
Abstract
Improvement of antioxidant and anti-inflammatory functions is believed to be an effective strategy for protection against various diseases such as cancer, aging, and neurodegenerative disease. This study focused on investigating antioxidant and anti-inflammatory abilities of Zingiber montanum oil (ZMO) extracted by the supercritical CO2 fluid system in HepG2 cells and lipopolysaccharide (LPS)-treated RAW 264.7 macrophages. Ten predominant constituents of ZMO were identified, in which triquinacene, 1,4-bis (methoxy), terpinen-4-ol, triquinacene, 1,4,7-tris (methoxy), α-terpinene, sabinene hydrate, and (E and Z)-1-(3,4-dimethoxyphenyl)butadiene account for 86.47%. ZMO exhibited anti-inflammatory capacity by inhibiting the formation of pro-inflammatory markers such as nitric oxide, inducible nitric oxide synthase, cyclooxygenase-2, interleukin (IL)-1β, IL-6, and monocyte chemoattractant protein-1 in LPS-treated macrophages. The LPS-induced stimulation of nuclear factor-kappa B, signal transducer and activator of transcription 3 (Stat3) and mitogen-activated protein kinase (MAPK) pathways as evident from increased phosphorylation of IKKα/β, IκBα, p65, Stat3, ERK, JNK, and p38 MAPK was also suppressed by ZMO pretreatment. Further, ZMO enhanced the expression of nuclear factor erythroid 2-related factor (Nrf2) and heme oxygenase-1 (HO-1), and concurrently, reduced intracellular reactive oxygen species accumulation in LPS-treated RAW 264.7 cells. In addition, ZMO treatment markedly upregulated the expression of Nrf2 as well as its target genes, HO-1 and NAD(P)H:quinone oxidoreductase 1 in HepG2 cells. These data propose that ZMO may be a potent candidate for prevention and/or treatment of inflammatory and oxidative conditions.
Introduction
An imbalance of redox homeostasis importantly contributes to the initiation and progression of numerous diseases such as atherosclerosis, inflammatory bowel disease, cardiovascular disease, and cancer. 1 Alterations in intracellular redox status are closely associated with an overall increase in the degree of reactive oxygen species (ROS), including superoxide anion radical, singlet oxygen, and hydroxyl radical. 2 Under physiological concentration, ROS act as signaling molecules mediating functional processes of cells; however, excessive ROS production directly and/or indirectly attacks DNA, lipids, and proteins, resulting in impaired cellular functions. 3 Along with the inflammatory stimuli, oxidative stress also triggers inflammatory response by modulating redox-sensitive signaling pathways in the inflammatory or immune cells. 4,5 Parallelly, acute and/or chronic inflammation contributes to the intercellular accumulation of ROS, reactive nitrogen species (RNS), as well as electrophiles that overwhelm cellular antioxidant defense systems. 6 Inflammation that accompanies oxidative damage is believed to aggravate pathological conditions.
Nuclear factor-kappa B (NF-κB), a redox-sensitive transcription factor, participates in regulating inflammation and multiple physiological and pathological processes such as innate immune response, cell growth and proliferation, apoptosis, and cancer. 5,6 Under the inflammatory stimuli such as lipopolysaccharide (LPS) and oxidative stress, NF-κB signal is one of the major mechanisms to produce numerous pro-inflammatory mediators and cytokines. Nuclear factor erythroid 2-related factor (Nrf2) is a key regulator of phase II detoxification and antioxidant genes that are the important members of cytoprotective systems against oxidative and xenobiotic damage. 7 –9 Recent epidemiological studies have supplied convincing evidence to support the assumption that Nrf2 contributes to the anti-inflammatory process through the interfering NF-κB pathway. 10,11 Therefore, the stimulation of the Nrf2 pathway as well as the inhibition of the NF-κB network might be a promising approach for the development of chemopreventive agents.
Zingiber montanum (J. Koenig) (syn. Zingiber cassumunar Roxb.), Zingiberaceae family, is a medicinal plant that grows in tropical and subtropical Asia such as India, Thailand, and Vietnam. Rhizome of Z. montanun is found in folk remedies for the therapy of several disorders such as inflammation, pain, and asthma. 12 Accumulative studies have reported many pharmacological properties of Z. montanum such as anti-inflammation, anti-microbes, anti-cholinesterase, and anti-ulcer. 13,14 These beneficial effects could be attributed to bioactive constituents in Z. montanum in an independent and/or synergistic manner. Although several pharmacological properties of Zingiber montanum oil (ZMO) have been studied, the potencies of ZMO and its molecular mechanisms for antioxidant and anti-inflammatory activities have not been fully investigated.
Especially, ZMO was extracted by using a supercritical CO2 fluid extraction, which can be regarded as an alternative procedure for conventional methods using organic solvents. 15 This system uses supercritical CO2 fluid as a solvent, which is nontoxic, recyclable, and environmental. In addition, operation is remained at low temperature during extraction to avoid the thermal degradation of bioactive constituents in samples. 16 –18 Therefore, the study was conducted to elucidate antioxidant and anti-inflammatory properties of ZMO and relevant mechanisms were clarified.
