
Abstract
The beneficial effects of sodium butyrate (NaB) and sodium propionate (NaP) on fatty acid oxidation (FAO) genes and production of proinflammatory cytokines related to nonalcoholic fatty liver disease (NAFLD) were evaluated using HepG2 human liver hepatocellular carcinoma cells exposed to palmitate/oleate or lipopolysaccharides (LPSs) as a model. The results showed that NaP or NaB was able to promote FAO, regulate lipolysis, and reduce reactive oxygen species production by significantly increasing the mRNA expression of peroxisome proliferator-activated receptor gamma coactivator 1 alpha (PGC-1α), peroxisome proliferator-activated receptor alpha (PPARα), adipose triglyceride lipase (ATGL), carnitine palmitoyltransferase 1 alpha (CPT1α), fibroblast growth factor 21 (FGF21), and uncoupling protein 2 (UCP2) in HepG2 cells. Together, NaP and NaB may produce greater effects by increasing CPT1α, PPARα, and UCP2 mRNA expression in LPS-treated HepG2 cells and by increasing CPT1α and ATGL mRNA expression in palmitate-/oleate-treated HepG2 cells. Only NaP treatment significantly increased FGF21 mRNA expression in palmitate-/oleate-treated HepG2 cells. The enzyme-linked immunosorbent assay results revealed that only pretreatment with LPSs and not palmitate/oleate significantly increased tumor necrosis factor alpha (TNF-α) expression in HepG2 cells. NaP alone or in combination with NaB significantly decreased TNF-α expression in LPS-induced HepG2 cells. The expression of interleukin-8 in both models showed no significant differences in all treatments. NaP and NaB show potential for in vivo studies on NAFLD.
INTRODUCTION
Nonalcoholic fatty liver disease (NAFLD) is a series of disturbances that involve various steps of liver damage including simple steatosis to nonalcoholic steatohepatitis, fibrosis, cirrhosis, or liver cancer. 1 NAFLD affects individuals who consume the typical Western diet consisting of high levels of refined sugars, fats, and other refined carbohydrates such as starch. Consequently, the increase of circulating free fatty acids as a result of weight gain and accumulation of visceral fat causes oxidative stress and liver inflammation.
Obesity, type 2 diabetes, hyperlipidemia, hypertension, and metabolic syndrome are major metabolic risk factors associated with NAFLD onset and progression. 2 NAFLD has a global prevalence of 20–30% in the general population and 80% prevalence in obese people. 2,3 In the United States, NAFLD has a 30% prevalence. 4,5
Gut dysbiosis and the translocation of lipopolysaccharides (LPSs) from the gut to the circulation affect liver homeostasis and significantly contribute to the development of NAFLD. The liver, in response to the elevated levels of dietary palmitic and oleic acid or LPS, activates the release of numerous proinflammatory cytokines, such as interleukin (IL)-8, IL-6, and tumor necrosis factor alpha (TNF-α), within the liver microenvironment. 6,7
Fatty acid oxidation (FAO) pathway is disrupted in NAFLD. 8 Peroxisome proliferator-activated receptor alpha (PPARα), a transcriptional activator of hepatic lipid metabolism genes such as carnitine palmitoyltransferase 1 alpha (CPT1α), controls the FAO pathway. 8 CPT1α is the rate-limiting enzyme of fatty acid beta (β) oxidation because it is involved in translocation of palmitic acid across the outer mitochondrial membrane into the mitochondrial matrix for oxidation. 9,10 In NAFLD patients, both PPARα and CPT1α levels significantly decrease in comparison with a healthy liver. 11
A key effect of the short-chain fatty acids (SCFAs) butyrate and propionate may inhibit histone deacetylases (HDAC), which are transcriptional modulators. Notably, in human HepG2 hepatocytes, butyrate upregulates PPARα levels by inhibition of HDAC3. 12,13 PPARα is a transcriptional activator that controls fatty acid metabolism and expression of fibroblast growth factor 21 (FGF21), an important metabolic hormone. 14 HDAC inhibitors possess antiproliferative effects, anti-inflammatory properties, and regulate cytokine expression. 15 –17
After absorption of butyrate, colonocytes use butyrate as their primary source of energy, and the remaining fraction goes through the hepatic portal vein to the liver. 18 Butyrate has beneficial effects that include anti-inflammatory properties, increases in mitochondrial activity, prevention of metabolic endotoxemia, increases in intestinal barrier function, and reduced body weight. 19 Butyrate also has low systemic toxicity, which makes it an excellent agent for clinical trials.
