
Abstract
We investigated whether a standardized saw palmetto extract (SP, mixture of supercritical extract and ethanol extract at a ratio of 9.5 to 0.5) can relieve the symptoms of andropause, including metabolic syndrome, and decreases in muscle endurance and spermatogenesis, in old rats. Twenty-four-week-old male Sprague Dawley rats received oral supplementation of SP at 40, 80, and 160 mg/kg body weight (bw) for 4 weeks. We found that SP supplementation reduced body weight gain by decreasing visceral and epididymal fat weights and the levels of serum triglycerides, total cholesterol, and low-density lipoprotein/very low-density lipoprotein cholesterol. In addition, SP supplementation increased muscle endurance, sperm counts, and testosterone biosynthesis through hormonal regulation. In Leydig cells under hydrogen peroxide-induced oxidative stress, SP treatment directly induced testosterone biosynthesis by activating the mRNA expression of the genes encoding 17,20-desmolase and 3β-hydroxysteroid dehydrogenase 4. In conclusion, our results suggest that supplementation of SP may be useful for alleviating the symptoms of andropause via direct and indirect regulation of testosterone biosynthesis.
Introduction
Andropause, the male equivalent of menopause, is the set of symptoms caused by the age-related deficiency in male hormones that begins to occur in men in their late 40s to early 50s. It is known that total testosterone and free testosterone decrease by 0.4% and 1.2% each year, respectively; thus, aging is a major cause of andropause. 1 The symptoms of andropause include physical, psychological, and sexual problems, such as fatigue, increased body fat, decreased muscle strength and sexual function, depression, and memory loss. 1 –4
The most abundant androgen in men is testosterone, which plays important roles in fertility via spermatogenesis, muscle mass, and fat distribution. 5 Testosterone is produced and secreted by the Leydig cells of the testis during spermatogenesis in response to luteinizing hormone (LH). 6 Gonadotropin-releasing hormone, which is secreted by the hypothalamus, stimulates the production of LH and follicle stimulating hormone (FSH) in the pituitary. FSH indirectly regulates the production and maintenance of sperm through the somatic Sertoli cells. 7 Circulating LH binds to the LH receptor, which stimulates a signaling cascade that promotes cholesterol transport from the cytoplasm into the mitochondria. The cholesterol in the inner mitochondrial membrane is converted to pregnenolone, which is transported to the smooth endoplasmic reticulum where it is converted to progesterone by 3β-hydroxysteroid dehydrogenase (3β-HSD). Progesterone is converted to testosterone by cytochrome P450 17α-hydroxylase/C17-20, lyase (P450c17) and 17β-hydroxysteroid dehydrogenase (17β-HSD). 8 –10 Thus, damage to Leydig cells causes andropausal symptoms via inhibition of testosterone biosynthesis and spermatogenesis.
Several studies have shown that testosterone replacement therapy can improve the symptoms of andropause. However, the use of testosterone replacement therapy is limited by its liver toxicity and the potential risks of prostate cancer, cardiovascular diseases, and skin diseases. 11 Thus, other therapeutic agents need to be considered. Various natural materials have been explored as alternatives to avoid the possible risks and side effects of testosterone replacement therapy. 12,13 In this study, we evaluated the potential of saw palmetto extract (SP) for relieving andropause symptoms using old rats and Leydig cells under H2O2-induced oxidative stress as models. Saw palmetto is mostly used to treat benign prostatic hyperplasia. However, the exact mechanism underlying the effect of saw palmetto on benign prostatic hyperplasia is unknown, and its potential effects on andropause symptoms have not been studied. 14 –16 Therefore, we investigated whether SP can relieve the symptoms of andropause in old-aged rats with naturally induced andropause and whether it has a protective effect against H2O2-induced oxidative stress in Leydig cells.
Materials and Methods
Extract preparation
Saw palmetto was extracted using CO2 (50–60°C, 18–20 h; supercritical extraction) and ethanol (70–80°C, 5–6 h; ethanol extraction). The supercritical and ethanol extracts were mixed at a ratio of 9.5:0.5. This mixture (SP) was standardized by lauric acid 220–360 mg/g and β-sitosterol 20–35 mg/g (Fig. 1) and stored in an air-tight container at −20°C until use.