Materials and Methods
Chemicals
Dried powder of Z. montanum was obtained from a local market, Bangkok, Thailand. Sulforaphane (SFN), LPS (Escherichia coli O127:B8), sulfanilamide, phosphoric acid, and naphthyl ethylenediamine dihydrochloride were acquired from Sigma-Aldrich (St. Louis, MO, USA). Anti-cyclooxygenase-2 (COX-2), interleukin (IL)-1β, IL-6, Nrf2, NQO1 [NAD(P)H:quinone oxidoreductase 1], β-actin, and anti-mouse antibodies were acquired from Santa Cruz Biotechnology (Santa Cruz, CA, USA). Anti-p65, p-p65, signal transducer and activator of transcription 3 (Stat3), p-Stat3, heme oxygenase-1 (HO-1), monocyte chemoattractant protein-1 (MCP-1), p-extracellular signal-regulated kinase (ERK), ERK, p-stress-activated protein kinase/Jun-amino-terminal kinase (SAPK/JNK), SAPK/JNK, p-p38, p38, inducible nitric oxide synthase (iNOS), and anti-rabbit antibodies were purchased from Cell Signaling Technology (Beverly, MA, USA).
Preparation of Z. montanum extracts
Dried Z. montanum powder was extracted by using different solvents, including ethanol, hexane, and supercritical CO2 fluid. For ethanol and hexane extracts, the powder was extracted twice with 95% ethanol or hexane at room temperature. The homogenate of ethanol or hexane was separately filtered, and the filtrate of each solvent was concentrated by using a vacuum rotary evaporator to obtain Zingiber montanum ethanol extract (ZME) and Zingiber montanum hexane extract (ZMH), respectively.
For supercritical CO2 fluid extraction, dried Z. montanum powder was subjected to the extractor of the supercritical extraction system. Extraction with supercritical CO2 fluid was run at 6500 psi at 65°C. ZMO was harvested in an amber container and kept at −20°C.
Gas chromatography-mass spectrometry analysis
Chemical constituents of ZMO were analyzed by using a gas chromatography-mass spectrometry (GC-MS) (5975C; Agilent Technologies, Inc., Santa Clara, CA, USA) with an HP-5MS capillary column (30 m × 0.25 mm, 0.25 μm film thickness; Agilent Technologies, Inc.) according to a previous study. 19
Cell culture
Murine RAW 264.7 cells (ATCC, Rockville, MD, USA) were maintained in Dulbecco's modified Eagle medium (DMEM)-high glucose (Hyclone, Logan, UT, USA), containing 10% fetal bovine serum (FBS) and 1% penicillin/streptomycin solution at 37°C in a humidified CO2 incubator. Human hepatocarcinoma HepG2 cells (ATCC) were cultured in DMEM, supplemented 10% FBS, 1% essential amino acids, 1% Glutamax, and 1% penicillin/streptomycin solution at 37°C, 5% CO2 in a humidified atmosphere.
Cell viability
Cytotoxic effects of ZMO on HepG2 or RAW 264.7 cells were determined by using 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay. Briefly, the cells were maintained in medium containing MTT reagent (5 mg/mL, stock solution) for 4 h at 37°C. Formazan crystal was then solubilized in dimethyl sulfoxide, and the absorbance of solution was recorded at 570 nm with a multi-well spectrophotometer (BioTek, Winooski, VT, USA).
Nitrite assay
The level of nitric oxide (NO) was determined according to the Griess reaction. Briefly, mixtures of cell culture medium and an equal volume of Griess reagent were allowed to stand for 10 min at room temperature. The resulting colored solutions were quantified by measuring absorbance at 540 nm by using a multi-well spectrophotometer (BioTek).
Assay of intracellular ROS accumulation
Intracellular ROS accumulation was determined by using a cell-permeant 2′,7′-dichlorodihydrofluorescein diacetate (DCFH-DA). RAW 264.7 cells were prestained with DCFH-DA for 30 min before challenge with LPS (1 μg/mL) for another 12 h. After washing twice with phosphate buffered saline, dichlorofluorescein (DCF) fluorescence intensities were detected by a fluorescence microplate reader at 485/20 nm excitation and 528/20 mm emission (Synergy HT; BioTek).
Western blot analysis
Cells were lysed in an RIPA buffer, and protein contents were determined by using a BCA protein assay kit (Thermo Scientific, Rockford, IL, USA). The amounts of total proteins were determined by fractionating on sodium dodecyl sulfate polyacrylamide gels, followed by electrophoretically transferring onto polyvinylidene fluoride membranes. The membranes were then blocked with blocking solution (5% non-fat milk) and hybridized with specific primary antibodies for 24 h at 4°C, and this was followed by incubating them with appropriate secondary antibodies for 3 h at 4°C. Eventually, visualization was performed by using an enhanced chemiluminescence kit (Santa Cruz Biotechnology).
Statistical analysis
All data were displayed as the mean ± standard deviation. Comparisons between groups were performed by using one-way analysis of variance followed by Tukey's post hoc test. A P < .05 was regarded as statistically significant.
Results
Chemical compositions of ZMO
In this study, the yield of ZMO isolated by supercritical CO2 fluid extraction from rhizomes of Z. montanum was about 3% and the chemical composition of ZMO was analyzed by using GC-MS. Ten constituents, forming 100.0% of total oil compositions, were tentatively identified in ZMO, as listed in Table 1. According to GC-MS analysis, triquinacene, 1,4-bis (methoxy) (37.12%) was the most abundant compound in ZMO, followed by terpinen-4-ol (17.17%), triquinacene, 1,4,7-tris (methoxy) (9.60%), α-terpinene (8.44%), sabinene hydrate (7.50%), and (E and Z)-1-(3,4-dimethoxyphenyl)butadiene (6.64%). The remaining components, including terpinolene (4.94%), β-sesquiphellandrene (3.75%), ferulic acid methyl ester (3.25%), and ethanone, 1-[4-(1-methyl-2-propenyl)phenyl] (1.60%), were present at a relatively low concentration. Triquinacene, 1,4-bis (methoxy) and terpinen-4-ol were found to be major constituents, which account for 54.29% total oil compositions of ZMO.
