
Abstract
The objective of this study was to assay the ovicidal and larvicidal effects of two hydroalcoholic extracts (HA-E) and their aqueous fractions (Aq-F) and organic fractions (EtOAc-F) from Lippia graveolens (leaves) and Delonix regia (flowers) against the parasitic nematode Haemonchus contortus. The egg hatching inhibition test (% EHI) and mortality of infective larvae (% mortality) were used as biological models. The treatments of each plant (mg/mL, % EHI, and % mortality) were assigned as follows: HA-E (12.5–50, EHI; 25–200, mortality), Aq-F (6.25–25.00, EHI; 12.5–50, mortality), EtOAc-F (0.37–25, EHI; 3.12–50, mortality), negative controls (distilled water and methanol at 2% and 4%), and positive control (ivermectin at 5 mg/mL). Data were analyzed using a completely randomized design through an analysis of variance, considering the EHI and mortality percentages as the response variables. To identify the secondary metabolites, both plants were subjected to qualitative chemical analysis and HPLC. The results showed that both plants exhibited highly ovicidal and larvicidal effects. The bipartition in each HA extract allowed the increase of the anthelmintic effect in EtOAc-F, achieving a total ovicidal activity at 1.56 mg/mL and a larvicidal effect higher than 90% at 25 mg/mL. The qualitative chemical profile of both vegetal species identified alkaloids, flavonoids, condensed tannins, terpenes, and saponins. These results suggest that either L. graveolens leaves or D. regia flowers could be potential candidates for the control of parasitic gastrointestinal nematodes in small ruminants. However, in vivo studies with experimental animals to evaluate their possible toxicity and anthelmintic effect are necessary.
Introduction
Infections caused by gastrointestinal nematodes (GIN) in sheep are considered one of the main concerns in producers worldwide, since these parasites drastically affect the health and productivity of animals. 1 Haemonchus contortus is a GIN considered one of the most pathogenic parasites due to its hematophagous habit and its high prevalence in small ruminants. 2 The GIN are normally controlled using synthetic drugs and their indiscriminate use has triggered these parasites to develop resistance mechanisms to the different anthelmintic drug families. 3,4 For all these reasons, the need arises to search for other strategies of control, such as the selective deworming, 5,6 diets with nutritional quality, 7 copper particles, 8 biological control using nematophagous fungi, and using plants rich in secondary metabolites. 9,10 A number of reports from plant extracts under in vitro and in vivo conditions have shown important anthelmintic effects on the GIN of livestock. 11,12
Lippia graveolens H.B.K is an aromatic plant in the Verbenaceae family, commonly named oregano in Mexico, and it is used as a condiment in Mexican food. This plant has been used as herbal remedies that are purported to be analgesic, antipyretic, antidiarrheal and antimalarial. 13,14 Several parts of this plant have been reported to contain essential oils, iridoids, flavanones, and flavonoid glycosides. 15,16
Delonix regia Hook. Raf. is an arboreal legume named “tabachin” in Mexico, and it is a tree commonly found in some tropical and subtropical areas of Mexico. The uses of this legume in traditional medicine have been the treatment of rheumatism, bronchitis, diabetes, and malaria. 17 Many biological activities from extracts of this plant have been reported, such as anti-inflammatory, antioxidant, antimicrobial, antidiarrheal, and anthelmintic activities. 17 These bioactive compounds exist in leaves, flowers, seeds and bark, and include flavonoids, alkaloids, saponins, sterols, tannins, and phenolic acids. 18
Both plants are rich in secondary metabolites that have been identified with important anthelmintic effects. For instance, some compounds such as rutin, quercetin, tannins, hydroxycinnamic acid derivatives, and galloyl flavonoids are present in some plants identified as potential anthelmintics. 19 –22 Some of these constituents have been identified in L. graveolens and D. regia. The objective of this study was to evaluate the in vitro anthelmintic effect of the hydroalcoholic extracts (HA-E) and their fractions (aqueous and organic) from these two plants against eggs and infective larvae of the gastrointestinal parasite H. contortus.