Propionate can provide energy for epithelial cells and is considered a substrate for hepatic gluconeogenesis. 20 The liver absorbs ∼90% of propionate, and the rest is sent into the peripheral systemic blood. 11 Propionate has beneficial anti-inflammatory properties, antihypertensive effects, and cardioprotective effects. 21 Both butyrate and propionate reduce food intake, weight gain, and high blood sugar. 22,23 Dysbiosis-induced inflammation can be cost effectively improved by using exogenous postbiotics such as byturate and propionate compared with treatment such as microbial transplantation.
We hypothesized that sodium butyrate (NaB), sodium propionate (NaP), or NaB/NaP combination will promote FAO and reduce proinflammatory cytokines in HepG2 cells. We evaluated the mRNA expression of genes (Table 1) associated with fatty acid metabolism and the protein expression of proinflammatory cytokines related to NAFLD using the HepG2 cells exposed to palmitate and oleate or LPS followed by treatment with NaB, NaP, or NaB/NaP combination.
Primer and Probe Sequences for Quantitative Reverse Transcription-Polymerase Chain Reaction
ATGL, adipose triglyceride lipase; CPT1α, carnitine palmitoyltransferase I alpha; FGF21, fibroblast growth factor 21; NCBI, National Center for Biotechnology Information; PGC-1α, peroxisome proliferator-activated receptor gamma coactivator I alpha; PPARα, peroxisome proliferator activated receptor alpha; UCP2, uncoupling protein 2.
RESULTS
HepG2 cells were exposed to LPSs to induce oxidative stress, and reductions in PPARα, peroxisome proliferator-activated receptor gamma coactivator 1 alpha (PGC-1α), CPT1a, and uncoupling protein 2 (UCP2) levels were observed

LPS-induced oxidative stress.
CPT1α expression was only induced by NaB or the NaB+NaP combination. UCP2, a stimulator of energy expenditure and fat oxidation in mouse liver, was upregulated at least twofold by NaB, NaP, or the combination. These results suggest that both NaB and NaP have beneficial effects on oxidative stress in human liver cells.
Next, we evaluated the effects of NaB and NaP on fat oxidation genes in a high-fat diet model using palmitate- and oleate-loaded HepG2 cells. Expression levels of PPARα and PPARα target genes adipose triglyceride lipase (ATGL), CPT1α, and FGF21 trended lower after fatty acid loading of cells (Fig. 2). Both NaB and NaP significantly upregulated PPARα and ATGL. Although Lee et al 25 showed that MS-275, a class 1 HDAC inhibitor, increased PGC1α mRNA and PPARα mRNA in palmitate-treated C2C12 myotubes, our study is the first to show that this mechanism is also pertinent to hepatocytes. However, only NaP and not NaB was a potent transcriptional activator of FGF21.

Effects of SCFA on gene expression in an HFD model.
The effects on FGF21, a circulating factor secreted by liver, indicate that NaP may have additional beneficial effects in other tissues such as adipose fat oxidation. Moreover, FGF21 reduces hepatic steatosis and has whole body anti-inflammatory effects. 26 NaB and the combination of NaB and NaP increased mRNA levels of CPT1α, but NaP had no effect even at the highest concentration used. These results show that NaB and NaP have different effects on gene expression in the high-fat model and suggest that the combination may have therapeutic benefits that are more potent than either SCFA alone.
LPSs significantly induced TNF-α (2112 pg/mL) compared with the control (1644 pg/mL) (Fig. 3a). Only NaP at 8 or 2 mM NaB/4 mM NaP combination significantly inhibited TNF-α secretion. NaB only at 2 or 4 mM significantly inhibited IL-8 secretion in the LPS model (Fig. 3b). Palmitate/oleate had no effects on TNF-α secretion compared with the control (Fig. 3c). The inhibition of IL-8 was not significant compared with the control in palmitate/oleate model (Fig. 3d). NaP, NaB, or in combination showed protective effects on palmitate-/oleate- or LPS-induced oxidative stress in HepG2 cells.
