
Abstract
Chestnut inner shell (CIS) is rich in phenols and flavonoids such as gallic acid and ellagic acid, which are known to exhibit effective antioxidant and anti-obesity properties. Fermentation using lactic acid bacteria can enhance the physiological activity by increasing the contents of such functional ingredients. In this study, we evaluated the anti-obesity effects of a CIS extract subjected to a fermentation process (fermented CIS [FCIS]). Treatment with CIS and FCIS extracts (125, 250, and 500 μg/mL) increased cell viability and did not induce apoptosis, indicating no toxicity. The extract suppressed the gene expression of adipogenic factors, peroxisome proliferation-activated receptor gamma, CCAAT/enhancer binding protein (C/EBP) alpha, and C/EBP beta (by 7.75% and 67.59%, 21.41% and 66.27% in 500 μg/mL, respectively), and consequently suppressed the expression of downstream lipogenic factors such as fatty acid synthase, stearoyl CoA desaturase-1, citrate synthase, and ATP citrate lyase. The expression of factors involved in fat catabolism and β-oxidation increased in a dose-dependent manner, thereby preventing fat accumulation. This observation was consistent with the significant decrease in the staining intensity for lipid droplets, which indicated that lipid accumulation was decreased by 15.46% and 29.44% in 3T3L-1 and 27.01% and 46.68% in C3H10T1/2. Together, these results demonstrate the higher anti-obesity effects of FCIS extract than that of CIS extract, indicating the potential applicability of FCIS as an effective natural raw material to curb obesity.
Introduction
Most healthy people maintain their weight by maintaining a good balance between energy intake and consumption. 1 However, there has been an increase in the prevalence of obesity owing to higher energy intake than energy consumption as a result of dietary and environmental changes. 2 The World Health Organization (WHO) estimates that there are 340 million cases of obesity among 5 to 19-year-old individuals, and they are associated with metabolic syndromes such as type 2 diabetes, hepatic disease, and arteriosclerosis. 3 –5 Obesity is accompanied with a metabolic disorder caused by accumulation of the fat tissue in the body, not simply through an increase in body weight. 6 Fat accumulation is determined by the balance between fat production (lipogenesis) and fat breakdown (fatty acid oxidation).
Activation of transcription factors (TFs) involved with adipogenesis promotes fat accumulation. 7 The major adipogenic TFs, peroxisome proliferation-activated receptor gamma (PPAR-γ), and CCAAT/enhancer binding protein (C/EBP), are overexpressed in differentiating adipocytes. These TFs regulate the expression of lipogenic genes such as sterol regulatory element-binding protein-1c (SREBP-1c), fatty acid synthase (FAS), and stearoyl CoA desaturase-1 (SCD-1). 8 –11 In addition, fat accumulation is increased by citrate synthase (CS) and ATP citrate lyase (ACL), together responsible for transfer of citrate to mitochondria and hence promote fat production. 12 Conversely, fatty acid catabolism inhibits fat accumulation, and the first step of the fatty acid catabolism is β-oxidation that is a process in which fatty acids are decomposed in mitochondria to produce acetyl-CoA, reduced nicotinamide adenine dinucleotide, and reduced flavin adenine dinucleotide. 13 The enzymes involved in this process are acyl CoA dehydrogenase (ACD) and enoyl-CoA hydratase (ECH), 14 which produce ATP through fat breakdown and energy release, thereby inhibiting fat accumulation. Furthermore, activated mitogen protein kinase (AMPK), is known to promote β-oxidation. 15
AMPK interacts with sirtuin 1 (SIRT1), a lipid metabolism regulator in the liver and adipose tissue, to inhibit adipogenesis target molecules and induces expression of the nuclear hormone receptor PPAR-α. 16 Fat catabolism is activated through SIRT1/AMPK signaling, hence is critical in regulation of fat accumulation. Modulation of the SIRT1-AMPK axis using natural compounds was demonstrated elsewhere. For instance, resveratrol, quercetin, and flavone are known to inhibit fat accumulation through the regulation of SIRT1/AMPK signaling. 17 –19 Likewise, using purified constituents (e.g., pharmaceutical molecules), multiple attempts have been made to target this signaling pathway to regulate fat synthesis/degradation and control metabolic diseases and obesity. 20 However, purified compounds often present side effects such as sleep disorders, warranting further investigations to develop using natural materials with less side effects without compromising their efficacy. 21,22
Chestnut (Castanea crenata Sieb) is a deciduous forest tree belonging to the family Fagaceae. The fruit of the tree is edible and has been long used as medicinal and processed foods. In addition to the fruit, tree leaves, bark, chestnut peel, and chestnut inner shell (CIS) are also used as medicinal materials. 23 In particular, the CIS is rich in various phenols and flavonoids such as tannin, ellagic acid, and gallic acid, and its health promoting efficacies were examined in diverse models: antioxidant, anticancer, anti-aging, antidiabetes, and anti-obesity. 23 –27
Chestnut flesh has commonly been consumed with inner shell as a form of tea or traditional wine (e.g., rice wine; also known as makgeolli). Among processed products, chestnut makgeolli is manufactured through a fermentation process using lactic acid bacteria. Previously, it was demonstrated that the alcohol fermentation using the lactic acid bacteria improves not only extraction yield but also biological activities of natural products. 28 However, it is relatively unknown how fermentation of CIS will impact effects on its biological potential especially in the context of fat metabolism. In this study, therefore, we examined anti-obesity effects of fermented CIS (FCIS) extract, to explore its potential as a functional food manufacturing ingredient, and cytotoxicity was also assessed.