Materials and Methods
Plant material
L. graveolens leaves were harvested from Iguala de la Independencia municipality (N 18°13′, W 99°29′, m.a.s.l), Guerrero, Mexico, in June 2018. D. regia flowers were collected from Yautepec municipality (N 18° 46′ y 18° 58′, W 98° 56′ y 99° 09′ m.a.s.l) State of Morelos, Mexico, in June 2018. One specimen of each plant was deposited in the Herbarium of the Centre for Research in Biodiversity and Conservation, Universidad Autónoma del Estado de Morelos (Voucher code numbers: to L. graveolens 39073 and to D. regia 39072). Both vegetal materials (5.0 kg) were dried at room temperature under dark conditions until a constant weight was obtained. After this process, the dry material in each plant was ground using an electrical miller (Willey mill, TS3375E15 model) to reduce the particle size to 3–6 mm.
Obtaining the HA-Es
The extracts in both plants were macerated with a hydroalcoholic solution (70% water and 30% methanol) using a ratio weight volume of 1:10 (1 g of sample to 10 mL of hydroalcoholic solution). Five hundred grams of each vegetal material was used, which was subjected to maceration process at room temperature (25–30°C) for 24 h. After this period, the liquid material was filtered using different filters (gauze, cotton, and filter paper) to obtain extracts free of material residues. A part of each liquid extract (750 mL) was totally concentrated by distillation under reduced pressure with the help of a rotary evaporator (Büchi R-300) to obtain an HA-E. For another part of the liquid material (3000 mL), only the methanol residues were eliminated by low-pressure distillation, and the solvent-free liquid extract was subjected to a liquid-liquid bipartition using ethyl acetate (1:1 v/v). Two fractions, an aqueous (Aq-F) and an organic fraction (EtOAc-F), were obtained. The solvents in both fractions were eliminated using low-pressure distillation. The dry extracts and fractions were stored at −80°C until later use in the in vitro bioassays.
Qualitative analysis of secondary compounds of the plant extracts
The chemical profile of the HA extracts, Aq-Fs and EtOAc-Fs, was carried out following several phytochemical testing procedures using different standard compounds and methods. Dragendorff, Mayer, and Wagner tests were used for alkaloids. 23 The Bornträger test was used to determine coumarin content, and the Mg2+ and HCl tests were used for flavonoids. 24,25 The ferric chloride test and the gelatin and saline solution tests were used for the confirmation of tannins. 26,27 The Liebermann-Burchard and Salkowski tests were used for triterpenes, and the foam formation test was used for saponins. 25
Major compound identification by HPLC
The HA-Es and their fractions (Aq-F and EtOAc-F) of each plant were subjected to a chromatographic analysis by HPLC using a Waters 2695 separation module HPLC system equipped with a Waters 996 photodiode array detector and Empower Pro software (Waters Corporation, USA). Chemical separation was achieved in a SUPELCOSIL LC-F column (4.6 × 250 mm, i.d., 5-μm particle size; Sigma-Aldrich, Bellefonte, USA). The phase mode consisted of 0.5% trifluoroacetic acid aqueous solution as solvent A and acetonitrile as solvent B. The gradient system was obtained as follows: 0–1 min, 0% B; 2–3 min, 5% B; 4–20 min, 30% B; 21–23 min, 50% B; 24–25 min, 80% B; 26–27 min, 100% B; and 28–30 min, 0% B. The flow rate was maintained at 0.9 mL/min and the injection volume was 10 μL. The absorbance was measured at 330 nm. The identification of the major compounds was established based on their UV spectra. 28,29
Biological material
H. contortus egg recovery procedure
To recover the H. contortus eggs, two sheep previously infected with the parasite (350 infective larvae/kg, INIFAP strain, Mexico) were used to act as egg-donor animals. The mean weight of the animals was 22.3 ± 0.4 kg of bodyweight. Animals were maintained indoors on a metabolic cage and they were supplied with hay and commercial concentrate food and water ad libitum. Egg recovery was performed according to the technique described by Coles et al. 30 Briefly, 35–50 g feces was weighed and macerated in a glass containing tap water (500 mL), and the fecal material was filtered through four sieves (400, 140, 74, and 37 μm), and finally, the eggs were cleaned by density gradient with 40% saccharose.