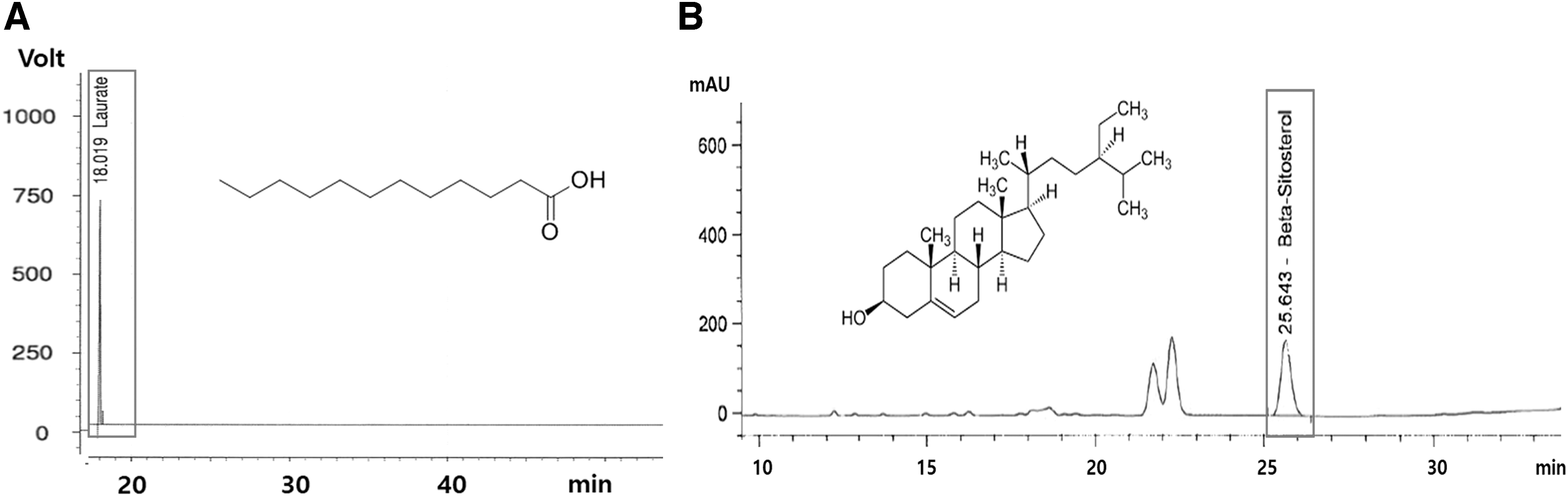
High-performance liquid chromatography analysis of lauric acid
Animal experiment
Male 24-week-old Sprague Dawley rats were obtained from Japan SLC, Inc., The experimental protocol was approved by the animal care and use review committee of Kyung Hee University (KHGASP-20-409). The rats were housed in an environmentally controlled facility (22°C ± 2°C, 50–60% humidity, and a 12-h light/dark cycle) and fed an AIN 93G diet for an acclimation period of 7 days. Then, the rats were randomly divided into four groups: the normal control (NC), SP 40 (orally supplemented with SP at 40 mg/kg bw), SP 80 (orally supplemented with SP at 80 mg/kg bw), and SP 160 (orally supplemented with SP at 160 mg/kg bw) groups. Four weeks after SP administration, the rats were euthanized via cervical dislocation.
Analysis of lipid profiles, aspartate transaminase, and prostate-specific antigen
Serum concentrations of low-density lipoprotein (LDL)/very low-density lipoprotein (VLDL) cholesterol, high-density lipoprotein cholesterol, total cholesterol (TC), triglyceride (TG), and aspartate transaminase (AST) were measured using enzyme assay kits (Biovision, Inc., Mountainview, CA, USA). The serum concentration of prostate-specific antigen (PSA) was measured using an enzyme assay kit (Cusabio, Wuhan, China).
Forced swimming experiment
A forced swimming experiment was performed to assess muscle endurance. A clear acrylic plexiglass chamber (50 cm [W] × 40 cm [D] × 60 cm [H]) was filled with water up to 40 cm so that the rat's tail would not touch the bottom of the chamber. Swimming time was defined as the exhausted swimming time, which is the time point at which the rat became exhausted and sank below the surface of the water for 2 sec five times.