Chemical Compositions of Zingiber montanum Oil Analyzed by Gas Chromatography-Mass Spectrometry
Inhibitory effects of Z. montanum extracts on NO formation in LPS-treated RAW 264.7 cells
Initially, we screened the anti-inflammatory capacity of different Z. montanum extracts by using LPS-treated RAW 264.7 cells. The cells were pre-incubated with ZME, ZMH, or ZMO ranging from 10 to 50 μg/mL for 1 h before LPS (1 μg/mL) challenge. As shown in Figure 1A, NO formation was markedly increased in culture medium after 12 h of LPS challenge when compared with the normal control. However, LPS-induced NO accumulation was significantly diminished by pretreatment of ZME, ZMH, or ZMO without cytotoxic effects at doses below 50 μg/mL (Fig. 1B). Among them, ZMO seemed to exhibit the greatest inhibitory effect on NO production compared with other extracts. Therefore, ZMO up to 50 μg/mL was applied for further experiments in this study.
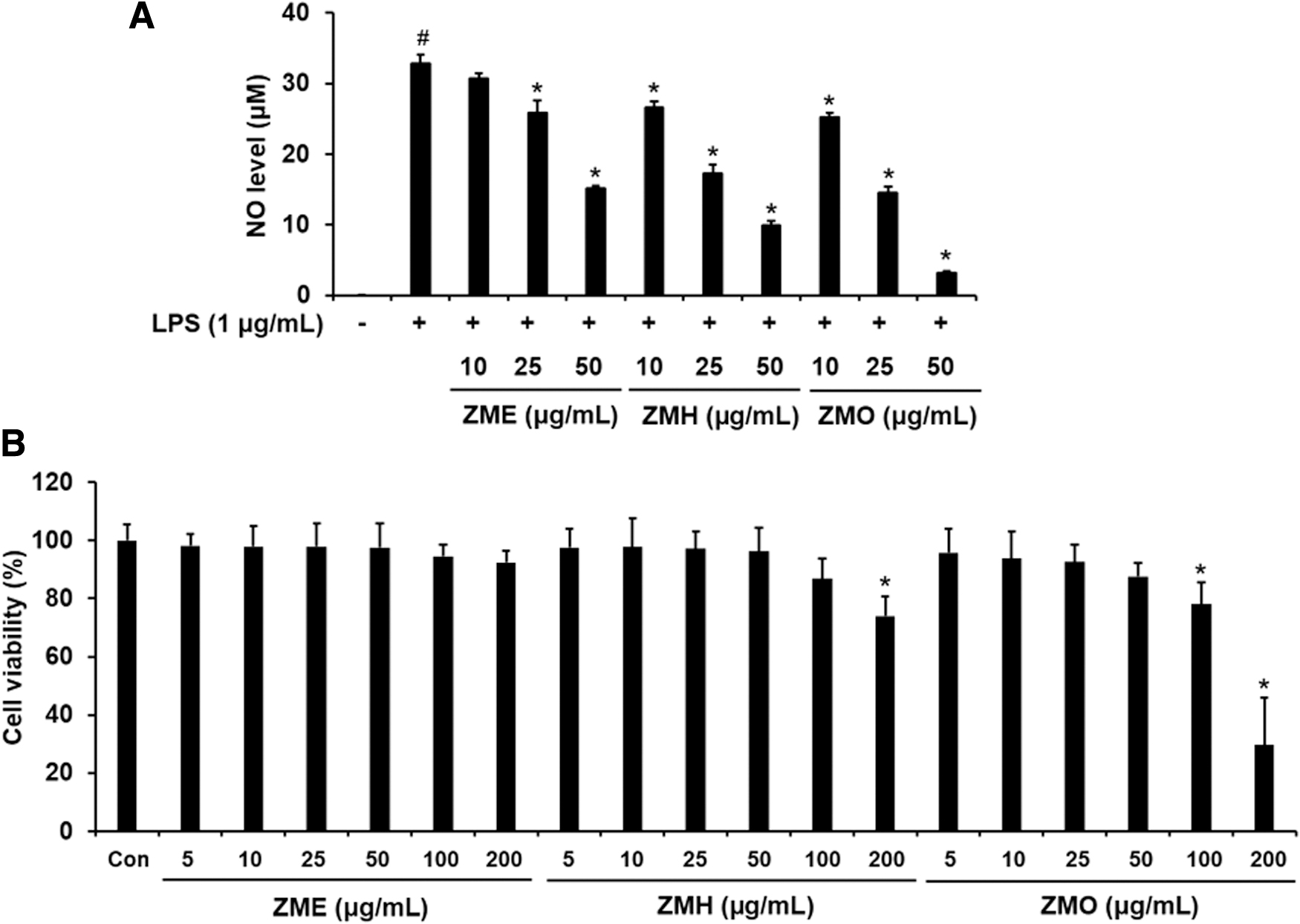
Effects of ZME, ZMH, and ZMO on cell viability and NO production in LPS-stimulated RAW 264.7 macrophages. The cells were pretreated with ZME, ZMH, or ZMO (10, 25, and 50 μg/mL) for 1 h before exposing them to LPS (1 μg/mL) for 12 h.
Inhibitory effects of Z. montanum extracts on the production of inflammatory markers in LPS-treated RAW 264.7 cells
To investigate whether or not the ZME, ZMH, and ZMO may inhibit inflammatory response, the level of pro-inflammatory makers was examined in LPS-stimulated macrophages. Compared with the untreated control, LPS stimulated the upregulation of iNOS, COX-2, and IL-1β. In contrast, cells treated with ZME, ZMH, or ZMO significantly reduced the expression of these proteins in LPS-treated RAW 264.7 macrophages. The pretreatment of cells with ZMH or ZMO at both concentrations (25 and 50 μg/mL) significantly decreased the levels of COX-2, iNOS, and IL-1β, whereas ZME at 25 μg/mL was unlikely to produce a significantly blocking effect on the expression of these proteins (Fig. 2). Moreover, ZMO seemed to exhibit a highest suppressing efficacy on the pro-inflammatory markers compared with other extracts. To confirm the role of ZMO, we continuously examined the expression level of several inflammatory markers. As expected, the suppressing effect of ZMO on the expression of COX-2, iNOS, IL-1β, IL-6, and MCP-1 was found in LPS-treated RAW 264.7 cells (Fig. 3).