H. contortus infective larvae recovery procedure
Larvae were obtained from the donor sheep by daily collection (24 h). Fecal cultures were prepared by mixing feces with polystyrene particles in plastic bowls. Water was added to the fecal cultures and mixed with a wooden spoon to promote a better oxygenation and egg hatching. The fecal cultures were covered with foil and incubated for 7 days at room temperature (24–32°C). After this period, the infective larvae were extracted from fecal material using the Baermann technique. 31 The larvae were cleaned by density gradient (40% saccharose) and centrifugation, and exsheathed with sodium hypochlorite at 0.187%. Finally, the exsheathed larvae were used for the mortality assay.
Egg hatching inhibition test
The assay was carried out using 96-well microtitration plates (n = 12). One hundred H. contortus eggs in 50 μL distilled water were deposited in each well. This was followed by the addition of 50 μL extracts and fractions to obtain a final volume of 100 μL. Treatments in each plant were assigned as follows: HA-E (50, 25, and 12.5 mg/mL), Aq-F (25, 12.5, and 6.25 mg/mL), and EtOAc-F (25, 12.5, 6.25, and 3.12 mg/mL). For each treatment, two appropriate negative controls were included: 2% methanol and distilled water. Ivermectin (5 mg/mL) was used as a positive control. The plates were incubated at room temperature (18–25°C) for 48 h. After this period, the egg hatching process was stopped by adding 10 μL of Lugol's solution. Finally, the total eggs or larvae (L1 or L2) of each well were counted and the percentage of egg hatching inhibition (% EHI) was determined by the following formula: % EHI = [(number of eggs)/(number of larvae + number of eggs)] × 100.
Larval mortality assay
The assay was performed using 96-well microtitration plates (n = 12) for each treatment. Treatments were designed according to each plant as follows: L. graveolens. HA-E (100, 50, and 25 mg/mL), Aq-F (50, 25, and 12.5 mg/mL), and EtOAc-F (50, 25, 12.5, 6.25, and 3.12 mg/mL). In the case of D. regia, the assessed concentrations were assigned as follows: HA-E (200, 150, and 100 mg/mL), and Aq-F and EtOAc-F (50, 25, and 12.5 mg/mL). Each treatment was assessed using a negative control (distilled water and 4% methanol) and an anthelmintic (ivermectin, 5 mg/mL) as a positive control. Fifty microliters of an aqueous suspension containing 150 H. contortus larvae was deposited into each well. Then, 50-μL aliquots of extract and fractions, as well as controls, were individually deposited into each well. The plates were incubated at room temperature (18–25°C) for 48 h. Ten-microliter aliquots were taken from each well for dead or alive larvae counting.
The mortality percentage was estimated based on the criteria used by Olmedo-Juárez et al. 10 For this purpose, the following formula was used:
% Mortality = [(number of dead larvae)/(number of living larvae + number of dead larvae)] × 100.
Statistical analysis
The date of EHI and mortality percentages was normalized using a square root transformation and analyzed based on a completely randomized design using an analysis of variance in the general linear model in SAS. The difference among treatments was compared with the Tukey test (P < .05). Likewise, the lethal concentrations (LC50 and LC90) were estimated using a Probit analysis. 32
Results
Secondary compounds of the plant extracts and their fractions
The qualitative phytochemical profile of L. graveolens and D. regia is shown in Table 1. Both HA extracts of these plants contain alkaloids, flavonoids and tannins. L. graveolens extracts and their EtOAc-F displayed the presence of triterpenes. On the other hand, the HA extract and the Aq-F of this vegetal species were positive for saponins. With respect to D. regia, the HA extracts and their factions (Aq and EtOAc) were positive for alkaloids, flavonoids, and tannins. On the other hand, the HPLC chromatograms of both extracts and its fractions are shown in Figures 1 and 2. The retention time and UV spectra in L. graveolens showed the presence of flavonol (peak A), flavone (peaks B, F, G, J), coumarin (Peaks C, E), scopoletin (peak D), luteolin (peak H), and coumarate (peak I) (Fig. 1). Likewise, the major compounds found in D. regia flowers were coumarin (peaks A, C), quercetin glycoside (peak B), quercetin rhamnoside (peak D), coumarate (peaks E, G), and luteolin (peak F) (Fig. 2).