Counting the number of sperm and measuring sperm activity
The epididymis was harvested from the rats and placed in M199 medium containing 0.5% bovine serum albumin. The sperm-containing fluid was incubated in a water bath at 37°C for 10 min. A hemocytometer was used to determine the number of spermatozoa in the fluid, and spermatozoa that were motile, that is, moved even a little for 3 sec, were considered active.
Measurement of serum total testosterone, free testosterone, sex hormone binding globulin, 5α-reductase, growth hormone, insulin like growth factor 1, LH, and FSH
Serum samples were obtained from the rats, and the levels of total testosterone, free testosterone, sex hormone binding globulin (SHBG), 5α-reductase, and growth hormone (GH) were determined using ELISA kits obtained from MyBioSource (San Diego, CA, USA), and insulin like growth factor 1 (IGF-1), LH, and FSH levels were determined using ELISA kits from LifeSpan Biosciences (Seattle, WA, USA).
Cell culture and treatments
TM3 Leydig cells were obtained from American Type Culture Collection (ATCC, Manassas, VA, USA). The cells were cultured in Dulbecco's minimal essential medium (DMEM; Hyclone Laboratories, Logan, UT, USA) supplemented with 10% fetal bovine serum (FBS; Hyclone Laboratories), 100 mg/L penicillin–streptomycin, and 2 mmol/L glutamine (Hyclone Laboratories) at 37°C in a humid atmosphere of 5% CO2. After treatment with 400 μM H2O2 for 1 h, assays were performed to measure cell viability, testosterone, and 5α-reductase levels and the mRNA expression levels related to testosterone metabolism.
mRNA extraction and reverse transcription polymerase chain reaction
Total mRNA was extracted from cells and tissues using the RNeasy Mini Kit (QIAGEN, Germantown, MD, USA). Then, cDNA was synthesized using the iScript™ cDNA Synthesis Kit (Bio-Rad, Hercules, CA, USA), and RT-PCR analysis was performed using SYBR Green PCR Master Mix (iQ SYBR Green Supermix; Bio-Rad) under the following cycling conditions: 35 cycles of denaturation at 95°C for 15 sec, annealing at 58°C for 30 sec, and extension at 72°C for 45 sec. The following primers were used: GAPDH (mouse, M) forward 5′-CCC CAC ACA CAT GCA CTT ACC-3′ and reverse 5′-TTG CCA AGT TGC CTG TCC TT-3; 3β-HSD4 (M) forward 5′-CCA CAC CGC TGC TGC TAT T-3′ and reverse 5′-GAC ATC TAG GAT GGT CTG TCT GGA A-3′; 17,20-desmorase (M) forward 5′-CCC TGG CTG TAG CTC TGA TGT-3′ and reverse 5′-GCT TCG GAG AGA CAA GTC AAT CTT-3′; aromatase (M) forward 5′-GAT GCT GGC CTT GGC TGT AG-3′ and reverse 5′-GTT GGT GTG CTG ACC AGA ACA-3′; 5α-reductase (M) forward 5′-TAG CGT CGG TGT CTT CTT CTT TAT T-3′ and reverse 5′-CTG AGC TGG CGC AGC AT-3′; GAPDH (rats, R) forward F 5′-CCC AAC CTG CTC CCA AGT C-3′ and reverse 5′-GTG GAT GAC ACT CTG GCT CTC A-3′; 17β-HSD3 (R) forward F 5′-GAA AGG CGG GAT ACG AAA AGA-3′ and reverse 5′-TCT GAC GCT TGT GCT TCA ACA-3′; 3β-HSD2 (R) forward 5′-AGG CCT GTG TCC AAG CTA GTG T-3′ and reverse 5′-GCC CTG CAA CGT CAA CTG T-3′; 5α-reductase (R) forward 5′-CTA CCT GGT TTA CTG CGC AGA A-3′ and reverse 5′-ACC AAA GCT AAA CCG CAC ATC T-3′.
Statistical analysis
The experimental results are presented as the mean ± standard deviation. The data were analyzed by one-way analysis of variance or Student's t-test using SPSS statistical procedures for Windows (SPSS PASW Statistic 23.0; SPSS, Inc., Chicago, IL, USA). Duncan's multiple range test was used to determine significance of differences. A P value <.05 was considered significant.