Effects of ZME, ZMH, and ZMO on the expression of iNOS, COX-2, and IL-1β proteins in LPS-stimulated RAW 264.7 macrophages. The cells were pretreated with ZME, ZMH, or ZMO (25 and 50 μg/mL) for 1 h before exposing them to LPS (1 μg/mL) for 12 h. Protein levels of iNOS, COX-2
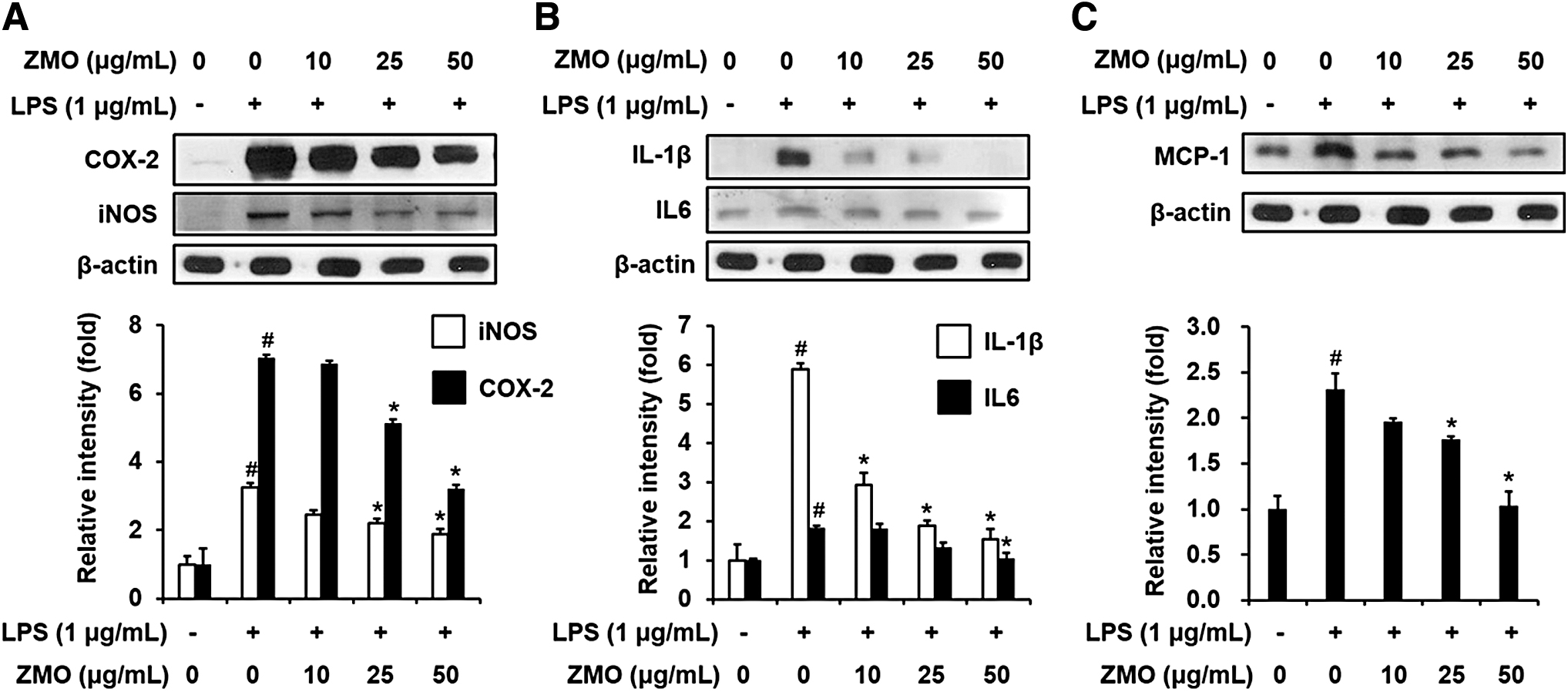
Concentration-dependent effect of ZMO on the expression of pro-inflammatory markers in LPS-stimulated RAW 264.7 macrophages. The cells were pretreated with ZMO (10, 25, and 50 μg/mL) for 1 h before exposing them to LPS (1 μg/mL) for 12 h. Protein levels of iNOS, COX-2
ZMO inhibits NF-κB, Stat3, and mitogen-activated protein kinase signaling pathways in LPS-treated RAW 264.7 cells
Abnormal activations of NF-κB, Stat3 and mitogen-activated protein kinase (MAPK) signaling pathways are crucial events in upregulating the transcription of pro-inflammatory genes. Thus, we hypothesized that the suppressive effect of ZMO on the generation of pro-inflammatory markers is associated with blocking these pathways. To test this hypothesis, RAW 264.7 macrophages were pretreated with ZMO before LPS challenge, and the expression of NF-κB and MAPK pathways-related proteins was determined. As indicated in Figure 4A, the phosphorylation level of IKKα/β [IkappaB (inhibitor of NF-kappaB) kinase alpha/beta] was remarkably induced by LPS stimulation when compared with the untreated control. However, the LPS-induced phosphorylation status of IKKα/β was markedly diminished in the cells pretreated with ZMO. In addition, LPS also enhanced IκBα phosphorylation, which was reversed by the presence of ZMO (Fig. 4B). Further results demonstrated that ZMO produced a blocking effect on p65 phosphorylation in LPS-treated RAW 264.7 cells (Fig. 4C). Along with the NF-κB pathway, LPS-provoked phosphorylation of Stat3 was also suppressed by ZMO pretreatment (Fig. 4D). Besides the NF-κB and Stat3 pathways, we determined whether MAPK signaling pathways are required for ZMO-mediated downregulation of pro-inflammatory proteins. Data showed that LPS markedly activated the phosphorylation of ERK, SAPK/JNK, and p38 MAPK. In contrast, highly phosphorylated levels of MAPKs were significantly reduced by ZMO pretreatment in LPS-treated macrophages, suggesting a requirement for the involvement of MAPK signaling pathways for ZMO-mediated activity (Fig. 4E–G). Overall, these data demonstrate that ZMO could exert anti-inflammatory activity through blocking not only NF-κB and Stat3 networks but also MAPK signaling pathways.