MATERIALS AND METHODS
Reagent
Dulbecco's modified Eagle's medium (DMEM), Roswell Park Memorial Institute 1640 (RPMI-1640) medium, and penicillin–streptomycin (PS) were purchased from Gibco (Gaithersburg, MD, USA). Fetal bovine serum (FBS) was obtained from GenDEPOT (Barker, TX, USA). Dexamethasone (DEX), 3-isobutyl-1-methylxanthine (IBMX), troglitazone (Tro), insulin, and dimethyl sulfoxide (DMSO) were purchased from Sigma-Aldrich (St. Louis, MO, USA).
Sample extract preparation
CIS was added to 50% ethanol and extracted using a fermenter (MJS U3, Marubishi, Tokyo, Japan) for 4 h at 80°C. The extracted CIS was filtered and concentrated, sterilized at 121°C for 30 min, and then 5% (v/v) Saccharomyces cerevisiae strain was inoculated and fermented for 24 h at 30°C. The FCIS with the S. cerevisiae was sterilized for 30 min at 121°C, and then filtered, concentrated, sterilized at 1 h, 100°C, and spray dried to be used in the experiment.
Cell culture and differentiation
RAW 264.7, 3T3-L1, and C3H10T1/2 cell lines were purchased from the Korean Cell Line Bank (Seoul, Korea). RAW 264.7 cells were cultured in DMEM supplemented with 10% FBS and 1% PS at 37°C in a 5% CO2 incubator. Murine 3T3-L1 preadipocytes were maintained in DMEM with 10% bovine serum albumin and 1% PS, whereas C3H10T1/2 cells were maintained in RPMI medium supplemented with 10% FBS and 1% PS in an incubator with 5% CO2 at 37°C. For differentiation, 3T3-L1 and C3H10T1/2 cells were plated in 12-well plates. After 5 days, the differentiation was induced using an induction media containing 0.5 mM IBMX, 1 μM DEX, and 1 μg/mL insulin); MDI supplemented with 1 μM Tro (day 0). On day 2 after the induction, cells were incubated with a media containing 1 μg/mL insulin and 1 μM Tro, and the medium was refreshed every other day. On day 8, CIS extract, FCIS extract, or 5% yeast were treated for 24 h to assess adipogenesis and lipogenesis effects in adipocytes (Fig. 1).

Time chart of the experimental procedure for 3T3-L1 and C3H10T1/2 cell differentiation. When 3T3-L1 and C3H10T1/2 cells that reached 70% confluency (day −5 to 0), the medium was changed to that containing MDI and Tro (day 0). Subsequently, medium was changed to insulin and Tro containing medium (days 2, 4, and 6). Cells were incubated with samples the day before harvesting. All experiments were then performed. MDI, reagent mixture of 0.5 mM IBMX, 1 μM dexametasone, and 1 μg/mL insulin; Tro, troglitazone.
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide assay
Cell viability was measured using the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay. RAW 264.7, 3T3-L1, and C3H10T1/2 cells were seeded in 96-well plates and allowed to settle for 2 h. Cells were treated with CIS and FCIS extracts at three different concentrations (125, 250, and 500 μg/mL), or 5% yeast for 24 h, followed by an incubation with 5 mg/mL MTT for 4 h. After 4 h of incubation, the medium was then removed, and the formazan crystals were dissolved in DMSO. The absorbance of the solution in each well was measured at 540 nm using a microplate spectrophotometer (SpectraMax M5; Molecular Devices, LLC, Sunnyvale, CA, USA).
ATP measurement
RAW 264.7, 3T3-L1, and C3H10T1/2 were seeded into 96-well plates, respectively, followed by stabilization for 2 h. Subsequently, cells were treated with CIS, FCIS, or 5% yeast for 24 h in a 5% CO2 incubator. Afterward, the cytotoxicity of the extracts was calculated based on the degree of cell ATP luminescence using Cell Titer-Glo Luminescent Cell Viability Assay Reagent with the GloMax Discover Microplate reader (Promega, Madison, WI, USA).
Flow cytometric analysis for cell proliferation
To investigate the effects of CIS, FCIS, and 5% yeast on the proliferation of RAW 264.7 cells, a BD FACS Calibur Flow Cytometer (Becton, Dickinson and Company, Franklin Lakes, NJ, USA) was used. RAW 264.7 cells in 24-well plates were treated with CIS and FCIS extracts and 5% yeast for 24 h. Cells were double-stained with 7-aminoactinomycin D/carboxy fluorescein succinimidyl ester (CFSE) solutions. After staining, cell proliferation was monitored using flow cytometry.
Lipid droplet staining
After 6 days of adipocyte differentiation, 3T3-L1 and C3H10T1/2 cells were treated with CIS and FCIS extracts and 5% yeast for 24 h and then washed with phosphate-buffered saline (PBS) and fixed with 4% formaldehyde for 1 h. After fixation, the cells were washed twice with 2-propanol and stained with Oil Red O (Sigma-Aldrich) reagent or Nile red (Invitrogen, Waltham, MA, USA) for 1 h. The stained cells were observed a light (Leica Microsystems, Wetzlar, Germany) or fluorescence microscope (Carl-Zeiss, Oberkochen, Germany). Thereafter, the Oil Red O reagent was dissolved with 2-isopropanol and the absorbance was measured at 520 nm.
