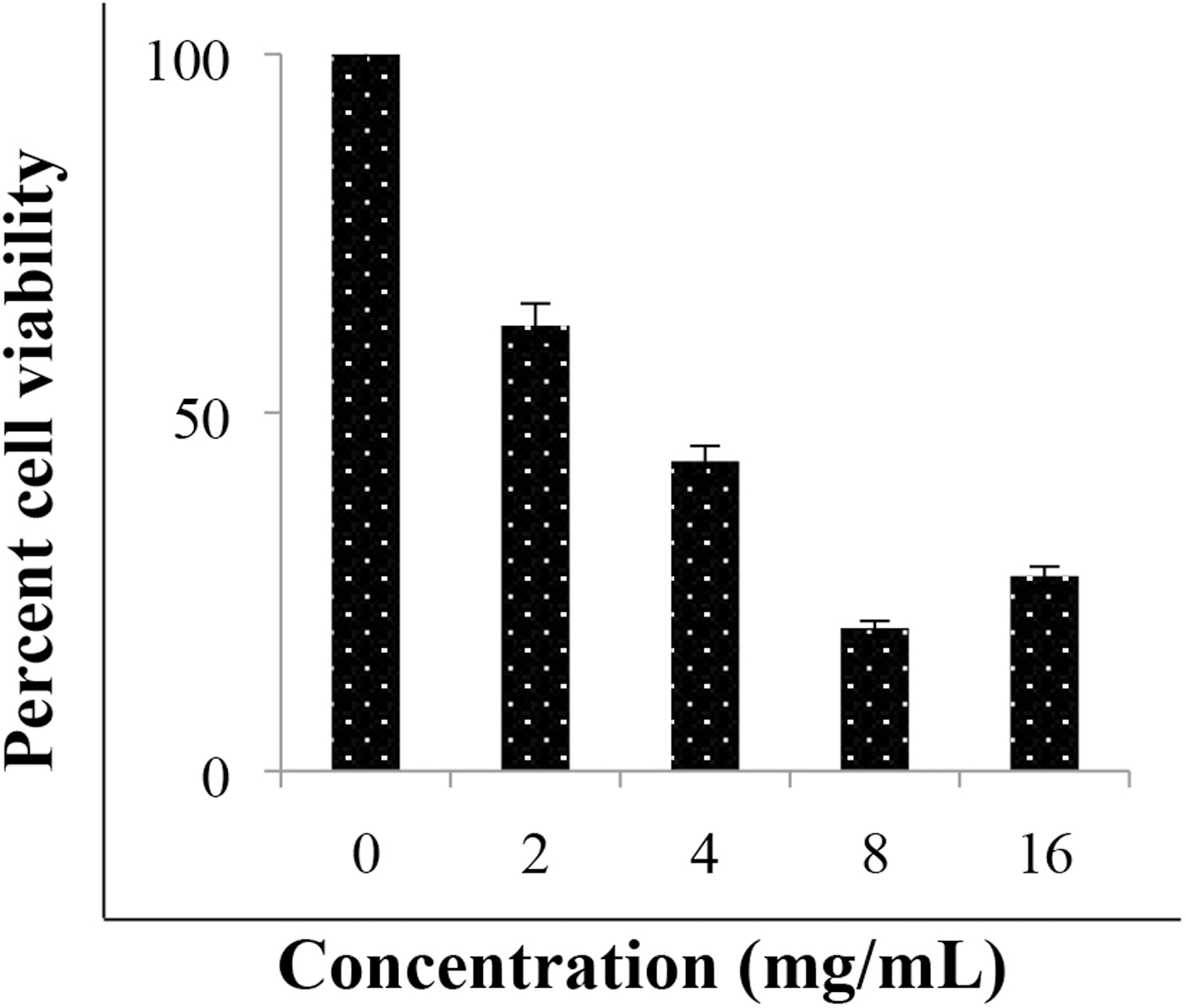
Percent viability of tonsil cells measured by MTT after 1 h of treatment with cranberry aqueous extract. The error bars represent the standard deviation of at least two independent experiments. MTT, 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide.
Inhibition Effect of Selected Compounds on the Adhesion of Streptococcus pyogenes to Tonsil Cells
Results are expressed as percent adherence of the bacteria to the HTEpiC cells relative to the control (control: 100%) ± standard deviation of triplicate measurements. Differences among solvent and positive controls were insignificant.