Results
Effects of SP on body and tissue weights in old male rats
Conditions associated with metabolic syndrome, such as obesity, abnormal lipid and glucose metabolism, and cardiovascular disease, are common in men after middle age and can be caused by decreased testosterone levels. 2 We measured the body and tissue weights of old male rats treated with SP to assess whether supplementation with SP affects weight gain and tissue weight. The results in Table 1 show that supplementation of SP led to significant decreases in weight gain, food efficiency ratio, and fat tissue weight in old male rats. However, supplementation of SP did not affect the weights of other tissues in the rats (P < .05). These results suggest that SP can reduce body fat without side effects.
Changes in Body and Tissue Weights in Old Rats With and Without Saw Palmetto Extract Supplementation
NC, SP 40: orally supplemented with SP at 40 mg/kg bw, SP 80: orally supplemented with SP at 80 mg/kg bw, and SP 160: orally supplemented with SP at 160 mg/kg bw. Values are presented as means ± SD. Different letters indicate a significant difference at P < .05 as determined using Duncan's multiple range test.
NC, normal control; SD, standard deviation; SP, saw palmetto extract.
Effects of SP on lipid profiles, AST, and PSA levels in old male rats
Serum levels of TG, TC, and LDL/VLDL cholesterol decreased significantly in the SP supplementation groups compared with the corresponding levels in the NC group (P < .05). However, there were no significant differences in the serum levels of AST and PSA among the groups (Table 2). Therefore, we suggest that supplementation of SP does not induce hepatotoxicity or prostate toxic effects.
Serum Lipid Profiles and Aspartate Transaminase and Prostate-Specific Antigen Levels in Old Rats With and Without Saw Palmetto Extract Supplementation
NC, SP 40: orally supplemented with SP at 40 mg/kg bw, SP 80: orally supplemented with SP at 80 mg/kg bw, and SP 160: orally supplemented with SP 160 at mg/kg bw. Values are presented as means ± SD. Different letters indicate a significant difference at P < .05 as determined using Duncan's multiple range test.
AST, aspartate transaminase; HDL, high-density lipoprotein; PSA, prostate-specific antigen; LDL/VLDL, low-density lipoprotein/very low-density lipoprotein; TC, total cholesterol; TG, triglyceride.
Effects of SP on muscle endurance in old male rats
Since decreased muscle endurance is a common physical symptom of andropause syndrome, 17 we performed a forced swimming experiment to assess the effect of SP on muscle endurance and motor performance. When compared with the NC group, the groups with SP supplementation had significantly longer swimming times (P < .05; Fig. 2A). In addition, supplementation of SP led to a significant increase in the total sperm and active sperm counts when compared with that of the NC group (P < .05; Fig. 2B, C).

Muscle endurance as measured by a forced swimming experiment
Effects of SP on hormone levels in old male rats
We found that SP supplementation induced significant increases in serum total and free testosterone levels and a significant decrease in SHBG level compared with the corresponding level in the NC group. In addition, the SP-supplemented groups had lower serum 5α-reductase levels than the NC group (P < .05; Fig. 3A–D).

Serum total testosterone
We next investigated whether SP supplementation affects the production of testosterone-related hormones in old rats. The SP-supplemented groups showed significant increases in GH, IGF-1, LH, and FSH compared with the corresponding levels in the NC group (P < .05; Fig. 3E–H).
Effects of SP on the mRNA expression of enzymes involved in testosterone biosynthesis in the testis of old male rats
To evaluate whether SP supplementation affects the mRNA expression of 5α-reductase, 17β-HSD3, and 3β-HSD in the testis of old male rats, we measured transcript levels using RT-PCR. We observed that SP supplementation resulted in a significant decrease in the mRNA expression of 5α-reductase and significant increases in the mRNA expression levels of 17β-HSD3 and 3β-HSD2 compared with the corresponding levels in the NC group (P < .05; Fig. 4A–C). These data indicate that SP supplementation stimulates testosterone biosynthesis and suppresses testosterone degradation in the testis of old rats.