RNA isolation and real-time reverse-transcriptase polymerase chain reaction
Cells treated with CIS and FCIS extracts (125, 250, and 500 μg/mL) and 5% yeast were washed with PBS and the RNA was isolated using the RNeasy Plus Mini Kit (Qiagen, Valencia, CA, USA). The cDNA was synthesized using M-MLV Reverse Transcriptase kit (Promega) according to the manufacturer's protocol. The synthesized cDNA was subjected to reverse-transcriptase polymerase chain reaction (PCR) using the Amfisure qGreen Q-PCR Master Mix (GenDEPOT) and Qiagen rotor gene Q real-time PCR machine. The target gene was amplified by denaturation at 95°C for 15 sec, annealing at 57°C for 20 sec and extension at 72°C for 30 sec. The relative quantification of mRNA level was performed following normalization to glyceraldehyde 3-phosphate dehydrogenase (GAPDH) level. The following mouse specific primers were used: C/EBPβ, 5′-GAG CCA AAA GGG TCA TCA TC-3′ (forward), 5′-TAA GCA GTT GGT GGT GCA GG-3′ (reverse); PPAR-γ, 5′-GAG CCA AAA GGG TCA TCA TC-3′ (forward), 5′-TAA GCA GTT GGT GGT GCA GG-3′ (reverse); C/EBPα, 5′-GAG CCA AAA GGG TCA TCA TC-3′ (forward), 5′-TAA GCA GTT GGT GGT GCA GG-3′ (reverse); SREBP-1c, 5′-GAG CCA AAA GGG TCA TCA TC-3′ (forward), 5′-TAA GCA GTT GGT GGT GCA GG-3′ (reverse); FAS, 5′-GAG CCA AAA GGG TCA TCA TC-3′ (forward), 5′-TAA GCA GTT GGT GGT GCA GG-3′ (reverse); SCD-1, 5′-GTT CGT TAG CAC CTT CTT GCG-3′ (reverse); CS, 5′-GGC ACC CAA CAT TTG AGT TAT T-3′ (forward), 5′-CTT GAG GCA CAG CAG GTA TAG-3′ (reverse); ACL, 5′-AGA GAA GGT TGC CCA AGT ATT C-3′ (forward), 5′-CGA GCA CAG ATG ATG GTG TTA-3′ (reverse); ECH, 5′-GTG GAG CAA ATG ACA ACT TCT G-3′ (forward), 5′-GGC TTC TGG TAT CGC TGT ATT-3′ (reverse); ACD, 5′-CAG CCA ATG ATG TGT GCT TAC-3′ (forward), 5′-CAT ACT CGT CAC CCT TCT TCT C-3′ (reverse); SIRT1, 5′-GGA ACC TTT GCC TCA TCT ACA-3′ (forward), 5′-CAC CTA GCC TAT GAC ACA ACT C-3′ (reverse); AMPK, 5′-CGA CCT GGA AGC GAA TGA TAA-3′ (forward), 5′-GTT GAA GGA CCC AGA CAA GTA G-3′ (reverse); PPAR-α, 5′-GGT TCC TGG TGC CGA TTT AT-3′ (forward), 5′-CAC AGA CTA GCA TCC CAC TTA AT-3′ (reverse); GAPDH, 5′-GAG CCA AAA GGG TCA TCA TC-3′ (forward), 5′-TAA GCA GTT GGT GGT GCA GG-3′ (reverse).
Western blot analysis
3T3-L1 and C3H10T1/2 cells were seeded in 12-well plates, differentiated, and then treated with CIS extract, FCIS extract, and 5% yeast. After incubation, the cells were harvested in a lysis buffer on ice and centrifuged at 12,000 g for 10 min at 4°C to obtain protein. Protein quantification was performed using the BCA protein assay reagent (Gen-DEPOT). In total, 10 μg of protein was separated by 4–20% sodium dodecyl sulfate–polyacrylamide gel electrophoresis and the protein bands were transferred onto polyvinylidene difluoride membranes. The membranes were blocked with skimmed milk (50 mg/mL) for 1 h and then incubated with primary antibodies against C/EBPβ, PPAR-γ, C/EBPα, SREBP-1c, FAS, SCD-1, CS, ACL, ECH, ACD, AMPKα, PPAR-α, and β-actin for overnight at 4°C. The bands were visualized using an enhanced chemiluminescence reagent (Pierce, Rockford, IL, USA) and captured with a ChemiDoc image detector (Bio-Rad, Hercules, CA, USA).
Statistical analysis
Data were expressed as the mean ± standard error of the mean. Experimental group comparisons were performed using the one-way analysis of variance. When significance (P < .05) was found, the differences in mean values were determined with the Duncan's multiple range tests. The statistical analyses were performed using SPSS (version 25.0; SPSS, Inc., Chicago, IL, USA).
RESULTS
Effect of FCIS extracts on cell viability
To determine the noncytotoxic doses of FCIS extract for the antiadipogenic study, we performed cell viability analysis using two methods (MTT and ATP) and investigated the cytotoxic potential of CIS extract, FCIS extract, and 5% yeast in three cell lines. As given in Figure 2, the viability of all three cell lines increased after CIS and FCIS extract treatment in a dose-dependent manner. In particular, FCIS extract was not toxic to cells and the cell proliferation was higher after FCIS extract treatment than that observed after treatment with CIS extract. Therefore, CIS extract, FCIS extract at 125, 250, and 500 μg/mL concentrations, and 5% yeast were selected to evaluate the anti-obesity effects.