HPLC chromatograms corresponding to a HA-E, an Aq-Fn and an EtOAc-F from Lippia graveolens leaves. Showing the presence of phenolic compounds such as flavonol
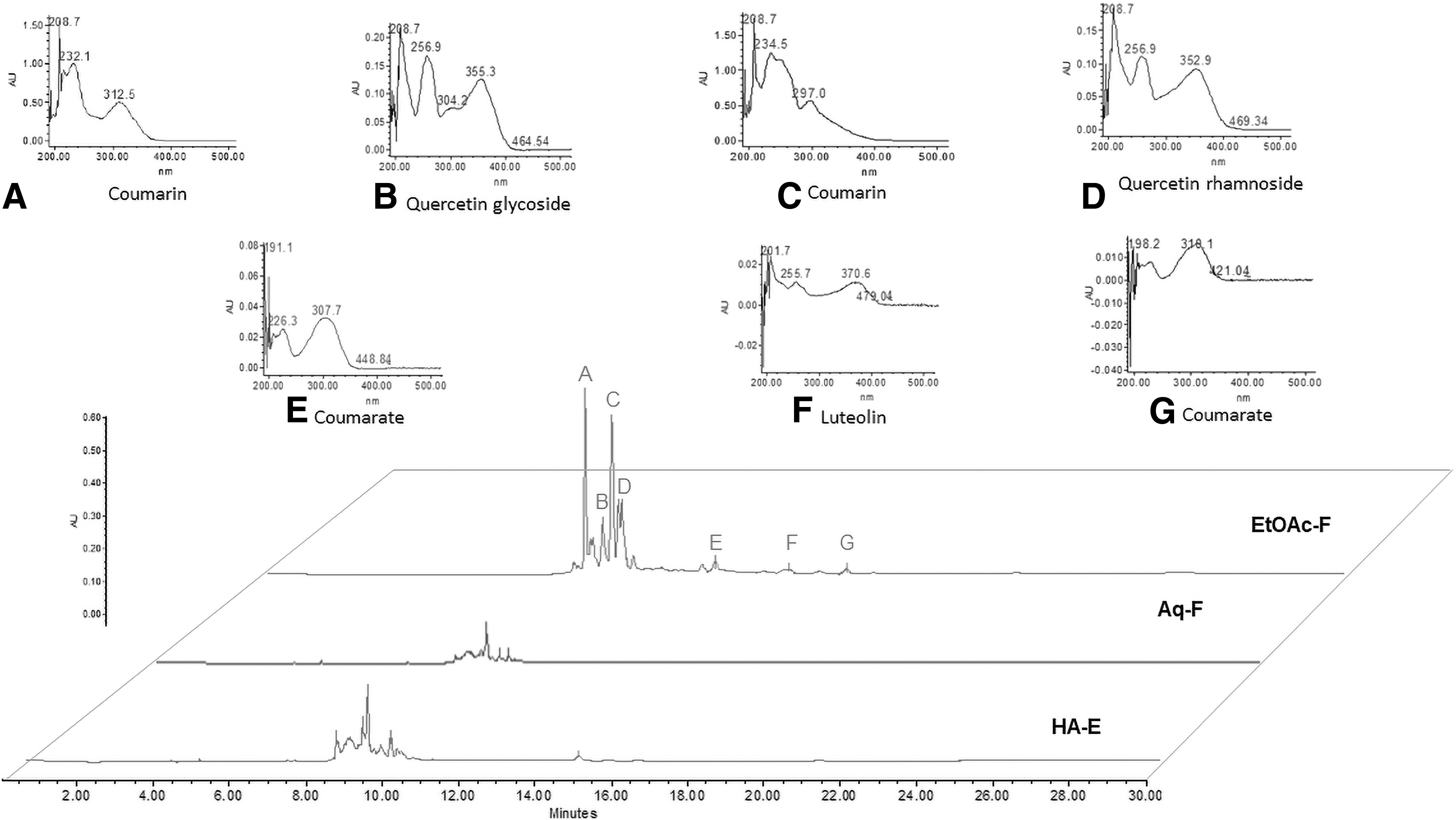
HPLC chromatograms corresponding to a HA-E, an Aq-F, and an EtOAc-F from Delonix regia flowers. Showing the presence of phenolic compounds such as coumarins
Phytochemical Screen of Lippia graveolens and Delonix regia
−, absence; +, presence; ++, positive reaction; +++, positive reaction strong; Aq-F, aqueous fraction; EtOAc-F, organic fraction; HA-E, hydroalcoholic extract.