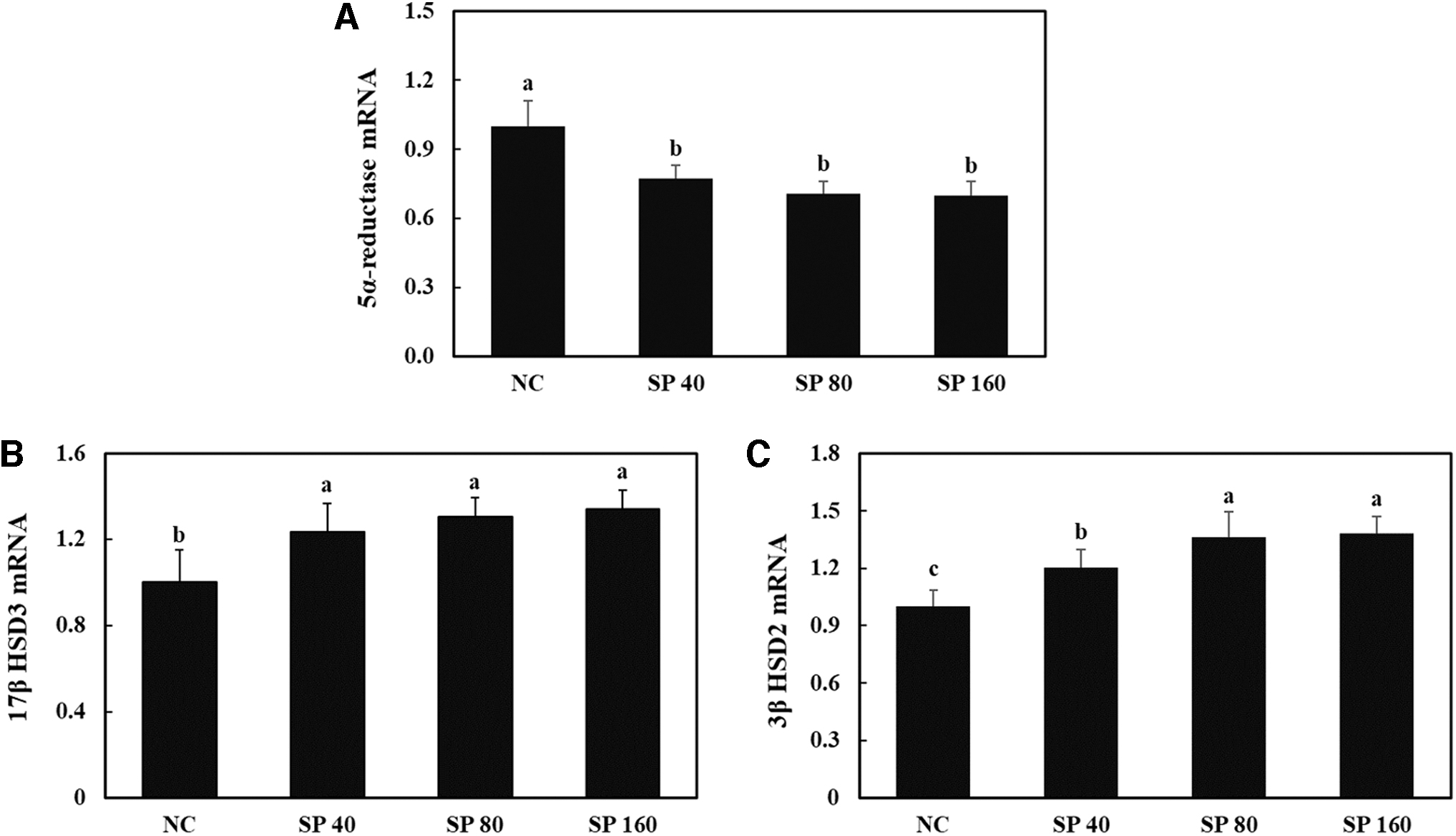
mRNA expression levels of 5α-reductase
Effects of SP on H2O2-induced oxidative stress in Leydig cells
We investigated whether SP treatment directly affects the production of testosterone using Leydig cells under H2O2-induced oxidative stress. H2O2-induced oxidative stress decreased the level of total testosterone and increased the level of 5α-reductase. However, treatment of these cells under H2O2-induced oxidative stress with SP increased total testosterone and decreased 5α-reductase compared with the corresponding levels in the control group (P < .05; Fig. 5A, B). In addition, we found that the mRNA expression levels of 5α-reductase and aromatase were decreased and the mRNA expression levels of 17,20-desmolase and 3β-HSD4 were increased in the SP-treated Leydig cells under H2O2-induced oxidative stress when compared with the corresponding levels in the control group (P < .05; Fig. 5C–F). Therefore, we suggest that supplementation with SP can increase circulating testosterone levels by directly promoting the production of testosterone in Leydig cells.