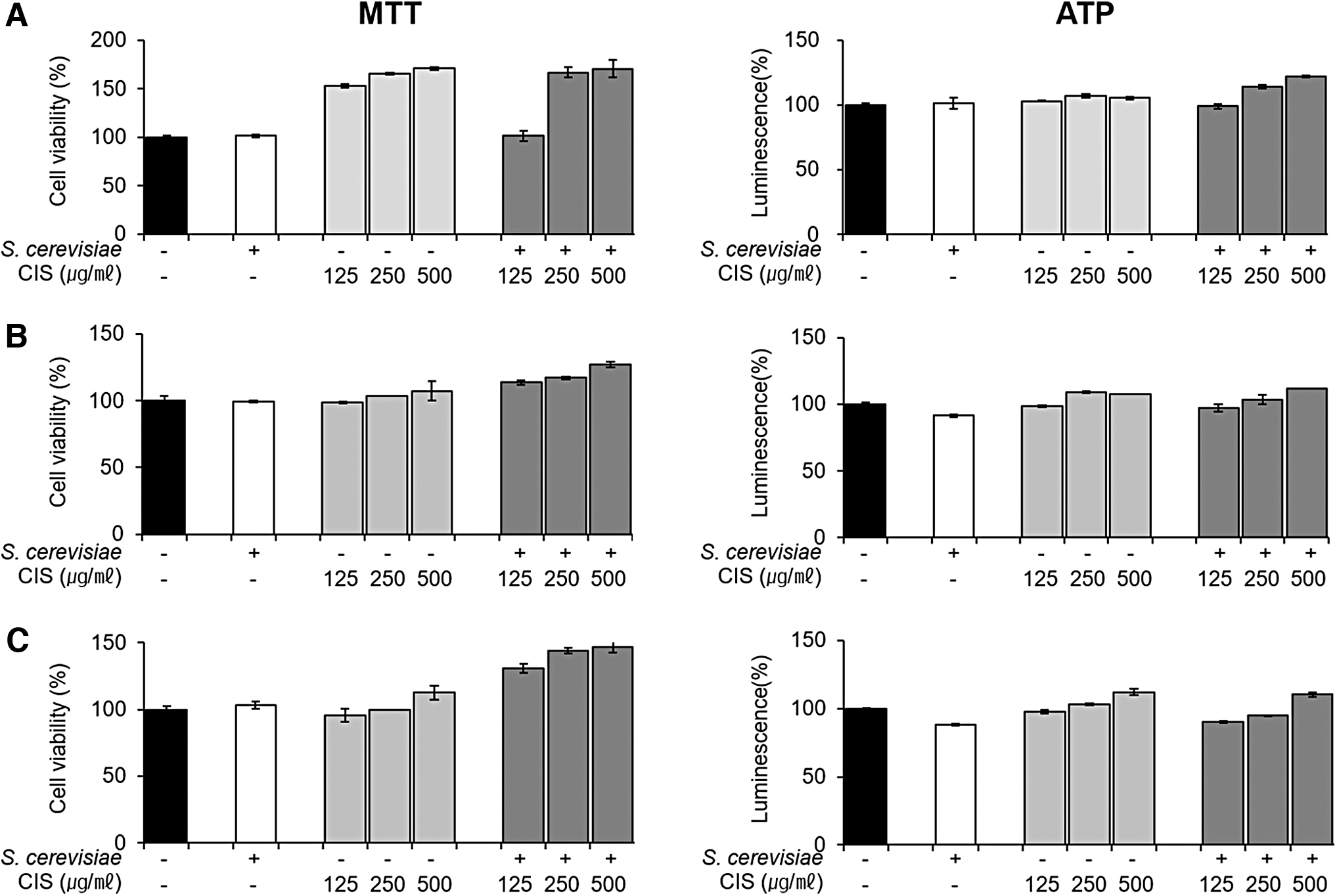
Effect of CIS extract fermented by 5% yeast (Saccharomyces cerevisiae) on the viability of three cell types. The viability of
Effect of FCIS extract on cell proliferation
The proliferation of RAW 264.7 cells treated with CIS extract, FCIS extract, and 5% yeast was measured by flow cytometry analysis following staining with CFSE. As given in Figure 3, the treatment with CIS and FCIS extracts increased RAW 264.7 cell proliferation as compared with the control treatment. However, 5% yeast had minor effects on the proliferation of RAW264.7 cells. In addition, the cell proliferation increased after treatment with the extracts in a dose-dependent manner. The cell proliferation was higher with FCIS than with CIS. Overall, these observations suggest that the extracts did not exert any toxicity to cells, but instead promoted their proliferation.