EHI test
The results corresponding to EHI of H. contortus attributed to both HA extracts of L. graveolens and D. regia, and their aqueous fractions (Aq-F) and EtOAc-Fs are shown in Table 2. The L. graveolens HA-E displayed a total ovicidal effect for all evaluated concentrations. Meanwhile, the Aq-F showed no ovicidal activity for the tested concentrations (P > .05). On the other hand, the EtOAc-F had the highest inhibitory activity, causing a 100% EHI at 1.5 mg/mL.
Haemonchus contortus Egg-Hatching Inhibition After Exposure to Different Concentrations of Lippia graveolens and Delonix regia Hydroalcoholic Extracts and Their Fractions Expressed as Percentage
Means with different literal in the same column are statistically different (P < .05).
EHI, egg hatching inhibition; N/A, not active; SD, standard deviation.
With respect to the HA-E from D. regia, an egg-lethal activity ranging from 52.74% to 63.72% was observed in the tested concentrations. The Aq-F of this plant displayed a concentration-dependent effect (P < .05). Likewise, the EtOAc-F showed a total ovicidal effect at 1.5 mg/mL.
The lethal concentrations (LC50 and LC90) corresponding to EHI test of both plants are shown in Table 3. The LC50 of the EtOAc-F from L. graveolens was 0.45 mg/mL with confidence interval (CI) lower and upper limits of 0.49 and 0.52 mg/mL, respectively. The LC90 of this same fraction was 0.93 mg/mL (CI = 0.88–0.99 mg/mL).
Lethal Concentrations Required to Inhibit 50% and 90% of Haemonchus contortus Egg Hatching (LC50 and LC90) Following a 48-H Incubation Period with Lippia graveolens and Delonix regia
The bold values highlight the probability of lethal concentrations.
In regard to the LC50 and LC90 required for inhibiting the egg hatching processes of D. regia, the LC50 and LC90 for EtOAc-F were 0.34 and 0.89 mg/mL, respectively. On the other hand, the LC50 and LC90 for Aq-F of this plant were 11.83 and 128.55 mg/mL.
Mortality test of infective larvae (L3)
Table 4 shows the results of the mortality percentages of H. contortus L3 exposed to the HA extracts and fractions (Aq-F and EtOAc-F) of L. graveolens and D. regia as well as the negative controls (water and 4% methanol) and positive control (ivermectin at 5 mg/mL). The HA extract of L. graveolens had a concentration-dependent effect ranging from 35.38% to 78.94% mortality using 25–100 mg/mL. A similar effect was observed in the Aq-F of this plant with a 71.62% larvicidal efficacy at 50 mg/mL. The EtOAc-F of L. graveolens displayed the best larvicidal effect (P < .05) reaching 100% mortality with 50 mg/mL.
Haemonchus contortus Infective Larvae (L3) Mortality After Exposure to Different Concentrations of Lippia graveolens and Delonix regia Hydroalcoholic Extracts and Their Fractions Expressed as Percentage
Means with different literal in the same column are statistically different (P < .05).
—, not tested.
In regard to D. regia, the HA extract and the EtOAc-F had a larvicidal effect close to 100% at 100 mg/mL and the Aq-F showed a 86.82% mortality at 50 mg/mL.