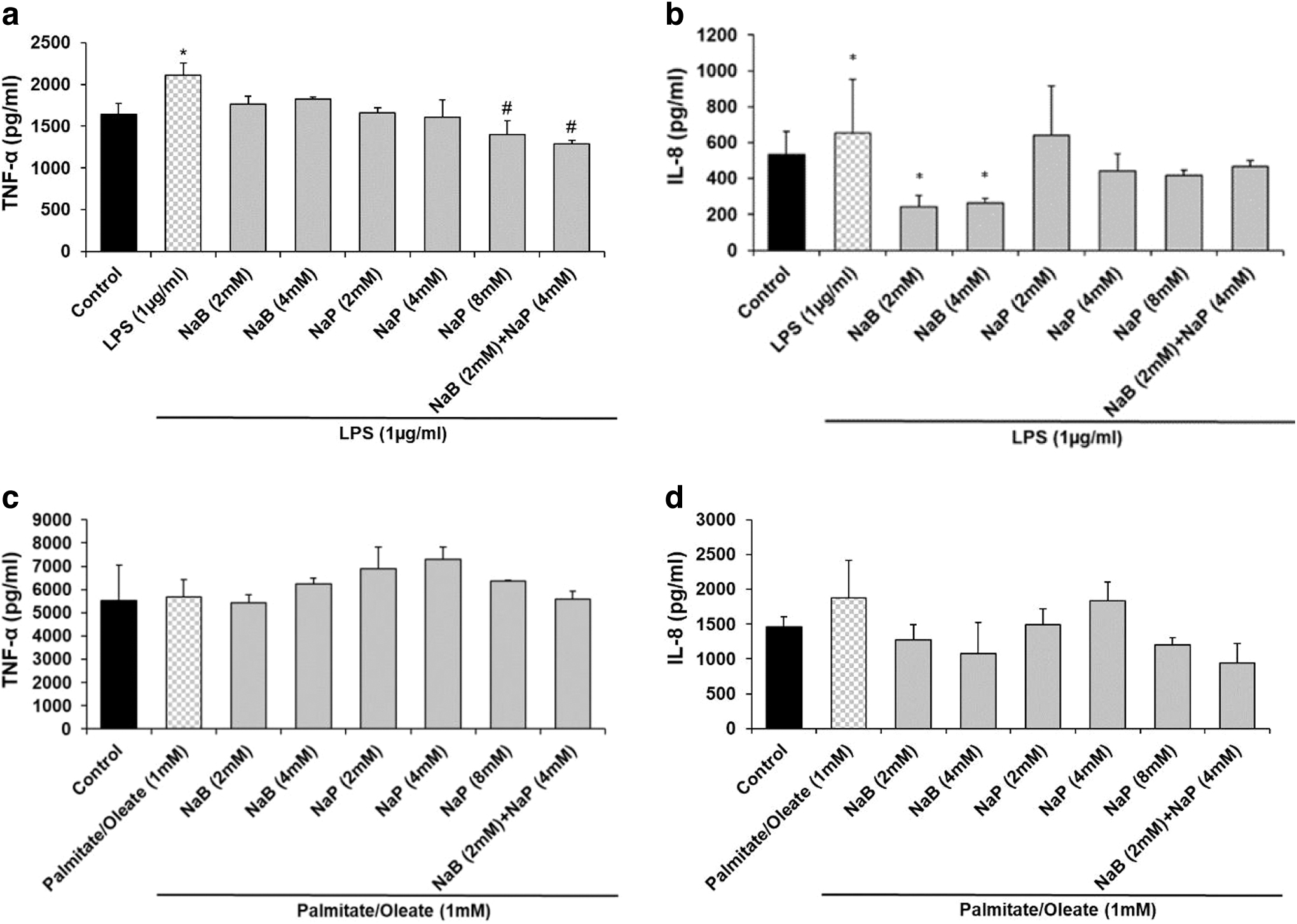
Effects of SCFA on inflammatory cytokines.
DISCUSSION AND CONCLUSION
NaP or NaB treatment promoted FAO, regulated lipolysis, and reduced ROS production through significant upregulation of PGC-1α, PPARα, ATGL, CPT1α, FGF21, and UCP2 mRNA expression in HepG2 cells. Together, NaP and NaB may produce synergistic effects by significantly increasing CPT1α, PPARα, and UCP2 mRNA expression in LPS-treated HepG2 cells and CPT1α and ATGL mRNA expression in palmitate-/oleate-treated HepG2 cells. NaP at 8 and 2 mM NaB/4 mM NaP combination increased the expression of CPT1α in the palmitate/oleate model.
Thus, NaB and NaP appear to reverse the effects of NAFLD, presumably through their ability to inhibit HDAC-3. NaB and NaP are both foods and increased delivery to the liver should be possible through dietary manipulations. Butyrate when given by itself is associated with a smell and taste that are associated with certain cheese and are offensive to some people. Increasing SCFA in the colon through dietary fiber fermentation would be a potential way to circumvent the taste problem.
Footnotes
AUTHOR DISCLOSURE STATEMENT
No competing financial interests exist.
FUNDING INFORMATION
This study was funded in part by LAB 94380 from the LSU AgCenter (J.N.L.). F.G. was Supported in part by 1 U54 GM104940 from the National Institute of General Medical Sciences of the National Institutes of Health, which funds the Louisiana Clinical and Translational Science Center. The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institutes of Health.