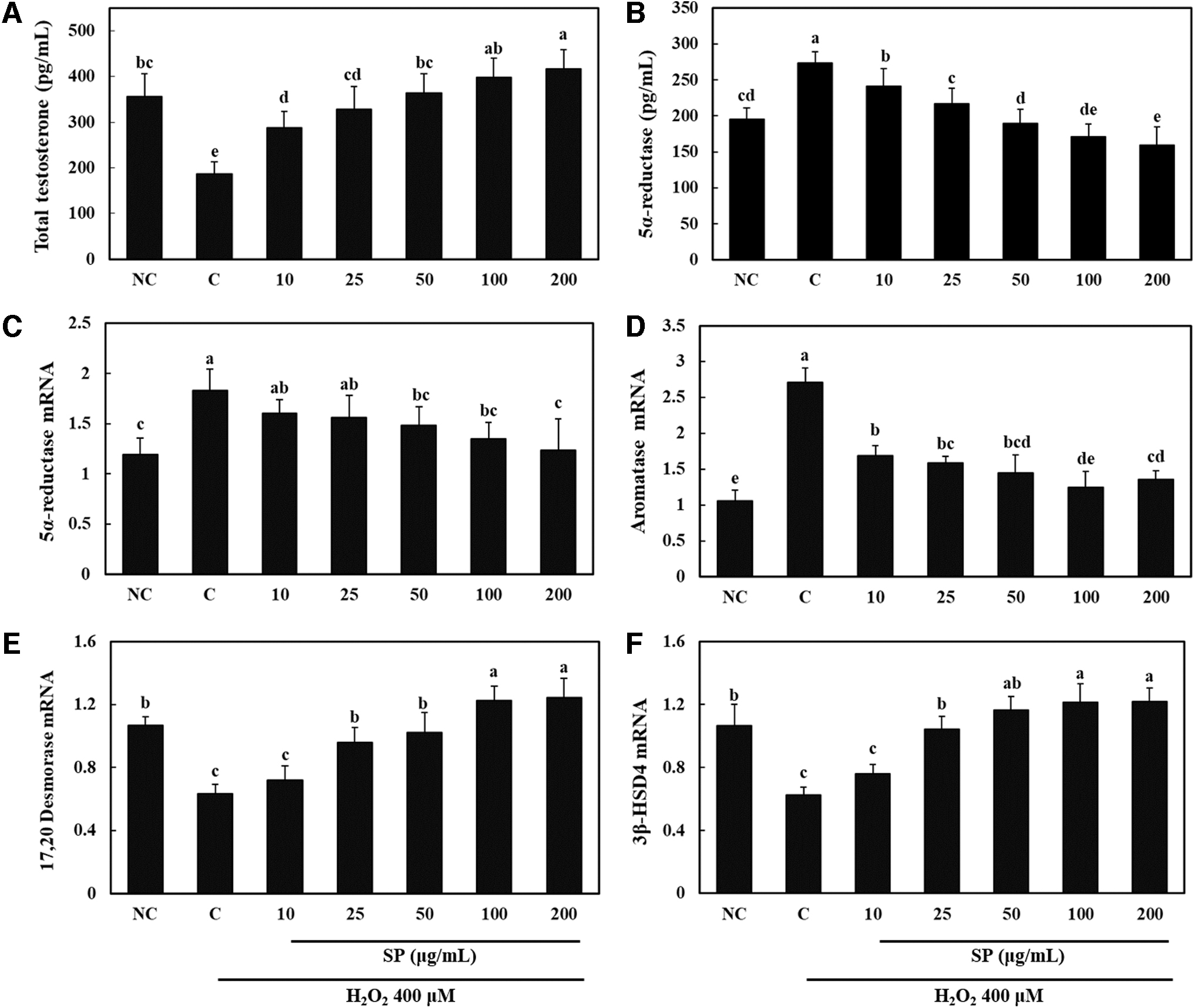
Total testosterone
Discussion
In this study, we investigated whether a standardized SP affects testosterone biosynthesis and spermatogenesis in old rats and H2O2-induced oxidative stress in Leydig cells to assess its potential andropause symptom-relieving effects. Phillips et al. demonstrated that testosterone deficiency contributes to an increased risk of coronary artery disease, 3 and McInnes et al. demonstrated that testosterone deficiency contributes to the development of obesity by increasing visceral fat. 4 Therefore, testosterone plays important roles not only in fertility via spermatogenesis but also in metabolic regulation. We found that supplementation of SP in old rats reduced body weight gain by decreasing visceral and epididymal fat weight without changing the weights of other tissues. In addition, SP supplementation induced decreases in the levels of serum TG, TC, and LDL/VLDL cholesterol and an increase in muscle endurance by reversing the testosterone deficiency. These results indicate that SP supplementation may prevent the testosterone deficiency-induced decrease in muscle endurance and the symptoms of metabolic syndrome, including the accumulation of adipose tissue and dyslipidemia.
Male fertility and maintenance of spermatogenesis are dependent on testosterone levels in the testis. Testosterone contributes to spermatogenesis in Sertoli cells via two pathways, referred to as the classical and nonclassical pathways, by maintaining the blood–testis barrier and promoting meiosis, Sertoli–spermatid adhesion, and sperm release. 7,18 In this study, we showed that SP supplementation increased the total sperm and active sperm counts in old rats. To determine whether the effect of SP on spermatogenesis is related to testosterone biosynthesis, we measured the levels of testosterone and the mRNA expression levels of enzymes and hormones involved in testosterone biosynthesis. We found that circulating levels of total and free testosterone were increased and SHBG and 5α-reductase mRNA expression levels were decreased by SP supplementation. SHBG is produced in the liver, and it binds to a portion of the total testosterone (40–60%) and inhibits its function. Free testosterone, which is not bound to SHBG, is biologically active. 