Table 5 shows the mean lethal concentrations (LC50 and LC90) for both HA extracts L. graveolens and D. regia and their fractions required to cause mortality of H. contortus infective larvae. The HA extract of L. graveolens displayed an LC50 and LC90 of 39.41 and 175.21 mg/mL, respectively. The lethal concentrations recorded for the Aq-F of this plant were 14.84 and 244.79 mg/mL, respectively. Meanwhile, the EtOAc-F exhibited the best larvicidal effect, with an LC50 of 4.12 mg/mL and an LC90 of 9.60 mg/mL. On the other hand, the LC50 and LC90 values of D. regia HA extract were 17.20 and 83.54 mg/mL, respectively. The Aq-F had an LC50 of 29.82 mg/mL and an LC90 of 58.26 mg/mL. The EtOAc-F of this legume was more active, reaching LC50 and LC90 values of 2.50 and 18.13 mg/mL, respectively.
Lethal Concentrations Required to Cause 50% and 90% of Haemonchus contortus Infective Larvae Mortality (LC50 and LC90) Following a 48-H Incubation Period with Lippia graveolens and Delonix regia
The bold values highlight the probability of lethal concentrations.
Discussion
Results found in this study show evidence of the anthelmintic properties of both oregano leaves (L. graveolens) and Tabachin flowers (D. regia) on eggs and infective larvae of H. contortus. Gastrointestinal parasites affect all domestic species of animals. 3 The GIN are the main parasites that involve the health of the animals, and their control with synthetic drugs has resulted in these organisms developing mechanisms of resistance to different commercial anthelmintic families. 33,34 In this context, the search of other strategies for control of GNI is necessary and the plants rich in secondary metabolites could represent a viable way to control these parasites. A number of studies have shown that several plant families rich in secondary compounds exhibit anthelmintic effect against different gastrointestinal parasites. 10,21,35
L. graveolens is a plant with some pharmacological properties such as antibacterial, antioxidant, and antifungal activities. 36 –38 Likewise, it has been reported that the essential oils obtained from this plant exhibited anthelmintic effects against Giardia lamblia, a protozoan parasite of importance to public health. 39 In another study carried out by Ibarra-Moreno et al., 40 who evaluated extracts of different plants (among these L. graveolens) against the trematode Fasciola hepatica, it was found that the hexanic extract of L. graveolens displayed a 100% fasciolicidal activity. The evident anthelmintic activity of oregano in this study indicates that it could be a potential candidate plant as anthelmintic on H. contortus. The chemical separation of HA extract to obtain an Aq-F (Aq-F, polar compounds) and an EtOAc-F (EtOAc-F, low polar compounds) allowed us to obtain the best anthelmintic effect in the EtOAc-F (Tables 2 and 4). This marked effect was observed in the lethal concentrations (LC50 and LC90) of EtOAc-F in the EHI test, where the LC50 was 4.27 times more powerful than the HA extract. In contrast, in a study conducted by López-Aroche et al., 35 they evaluated an ethanolic extract from L. graveolens leaves and did not observe nematocidal effects against infective larvae of H. contortus. Therefore, the evident anthelmintic activity in our work could be due to an HA extract being used, which allowed us to obtain polar and nonpolar compounds, and this was confirmed when evaluating the EtOAc-F, resulting in a better biological activity.
D. regia is an arboreal legume that contains a number of phytochemical constituents, both in their leaves and flowers, for which it has attributed several medicinal properties, such as insecticidal, anti-inflammatory, antifungal, and anthelmintic activities. 17,41 –43 In this study, clear evidence was shown of the anthelmintic effect of flowers from this legume. The results obtained from the D. regia HA extract were similar to those observed in L. graveolens HA extract against H. contortus eggs. Meanwhile, in the mortality test, a larvicidal efficacy greater than 90% at 100 mg/mL was obtained (Table 4). The obtaining of Aq-Fs and EtOAc-Fs by a bipartition, allowed us to identify the main ovicidal (100% EHI) and larvicidal activity (greater to 90%) within the last fraction. When it was analyzed, the LC50 of the extracts and fractions (Aq and EtOAc) from D. regia against H. contortus eggs and infective larvae allowed us to determine that the EtOAc-F was 36.7 and 6.8 times more powerful than the HA extract (Tables 2 and 4). With respect to the LC90 against H. contortus eggs of this same plant, the EtOAc-F was 144.4 times more powerful than the Aq-F. Meanwhile, for the LC90 in the mortality test, the EtOAc-F was 4.6 and 3.2 times more active compared with the HA extract and the Aq-F, respectively.