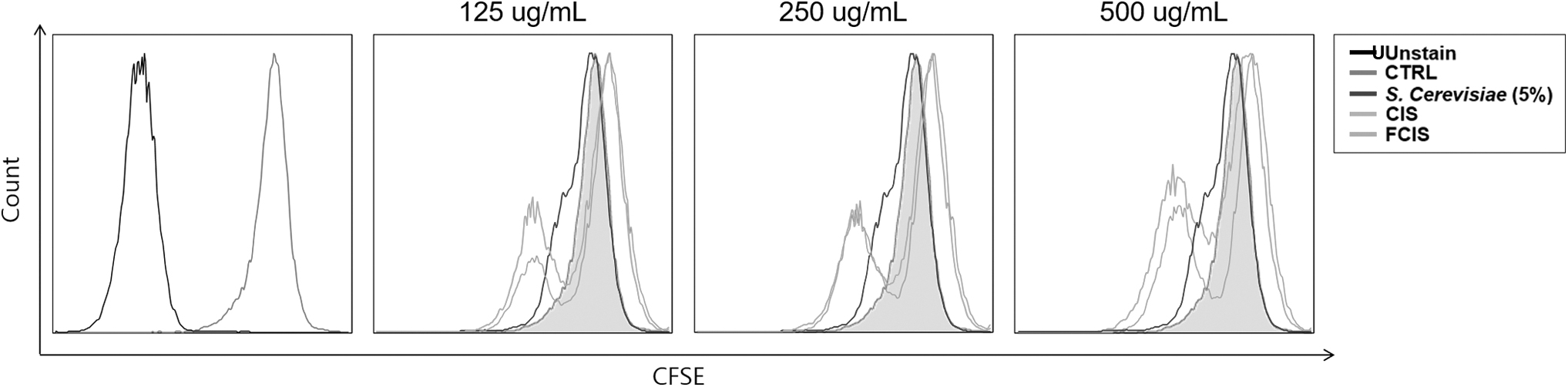
Effect of CIS extract fermented by 5% yeast (S. cerevisiae) on the proliferation of RAW 264.7 cells. After pretreatment with extracts or yeast for 24 h, the proliferation of RAW264.7 cells was analyzed by flow cytometry following staining with CFSE. CFSE, carboxy fluorescein succinimidyl ester; CTRL, control; FCIS, fermented chestnut inner shell.
Effect of FCIS extracts on lipid accumulation in adipocytes
Oil Red O (Fig. 4A, B) and Nile red (Fig. 5A, B) staining were performed to evaluate the inhibition of lipid accumulation following treatment of differentiated 3T3-L1 and C3H10T1/2 cells with FCIS extract. Lipid accumulation was observed under a microscope (light or fluorescence) by lipid droplet staining. At a high concentration of 500 μg/mL, FCIS extract remarkably inhibited lipid droplet accumulation. Furthermore, FCIS extract reduced lipid droplets more effectively than CIS extract. Thus, the anti-obesity effects of CIS extract seemed to have improved after fermentation as compared with the effect of 5% yeast. As given in Figure 4C and D, lipid accumulation in adipocytes increased by 61% as compared with that in preadipocytes, and lipid levels decreased by 15.46% and 29.44% in 3T3L-1, 27.01% and 46.68% in C3H10T1/2 at the highest CIS and FCIS extract concentrations, respectively.

Effect of CIS extract fermented by 5% yeast (S. cerevisiae) on lipid accumulation in differentiated adipocytes. Differentiated
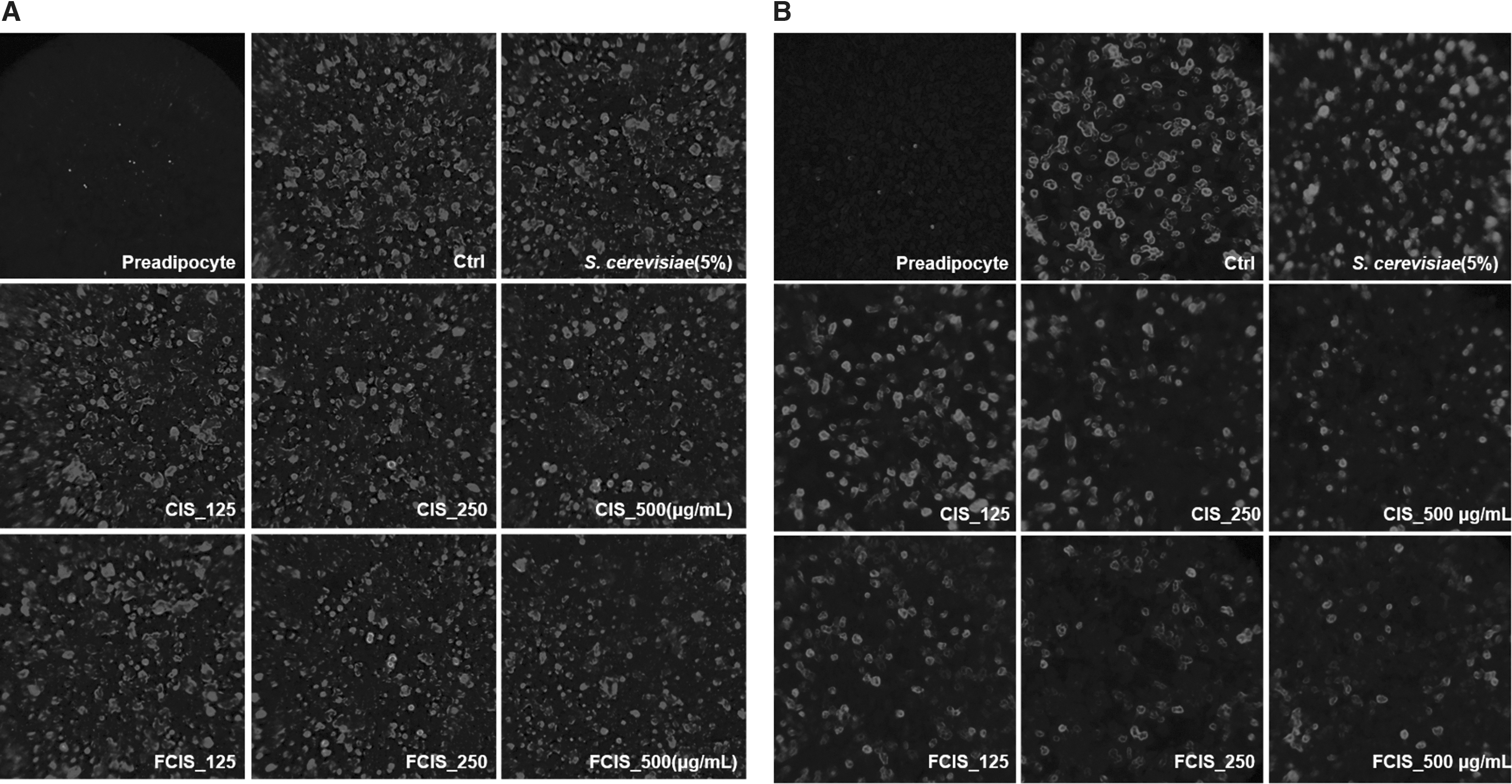
Effect of CIS extract fermented by 5% yeast (S. cerevisiae) on lipid accumulation in differentiated adipocytes. After treatment of differentiated
Effect of FCIS extracts on lipogenesis and adipogenesis expression in adipocytes
Real-time PCR was performed to confirm the expression of adipogenesis-related genes in differentiated adipocytes (Fig. 6). The expression of lipogenesis-related genes SREBP-1c, FAS, and SCD-1 was significantly downregulated after CIS and FCIS extract treatment (Fig. 6A). The adipogenesis TFs C/EBPβ, PPAR-γ, and C/EBPα were expressed during differentiation, and their expression was regulated by CIS and FCIS extracts to control the levels of lipogenesis-related factors. The mRNA expression levels of CS and ACL, which synthesize fats by stimulating citric acid production, also significantly decreased after FCIS treatment. The