19 Thus, SP supplementation increases not only total testosterone by promoting its biosynthesis but also bioavailable testosterone by reducing its binding to SHBG. In addition, SP supplementation increased the levels of circulating GH, LH, and FSH as well as the mRNA expression levels of 17β-HSD3 and 3β-HSD in the testis. Thus, these results indicate that SP supplementation increases the testosterone level indirectly through hormonal regulation by increased LH and FSH and inhibition of binding to SHBG. We suggest that supplementation of SP may relieve the symptoms of andropause syndrome, including decreased spermatogenesis and muscle endurance and metabolic syndrome by increasing testosterone biosynthesis and bioavailability.
Testosterone biosynthesis occurs in the Leydig cells of the testis and is subjected to hormonal regulation. LH stimulates testosterone biosynthesis by binding to the LH receptor on the Leydig cell membrane. However, aging-induced oxidative stress caused by reactive oxygen species inhibits testosterone biosynthesis by damaging Leydig cells. 20,21 In a previous study, we investigated whether treatment with SP can directly stimulate testosterone biosynthesis in a model of H2O2-induced oxidative stress in Leydig cells. We found that the mRNA expression levels of enzymes involved in testosterone biosynthesis were increased, whereas the mRNA expression levels of enzymes involved in testosterone degradation were decreased in Leydig cells under H2O2-induced oxidative stress when treated with SP. These data indicate that SP relieves andropause symptoms via testosterone biosynthesis, both directly by acting on damaged Leydig cells and indirectly through hormonal regulation.
In conclusion, we confirmed that a standardized SP relieves andropause symptoms, including decreased muscle endurance and spermatogenesis and metabolic syndrome, by increasing testosterone levels in a rat model of natural age-induced andropause. In addition, SP treatment directly induced testosterone biosynthesis in Leydig cells under H2O2-induced oxidative stress. Therefore, the results of this study suggest that supplementing with SP may be useful for alleviating andropause symptoms through direct and indirect regulation of testosterone biosynthesis.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.