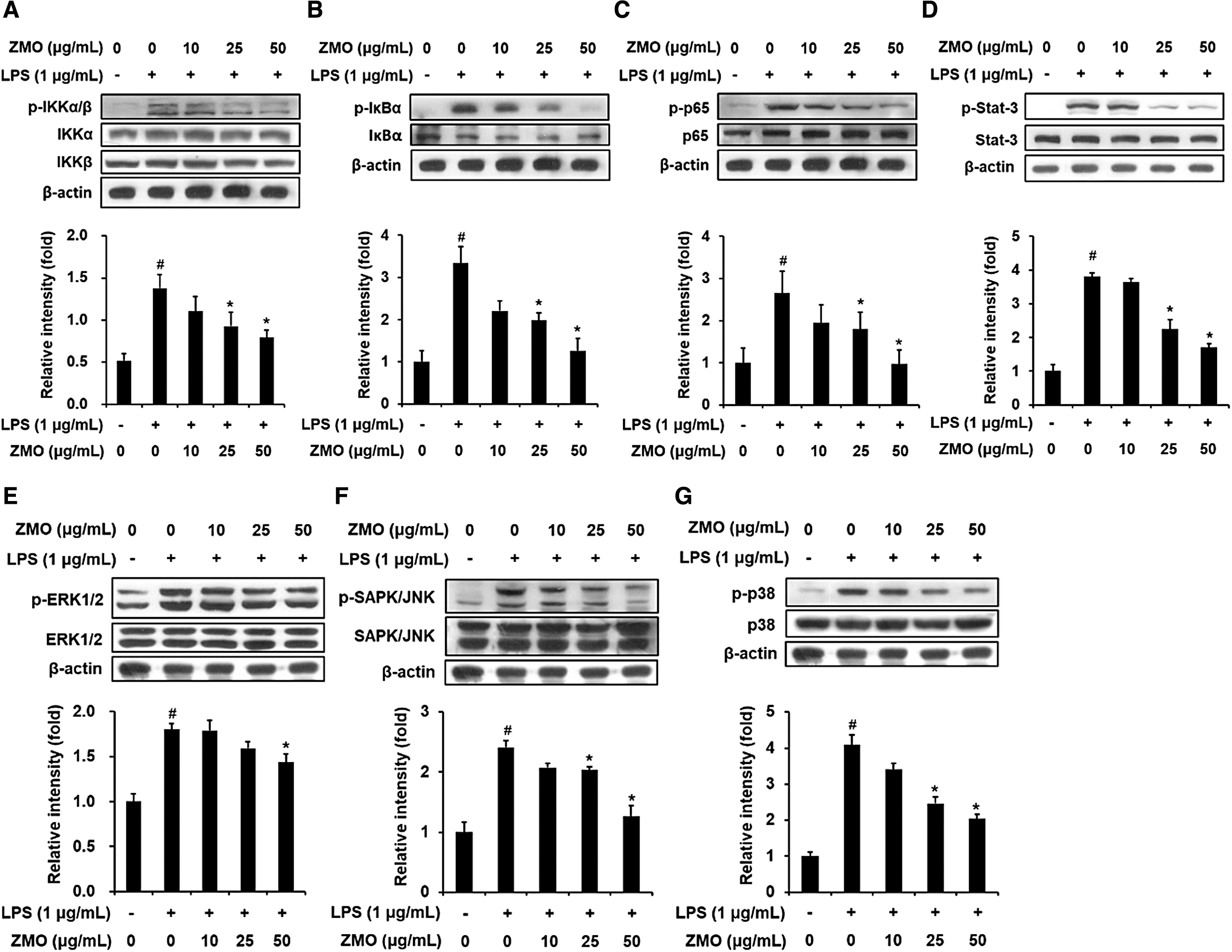
Effect of ZMO on the NF-κB, Stat3, and MAPK signaling pathways in LPS-stimulated RAW 264.7 macrophages. The cells were pretreated with ZMO (10, 25, and 50 μg/mL) for 1 h before exposing them to LPS (1 μg/mL). Protein expression was analyzed by Western blotting. The protein levels of NF-κB-related genes, including p-IKKα/β and IKKα/β
ZMO induces Nrf2-driven HO-1 expression in LPS-treated RAW 264.7 cells
We further examined the role of ZMO in activating Nrf2-phase II antioxidant pathway in LPS-treated RAW 264.7 cells. As illustrated in Figure 5A, ZMO pretreatment remarkably enhanced Nrf2 expression, whereas LPS challenge did not alter the expression level of this protein, compared with the vehicle-treated control. In addition, an antioxidant enzyme HO-1, an Nrf2 target gene, was markedly augmented by the ZMO pretreatment in a dose-dependent manner (Fig. 5B). Consequently, ZMO pretreatment considerably reduced LPS-induced intracellular ROS production in macrophages (Fig. 5C). These data suggest that the enhancement of Nrf2-mediated phase II antioxidant enzyme may contribute to the anti-inflammatory activity of ZMO.