effect was similar at all tested concentrations. This effect was more pronounced with FCIS extract than with CIS extract. Adipogenesis is promoted by the expression of downstream factors through an increase in the expression of C/EBPα and PPAR-γ by C/EBPβ factors. The treatment of differentiated adipocytes with FCIS extract led to a decrease in the protein expression of adipogenic TFs in a dose-dependent manner. Thus, the levels of lipogenic factors called as downstream factors of lipogenesis such as FAS, SREBP-1c, and SCD-1 also decreased. In addition, the protein expression of CS and ACL also reduced after FCIS extract treatment (Fig. 6B).
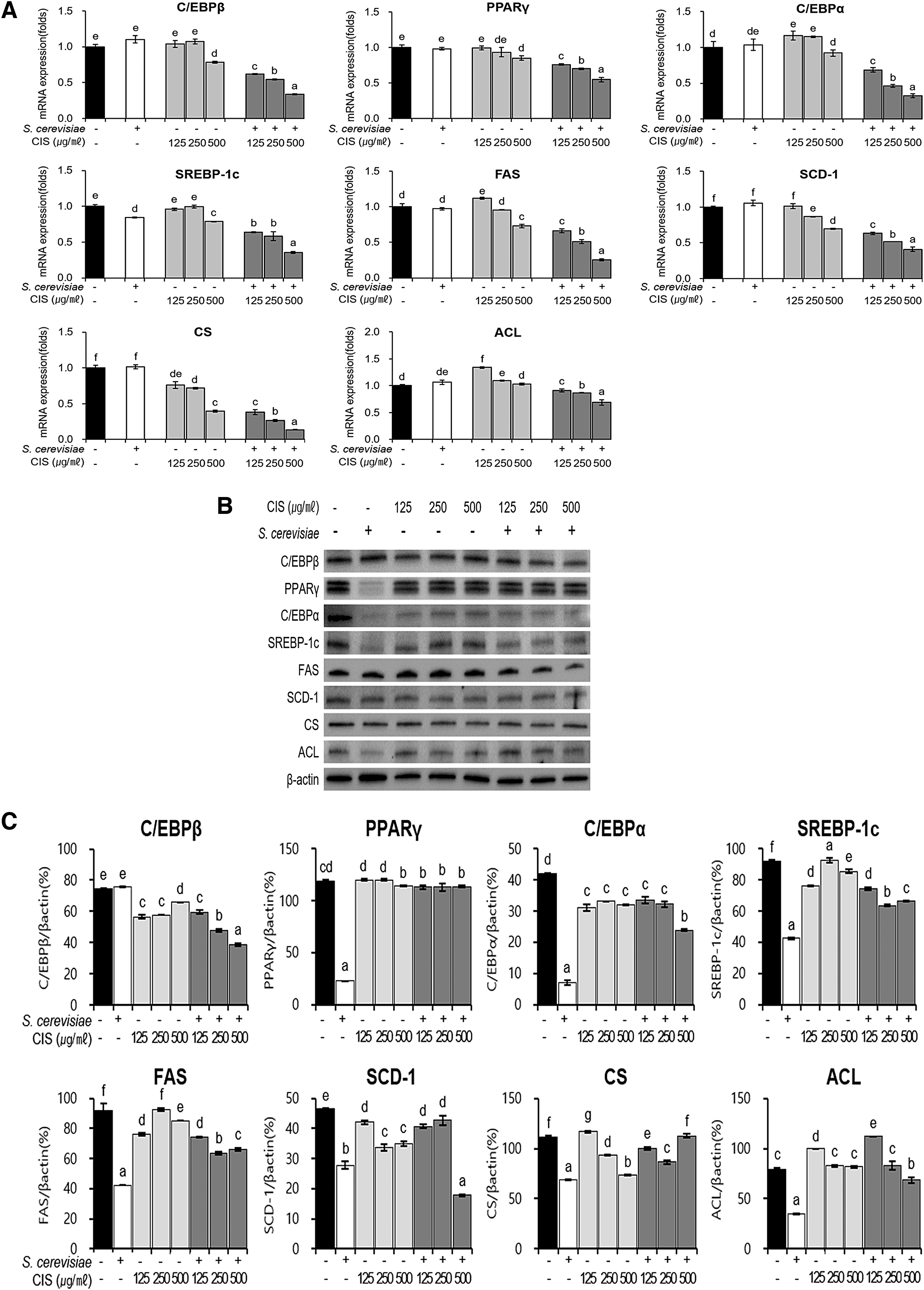
Effects of CIS extract fermented by 5% yeast (S. cerevisiae) on the expression of the genes associated with adipogenesis and lipogenesis in C3H10T1/2 cells. After treatment of differentiated C3H10T1/2 cells with extracts or yeast for 24 h,
Effect of FCIS extract on the genes related to fatty acid catabolism in adipocytes
We evaluated the expression of the factors involved in β-oxidation, such as ACD, ECH, SIRT1, and AMPK, that inhibit fat accumulation by promoting fatty acid catabolism. The treatment of differentiated adipocyte cells with CIS and FCIS extracts led to an increase in the expression of ACD and ECH in a dose-dependent manner. In addition, the mRNA expressions of SIRT1, AMPK, and PPAR-α, which promote β-oxidation, also increased, which is thought to decrease lipid accumulation (Fig. 7A). In comparison with CIS extract, FCIS extract increased the protein expression level of the β-oxidation factor ECH, which boosts fat catabolism and decreases the protein expression of total AMPKα, the major biomarker of the SIRT1/AMPK pathway (Fig. 7B). Therefore, it was confirmed that FCIS was more effective in mediating the anti-obesity effects and suppressing fat accumulation than CIS extract, owing to the fermentation process. In particular, FCIS had better effects on activating fatty acid catabolism than CIS extract, thereby enhancing the anti-obesity function.
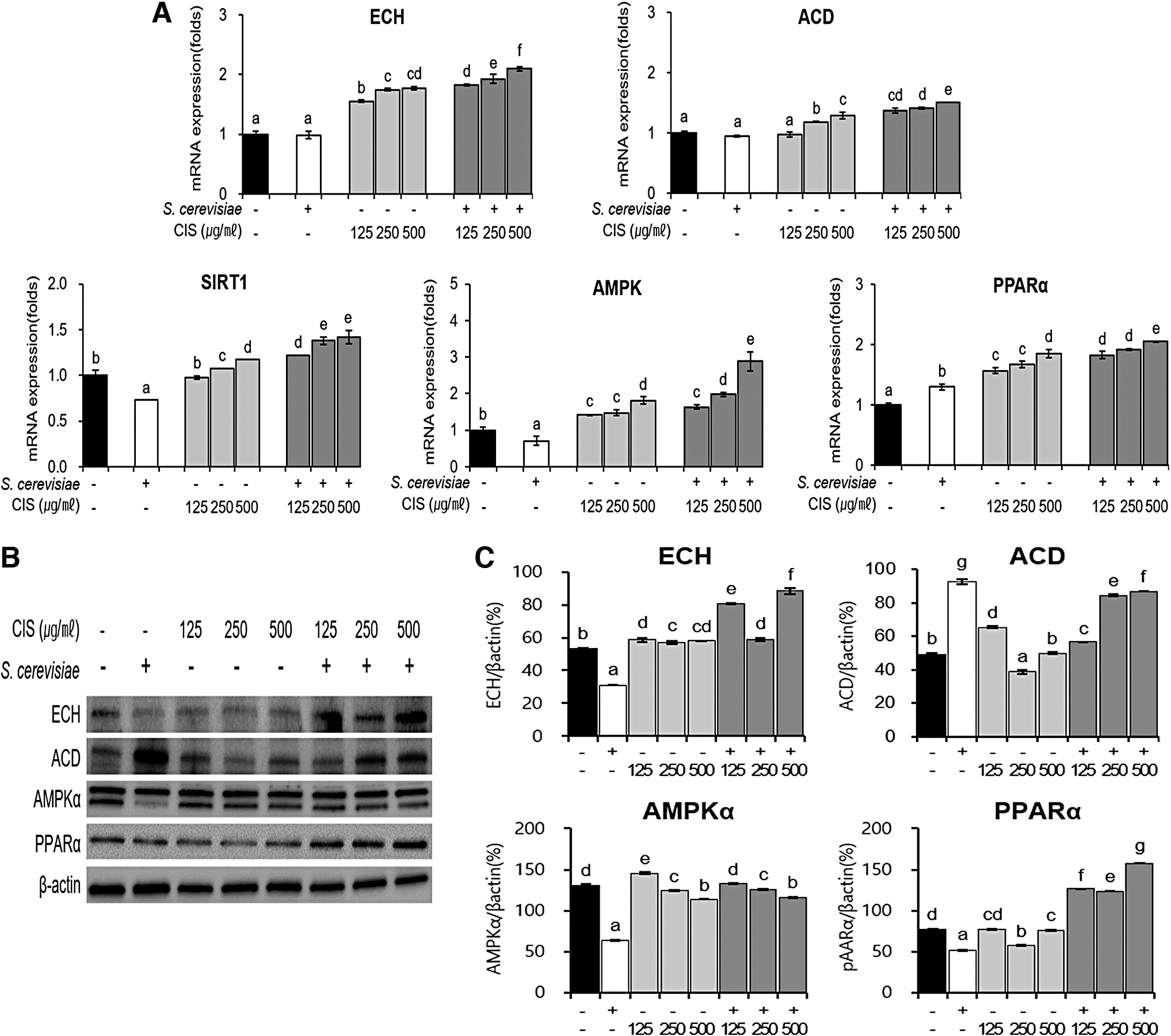
Effects of CIS extract fermented by 5% yeast (S. cerevisiae) on the expression of the genes associated with lipid β-oxidation and catabolism in C3H10T1/2 cells. After treatment of differentiated C3H10T1/2 cells with extracts or yeast for 24 h,