Columns with different superscript letters as obtained by Tukey's multiple comparison test are used to indicate statistical significance (P < .05) of different concentrations of compound, the solvent, and positive control.
Nonetheless, a handful of in vitro studies have documented the anti-adhesive effects of cranberry juice, its extract, and/or its constituents on the adhesion of bacteria to host cells. 29 Numerous studies have pointed out the explicit interactions between S. pyogenes and different niches of the host; however, to date, GAS adhesion mediated by specific molecules has not been completely understood. 3,4 This perhaps is due to the diversity of the adhesins expressed on the surface of GAS. 3
In a comparative study, the anti-adhesive activity of A-linked proanthocyanidins (PAC) from cranberry juice cocktail and B-linked PAC from commercial grape and apple juices, green tea, and dark chocolate were examined against the attachment of P-fimbriated uropathogenic Escherichia coli to uroepithelial cells. 29 Howell et al. showed the significant anti-adhesive effects of A-type PAC of cranberry at 60 μg/mL yet, at a higher concentration of 1200 μg/mL B-type PAC of grape juice could not produce any significant activity. 29 In agreement, there are a handful of reports on the higher anti-adhesive potency of cranberry A-linked dimers than its B-linked dimers.
Penicillin G at 16 ng/mL significantly reduced the adhesion of S. pyogenes, both ATCC and clinical strains, to the tonsil cells, which is in line with the studies of others. The sub-MIC of penicillin and rifampin alters the surface hydrophobicity of GAS, thus reducing the adhesion of bacteria to epithelial cells. Eickenberg et al. showed that penicillin G at a sub-MIC concentration (27 μg/mL) could reduce the adherence ability of E. coli to human buccal epithelial cells after 48 h of incubation (90% reduction). 30 In the same study, penicillin G at a sub-MIC concentration (1 μg/mL) had up to 70% anti-adhesive effects against the attachment of S. pyogenes to the same cells within the 3 h. 30
All taken together, no significant pattern of preferential attachment of both of the test strains to either fibronectin or tonsil cells was observed, suggesting that molecules implicated in the virulence of S. pyogenes are niche-specific and exceptionally distinctive adhesins are involved in the pathogenesis of the bacteria. Receptor-specific adhesins diverge greatly based on the streptococcal strain, host cell, and tissue. Furthermore, no final statement can be made on whether the number and location of A-type linkages in the oligomers, type of interflavan bonds, and/or degree of polymerization have any influence on the anti-adhesive activity of plant-derived bioactive material. 29 This may also be the case for the anti-adhesive activity of our cranberry extract.
Conclusion and perspectives
The findings of the current study suggest that crude extracts of fruits possess antibacterial effects against S. pyogenes. In the present study, both cranberry and sumac aqueous and ethanolic extracts dose- and time-dependently affected the growth of S. pyogenes ATCC 19615. Adherence of the bacteria was also significantly reduced by both the fruit extracts. It shall be noted that pretreating the bacteria with cranberry extract had an insignificant effect on the adhesion of S. pyogenes ATCC 19615 to the tonsil cells; however, the absence of activity may be due to the various confounding factors of the experiment rather than the sole bioactivity of the test material. Correspondingly, adhesion, the initial stage of infection, has to be the most important target for phytochemicals. Therefore, natural compounds with anti-adherence activities are ideal candidates for biofilm inhibition studies. In future studies, biofilm formation inhibition, as well as its eradication, could be examined.
Footnotes
Authors' Contributions
All the experiments were conducted by S.A. S.A. analyzed the data and wrote the draft article. All the co-authors contributed to designing the experiments and editing the article. H.P.V.R., the principal investigator, supervised the project.
Author Disclosure Statement
No competing financial interests exist. Animal and/or human subjects were not involved in this study.
Funding Information
We acknowledge the funds received from the Nova Scotia Graduate Entrance Scholarship, Graduate Research Training Initiative Scholarship, Natural Sciences and Engineering Research Council (NSERC) of Canada Collaborative Research and Development grant (CRDPJ 448052), and Island Abbey Foods Ltd., Charlottetown, PE, Canada. This article is based in part on research conducted in partial fulfillment of the requirements of S.A.'s Master of Science degree.