Effect of ZMO on the Nrf2-mediated phase II antioxidant enzyme and intracellular ROS accumulation in LPS-stimulated RAW 264.7 macrophages. The cells were pretreated with ZMO (10, 25, and 50 μg/mL) for 1 h before exposing them to LPS (1 μg/mL). Protein levels of Nrf2
ZMO induces Nrf2-driven phase II detoxifying/antioxidant enzyme expression in HepG2 cells
HepG2 cell line is a reliable model for defining novel botanical agents that are capable of inducing cytoprotective systems and ameliorating the adverse effects of oxidative stress. Thus, HepG2 cells were selected in the present study. To elucidate the cytotoxic ability of ZMO, HepG2 cells were incubated with ZMO at a series of concentrations (0–100 μg/mL) for 24 h, and the level of cell viability was determined by using MTT assay. As illustrated in Figure 6A, ZMO exhibited a dose-dependent cytotoxic effect on cell survival. The viability of HepG2 cells was reduced about 20% by 100 μg/mL of ZMO, whereas concentrations of ZMO up to 50 μg/mL were unlikely to cause a significant reduction in the cell survival.
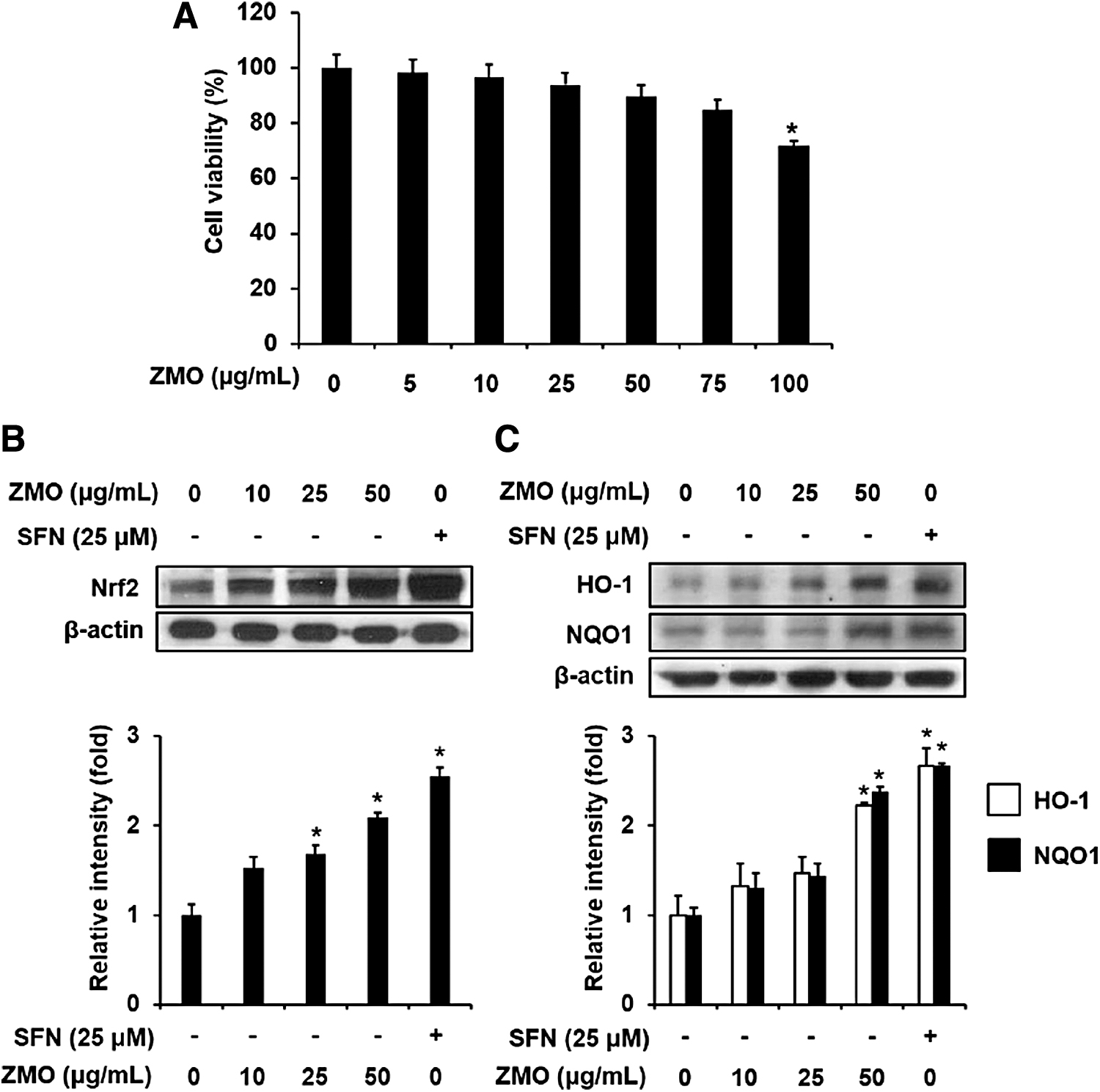
Effect of ZMO on the cell viability and the expression of Nrf2-mediated phase II detoxifying/antioxidant enzymes in HepG2 cells. The cells were treated with ZMO (10, 25, and 50 μg/mL) or SFN (positive control, 25 μM) for 24 h.