There are reports in the literature on studies with several plant families that have assessed anthelmintic activities using different acetonic, methanolic, hexanic, and HA-Es. 35,44,45 The results found in this study with L. graveolens HA extract are similar to those of other studies. For example, Oliveira et al. 46 evaluated three HA-Es from Turnera ulmifolia, Parkia platycephala, and Dimorphandra gardneriana against H. contortus eggs. These authors reported an LC50 of 10 mg/mL in the extracts of T. ulmifolia (root and leaves) and D. gardneriana (leaves and wood). On the other hand, Camurça-Vasconcelos et al. 47 obtained essential oils from a plant of the Lippia genus (L. sidoides), which were orally administered (283 mg/kg of bodyweight) to sheep infected with a mixture strains of GIN and obtained an efficacy of 56.9% on the fecal egg reduction count from 14 days postadministration. According to these results, the nematocidal effect found in our study using L. graveolens could be the basis to carry out in vivo studies either with laboratory animals (gerbils) or with experimental sheep to corroborate the biological activity of this vegetal species.
In regard to the anthelmintic activity observed in D. regia, the ovicidal and larvicidal effects were more powerful than those that were reported in other works using plant extracts of the same Fabaceae family. For instance, Castillo-Mitre et al. 21 evaluated a HA-E from Acacia cochliacantha leaves and observed a 100% ovicidal effect on H. contortus eggs using a high concentration (100 mg/mL). Likewise, Olmedo-Juárez et al. 10 reported larvicidal effects close to 100% against H. contortus, Haemonchus Placei, and Cooperia punctate using a concentration of 200 mg/mL from A. cochliacantha HA leaves. In contrast, De Jesús-Martínez et al. 48 showed a total ovicidal effect only after using 0.78 mg/mL of a methanolic extract of Caesalpinia coriaria fruits, and the larvicidal effect close to 80% with a high concentration (100 mg/mL).
There are reports that L. graveolens leaves contain high contents of flavonoids, such as flavones (galagine, metilgalagine, apigenin, and quercetin), flavanones (naringenin, pinocembrin, and taxifolin), and chalcones (phloretine and phloridzine). 16 The D. regia flowers contain several polyphenolic constituents, such as tannins, gallic acid, quercetin, and kaempferol. 17 In recent studies, gallic acid has been isolated from fruits of C. coriaria and caffeoyl and coumaroyl derivatives from A. cochliacantha leaves and galloyl flavonoids from Acacia farnesiana pods. 20,43,44 These compounds were assessed on eggs of GIN of livestock and showed an efficacy close to 100% using lower concentrations (<10 mg/mL). In other works, evaluating plant extracts rich in tannins, the higher anthelmintic effects were found in those plants that contained gallic acid and galloyl derivatives. 18,49 According to the above-explained facts, for the anthelmintic effects in L. graveolens and D. regia, the anthelmintic activity could be related to these identified metabolites (Figs. 1 and 2). Therefore, this finding suggests that the isolation of the bioactive compounds in future studies is necessary to understand the biological activity.
The results of this research show that L. graveolens leaves and D. regia flowers contain ovicidal and larvicidal properties against the parasitic nematode H. contortus. The EtOAc-Fs from both plants were the most potent compared with the HA-Es and the Aq-Fs. These results justify continuing the investigation on both vegetal species in vivo under controlled conditions to verify the activities recorded in the in vitro bioassays.
Footnotes
Acknowledgments
This study formed part of the thesis work of Miss Anayely Bahena-Vicencio to obtain a degree in Biotechnology Engineering (at the Universidad Politécnica del Estado de Morelos, at Jiutepec, Morelos, Mexico) under the direction of Dr. Agustín Olmedo-Juárez.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
Part of this work was supported by INIFAP (Projects SIGI: 8215734475 and 1448123925).