DISCUSSION
CIS is rich in phenols and flavonoids and exhibits beneficial physiological properties such as anti-aging and antioxidant effects. 26 Active constituents present in CIS include coumarin, gallic acid, and catechin, all of which are known to be effective against obesity. 24,29 In addition, reports have demonstrated the antioxidant and antihyperlipidemic properties of CIS extracts in animal models of high-fat diet-induced obesity. 30 With an aim to identify natural materials with anti-obesity functions, in this study we evaluated the anti-obesity effects of CIS and FCIS extracts and investigated their cytotoxicity to cells. We confirmed the increase in the proliferation of three different cell lines after treatment with these extracts. According to Niziol-Lukaszewska et al., 31 it was reported that high concentration of phenolic and flavonoid compounds was correlated with an increase in cell proliferation. Similarly, in our results, the cell proliferation increased more in FCIS extract than in CIS, which is thought to be owing to the increase in phenolic compounds such as gallic acid during fermentation.
FCIS extract affected the lipogenesis and adipogenesis-related TFs and the expression of fatty acid catabolism-related genes, thus preventing lipid accumulation. After confirming the absence of any cytotoxicity, the adipocytes were treated with CIS and FCIS extracts and stained with oil red O and Nile red to determine the degree of fat accumulation. Fat accumulation significantly decreased in differentiated adipocytes following treatment with CIS and FCIS extracts. In addition, the expression levels of the genes involved in adipogenesis and fat catabolism were regulated by CIS and FCIS extracts, thus mediating enhanced anti-obesity effects. The expression of adipogenesis-associated TFs such as C/EBPα and PPAR-γ increased following upregulation in the expression of C/EBPβ along with an increase in the expression levels of adipocyte-specific genes FAS and SCD-1. 32,33 Furthermore, the expression of SREBP-1c, which induces lipid synthesis, was changed. 