The Nrf2 network is one of the cytoprotective systems that protect against oxidative stress and carcinogens. Thus, we further explored the inductive activity of ZMO on the protein expression of Nrf2 pathway-related genes in HepG2 cells. Results showed that ZMO treatment significantly elevated the protein level of Nrf2, compared with the vehicle treatment only (Fig. 6B). The efficacy of ZMO at 50 μg/mL was comparable to that of SFN (25 μM), a well-established Nrf2 inducer. Accordingly, ZMO upregulated the expression of phase II detoxification and antioxidant enzymes, including NQO1 and HO-1 (Fig. 6C). Based on these findings, we propose that ZMO may enhance cellular defense systems, possibly through activating the Nrf2 pathway, suggesting a novel potent Nrf2 inducer.
Discussion
Our study showed that different Z. montanum extracts, including ZME, ZMH, and ZMO, exhibited blocking effects on NO production in LPS-treated macrophages. NO is well known as a signaling molecule that plays essential roles in vasodilation, neurotransmission, and immune defense through modifying the function of immune cells and resident cells of different tissues and organs. 20 However, NO also reflects the inflammatory process and importantly contributes to the development of various diseases, whereas its overproduction is mainly required by iNOS in response to inflammatory factors. 21,22 LPS (1 μg/mL) challenge remarkably upregulated the expression of iNOS, which was inhibited by the pretreatment of Z. montanum extracts. In addition, Z. montanum extracts pretreatment also reduced protein expression of COX-2 and IL-1β in the presence of LPS, suggesting that all three extracts of Z. montanum exhibited anti-inflammatory activities via blocking the expression of pro-inflammatory genes.
It is worth noting that among three Z. montanum extracts, ZMO exhibited the greatest inhibitory effects on NO production, pro-inflammatory gene expression, and ROS accumulation in LPS-treated macrophages without cytotoxicity. These findings imply that the majority of bioactive compounds in ZMO also have both anti-inflammatory and antioxidant activities, thus we attempted to identify the chemical compositions of ZMO. GC-MS analysis showed that triquinacene, 1,4-bis (methoxy), terpinen-4-ol, α-terpinene, sabinene hydrate, and (E and Z)-1-(3,4-dimethoxyphenyl)butadiene were major constituents in ZMO. A previous study indicated the antioxidant activity of total volatile oil of Z. montanum, which was possibly attributed to its major bioactive compounds such as sabinene, terpinen-4-ol, and (E)-1-(3′,4′-dimethylphenyl) butadiene. 23 Although the biological activities of triquinacene, 1,4-bis (methoxy) have not been reported, terpinen-4-ol has been demonstrated to exert antioxidant and anti-inflammatory properties. It has been indicated that terpinen-4-ol from tea tree oil (an essential oil of Melaleuca alternifolia) suppressed the generation of superoxide as well as pro-inflammatory mediators and cytokines such as TNF-α, IL-1β, IL-8, IL-10, and PGE2 in LPS-stimulated human peripheral blood monocytes. 24,25 Terpinen-4-ol treatment also significantly attenuated the inflammatory response in LPS-treated macrophages and dextran sodium sulfate (DSS)-induced colitis through suppressing pro-inflammatory cytokine secretion and NLRP3 (nucleotide-binding domain, leucine-rich-containing family, pyrin domain-containing-3 or Nod-like receptor protein 3) inflammasone. 26 In addition, phenylbutenoid derivatives present in ZMO, such as (E and Z)-1-(3,4-dimethoxyphenyl)butadiene, have also been documented to exert anti-inflammatory properties in in vitro and in vivo models. 27,28 These reports suggest that bioactive compounds contribute to the anti-inflammatory property of ZMO. However, little studies on the antioxidant and anti-inflammatory properties of ZMO extracted by supercritical CO2 fluid extraction as well as its major components have been documented.
Transcription factor NF-κB is involved in many aspects of innate and adaptive immune processes and plays a crucial role in the onset of inflammation and tumor progression through controlling the production of pro-inflammatory factors. 29 In normal condition, NF-κB is anchored in the cytoplasm in an inactive form by forming a complex with its inhibitor IκBα. When cells are exposed to inflammatory stimuli, the IKK complex is activated, leading to the phosphorylation and subsequent degradation of IκB protein. As a result, free NF-κB migrates to the nucleus, where it drives the transcription of a wide range of inflammatory genes such as iNOS, COX-2, IL-1β, and IL-6. 29 Earlier studies have demonstrated that LPS induces the inflammatory response through activating the NF-κB pathway; in contrast, various bioactive compounds exhibit inhibitory effects on LPS-mediated NF-κB activation in macrophages. 22,30 This study confirmed the ability of LPS in the activation of the NF-κB pathway, as evident from increased phosphorylation levels of IKKα/β, IκBα, and p65/NF-κB. However, ZMO pretreatment significantly decreased phosphorylation of these proteins, suggesting that ZMO could mediate some anti-inflammatory activities in an NF-κB-dependent manner. In addition to the NF-κB pathway, Stat3 pathway also participates in the LPS-triggered inflammatory response in macrophages. In the presence of LPS, Sat3 transcription factor is stabilized and then relocated to the nucleus to initiate the expression of target genes, especially iNOS, IL-1β, and IL-6. 31,32 Our data showed a reduced level of Stat3 phosphorylation by ZMO pretreatment in LPS-treated macrophages. Overall, these findings suggest that ZMO pretreatment suppresses both NF-κB and Stat3 pathways, thereby mitigating LPS-stimulated inflammatory response in macrophages.