34 CIS and FCIS extracts inhibited the expression of SREBP-1c, C/EBPα, and PPAR-γ mRNAs in differentiated adipocytes and downregulated the expression of FAS and SCD-1 mRNAs acting downstream of lipogenesis. The most potent effect was observed with 500 μg/mL FCIS, which was better than CIS extract. In contrast to the factors responsible for the production of fats, ACD and ECH levels related to β-oxidation of fatty acids dose dependently increased after extract treatment. Furthermore, the expression of SIRT1 and AMPK involved in the key signaling pathway underlying fat catabolism was upregulated in differentiated adipocytes treated with CIS and FICS extracts.
Together our results suggest that CIS and FICS extracts regulate C/EBPα and PPAR-γ expression by downregulating SREBP-1c expression. In addition, lipolysis is activated by increasing the levels of factors associated with fatty acid oxidization. These results are in line with the studies that showed inhibition of adipocyte differentiation following downregulation of SREBP-1c, PPAR-γ, and C/EBPα expression. 32 –34 Thus, fermented FCIS extract was more effective in suppressing obesity than CIS extract.
According to previous results, the anti-obesity effect of CIS is mediated by active ingredients such as gallic acid and ellagic acid. 24 Analyses of the components of FCIS extract by high-performance liquid chromatography revealed higher content of gallic acid in FCIS than in non-FCIS extract. 30 In a previous study, gallic acid fed to an obese mouse model for 9 weeks suppressed obesity by increasing the activity of lipid catabolism factors, AMPK/NAD-dependent deacetylase SIRT1/peroxisome proliferator-activated receptor gamma coactivator 1 alpha and uncoupling protein1. 35 These results indicate that a high gallic acid level exerts anti-obesity effects. Taken together, this study demonstrates that FCIS prevents obesity by reducing fat accumulation through the inhibition of adipogenesis and promotion of fat catabolism. In particular, the efficacy further improved after fermentation of CIS, probably owing to the increase in the content of gallic acid. Subsequent studies on the efficacy of the CIS, FCIS, and their functional components in vivo are in progress, it is expected that FCIS extract is a more effective raw material for preventing obesity than the CIS extract, and could be potentially developed as a therapeutic natural material for managing obesity.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This research was funded by BK21 FOUR (Fostering Outstanding Universities for Research) program and special research grant of Korea University.