The MAPKs, consisting of ERK1/2, SAPK/JNK, and p38 MAPK, belong to a large family of serine/threonine kinases that can transduce extracellular signals to create multiple cellular responses. 33 Downstream signaling pathways of MAPKs are implicated in various cellular functions such as inflammation, apoptosis, and cell proliferation in response to a wide range of stimuli such as cytokines, oxidative stress, and chemicals. It has been demonstrated that MAP kinases can mediate the inflammatory responses from LPS stimulation. 34 On extracellular stimulation, protein kinases are activated and thus they can modulate signaling cascades by phosphorylating serine/threonine amino acids on their target proteins. Eventually, various transcription factors existing in the cytoplasm can be stabilized and activated by the phosphorylation of the MAP kinases such as ERK1/2, JNK1/2, and p38 MAPK, leading to their nuclear translocation and transcriptional expression of inflammatory genes. 22 Our study indicated that ZMO pretreatment suppressed MAPK signals by blocking the LPS-stimulated phosphorylation of JNK and p38 MAPK. Along with the observations cited earlier, these findings suggest that ZMO may attenuate the LPS-stimulated inflammatory response through inhibiting JNK/p38 MAPK signal-mediated NF-κB and Stat3 pathways.
Oxidative stress plays an important role in inflammation. ROS over-production can trigger macrophage activation and enhance the generation of pro-inflammatory factors and cytokines. In addition, excessive production of ROS/RNS under inflammatory states not only alters intracellular signaling pathways but also attenuates cytoprotective systems. 3 It is well known that an LPS challenge dramatically increases the level of intracellular ROS in macrophages and initiates an inflammatory response. 35,36 Therefore, suppressing the accumulation of intracellular ROS can inhibit pro-inflammatory signaling. Our study denoted that ZMO was highly effective at decreasing LPS-triggered ROS production in macrophages.
Nrf2 signaling regulates the cellular defense system in response to oxidative stress as well as inflammation and that is believed to impede NF-κB activation. 37 It plays an important function in cellular redox equilibrium through transcriptionally inducing phase II detoxification and antioxidant proteins such as NQO1, HO-1, and glutathione-S transferase. This study showed an elevated level of Nrf2 and HO-1, an Nrf2 target gene, by ZMO in LPS-treated RAW 264.7 cells, suggesting an enhanced cytoprotective system. Phase II antioxidant enzyme HO-1 importantly contributes to protecting cells/tissues from extra and/or intracellular hazards through antioxidant and anti-inflammatory capacities. 3,38 Multiple lines of evidence have indicated that HO-1 and its enzymatic byproducts carbon monoxide and biliverdin exert cytoprotective abilities through maintaining redox homeostasis, upregulating the excretion of anti-inflammatory IL-10, and interfering with the generation of pro-inflammatory proteins such as iNOS, IL-1β, and IL-6. 39 –41 Further observations have indicated that upregulation of HO-1 could specifically block NF-κB phosphorylation via the action of free iron. 42 Therefore, the enhanced Nrf2/HO-1 axis can be an effective approach to counteract oxidative stress and inflammation-associated diseases. As expected, our study provided the first evidence that the Nrf2-dependent antioxidative mechanism may be required for ZMO-mediated anti-inflammation in LPS-treated macrophages. Thus, ZMO exerts its anti-inflammatory property via dual mechanisms as an Nrf2 activator and NF-κB suppressor.
In addition, we also reported that ZMO remarkably enhanced the expression of Nrf2-mediated phase II enzymes, NQO1 and HO-1 in the human liver HepG2 cell line. Along with HO-1, the detoxifying enzyme NQO1 is a prominent target gene of Nrf2 and it plays an important function in the protection of cells/tissues from oxidative stress as well as toxicants. 43 NQO1 is considered as a leading enzyme of the cellular defense system by providing multiple cytoprotective functions. Recent studies been obviously proved that NQO1 exerts antioxidant activity by its catalytic mechanism of two-electron reduction of quinone compounds to hydroquinones, thereby averting certain quinones from becoming involved in oxidative cycling and the formation of reactive intermediates. 43 –45 Phase II detoxifying enzymes contribute to the removal of oxidants and/or toxicants from cells before they can damage cells, and thus they prevent the initiation of various diseases, including cancer. The functional role of NQO1 in counteracting oxidative stress has been supported by evidence that overexpression or depletion of NQO1 level leads to the reduced or enhanced susceptibility to oxidative damage, respectively. 45 Therefore, the induction of detoxification and antioxidant enzymes may be considered a primary prevention approach to multiple diseases.
Overall, our study provides more information about the antioxidant and anti-inflammatory properties of ZMO obtained by supercritical CO2 fluid extraction. ZMO exhibits anti-inflammatory activity, as evident from reduced levels of pro-inflammatory markers. The inhibition of signaling networks such as NF-κB, Stat3, and MAPK was demonstrated to be a mechanism for ZMO-mediated anti-inflammatory properties. In addition, ZMO also enhances cytoprotective systems in HepG2 cells and in LPS-treated RAW 264.7 cells. The antioxidant activity of ZMO is shown by the decreased level of LPS-induced intracellular ROS accumulation. Moreover, this study indicates the chemical constituents of supercritical CO2 fluid extracted essential oil from the rhizome of Z. montanum. Therefore, ZMO may be developed as a potent functional product with antioxidant and anti-inflammatory properties.
Footnotes
Authors' Contributions
Designed and performed experiments, V.-L.T. and T.-T.P.; statistical analysis, V.-L.T. and B.M.; validation and data curation, V.-L.T. and W.-S.J.; writing—original draft preparation, V.L.T.; writing—review and editing, V.-L.T., B.M., and W.-S.J.; supervision, W.-S.J.
Author Disclosure Statement
All authors declare no conflict of interest.
Funding Information
This research was supported by the Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education (NRF-2017R1D1A1B03031112, NRF-2020R1F1A1073595, and NRF-2021R1A2C2006745).