
Abstract
Even though nasopharyngeal carcinoma (NPC) is not common worldwide, it is a major public health burden in endemic areas. Distant metastasis often leads to a poor prognosis for NPC; therefore, new and effective anticancer strategies are needed. Ginkgolic acid (GA) is small-molecule compound existing in Ginkgo biloba that has various biologically relevant activities, including antitumor properties; however, its effects and mechanism of action in NPC are unknown. The effects of GA on NPC and such underlying mechanisms were investigated using 5–8F and CNE2 cells and NP69 human immortalized nasopharyngeal epithelial cells in this study. Moreover, the xenograft models were built to examine GA's effection in vivo. GA treatment decreased the survival and invasive capacity of 5–8F and CNE2 and induced their apoptosis, which varied with dose; this was accompanied by downregulation of B cell lymphoma (Bcl)2, upregulation of Bcl2-associated X protein, and activation of poly-ADP ribose polymerase, and caspase-9/-3. G0/G1 phase arrest was induced by GA in NPCs. It also reduced the expression of cyclin-dependent kinase 6 and its regulators cyclin D2 and cyclin D3. GA inhibited the activation of protein kinase B/nuclear factor signaling; this effect was potentiated with GA and 5-fluorouracil (5-FU), which also enhanced 5-FU-induced apoptosis. In summary, GA may be effective as an adjuvant to conventional chemotherapy drugs in preventing the progression of NPC.
Introduction
Nasopharyngeal carcinoma (NPC) originates from nasopharyngeal epithelial cells. It is a highly aggressive and metastatic malignancy. 1 –3 Approximately 129,000 new NPC cases occurred in 2018, accounting for 0.7% of cancers diagnosed worldwide that year. 4 Although the global incidence of NPC is low, East and Southeast Asia have more than 70% of new cases. 4 Geography, environment, and genetics, coupled with Epstein/Barr virus infection, are all related to the occurrence of NPC. 5 Although the overall incidence and mortality of NPC are declining as a result of improvements in dietary habits, disease screening, and diagnosis and treatment methods. 6 However, in China, the incidence and mortality of NPC remain high, with a distant metastasis rate as high as 14.1%. 7
Unlike early-stage NPC that is treated with radiotherapy mainly, locally advanced disease is managed through a combination of chemotherapy and radiotherapy. Concurrent radiotherapy and chemotherapy combined with adjuvant chemotherapy or induction chemotherapy were recommended by the National Comprehensive Cancer Network (NCCN) guidelines for NPC treatment, which is supported by level 2A evidence. 8 The choice of chemotherapy drug affects the prognosis of NPC patients. Although the traditional agents recommended by the 2018 NCCN guidelines (e.g., cisplatin, carboplatin, 5-fluorouracil [5-FU], and docetaxel) are used as auxiliary or induction chemotherapy, 9 they are poorly tolerated and have low efficacy. The development of chemotherapy and radiotherapy resistance has also become one of the main factors for the poor treatment of NPC patients. As such, the development of safer and more effective treatment options for NPC is essential. Considering the importance of chemotherapy for NPC treatment, new drugs and approaches of combined chemotherapy are very important.
As an ancient gymnosperm in China, Ginkgo biloba L. is widely distributed worldwide nowadays. The plant extract has long been used in Traditional Chinese Medicine for its photoprotective, anti-inflammatory, and hepatoprotective effects along with its cardiotonic properties in the prevention and treatment of cardiovascular illness. 10 –12 G. biloba is commonly found in Traditional Chinese herbal Medicine. At present, mature G. biloba extract (G. biloba P.E, GBE) has been used in clinic for the treatment of cognitive decline and Alzheimer's disease. Many pharmacological studies have shown that terpene trilactone and flavonoids are the effective ingredients for GBE to play a pharmacological role in the treatment of cognitive impairment. 13,14 Ginkgolic acid (GA) is a primary bioactive compound GBE. At this time, the medicinal value of GA has not been explored, and it is recognized as a harmful substance. GA is a phenolic acid present in the GBE (Fig. 1A) with many biological activities. Its therapeutic potential has been investigated in relation to gastric, pancreatic, lung, colon, and breast cancers and oral squamous cell carcinoma. The antitumor effects of GA include inhibiting tumor growth and proliferation, invasion and metastasis; blocking cell cycle progression; and inducing cell apoptosis. However, whether these effects apply to NPC—as well as the underlying mechanisms—remains unclear.
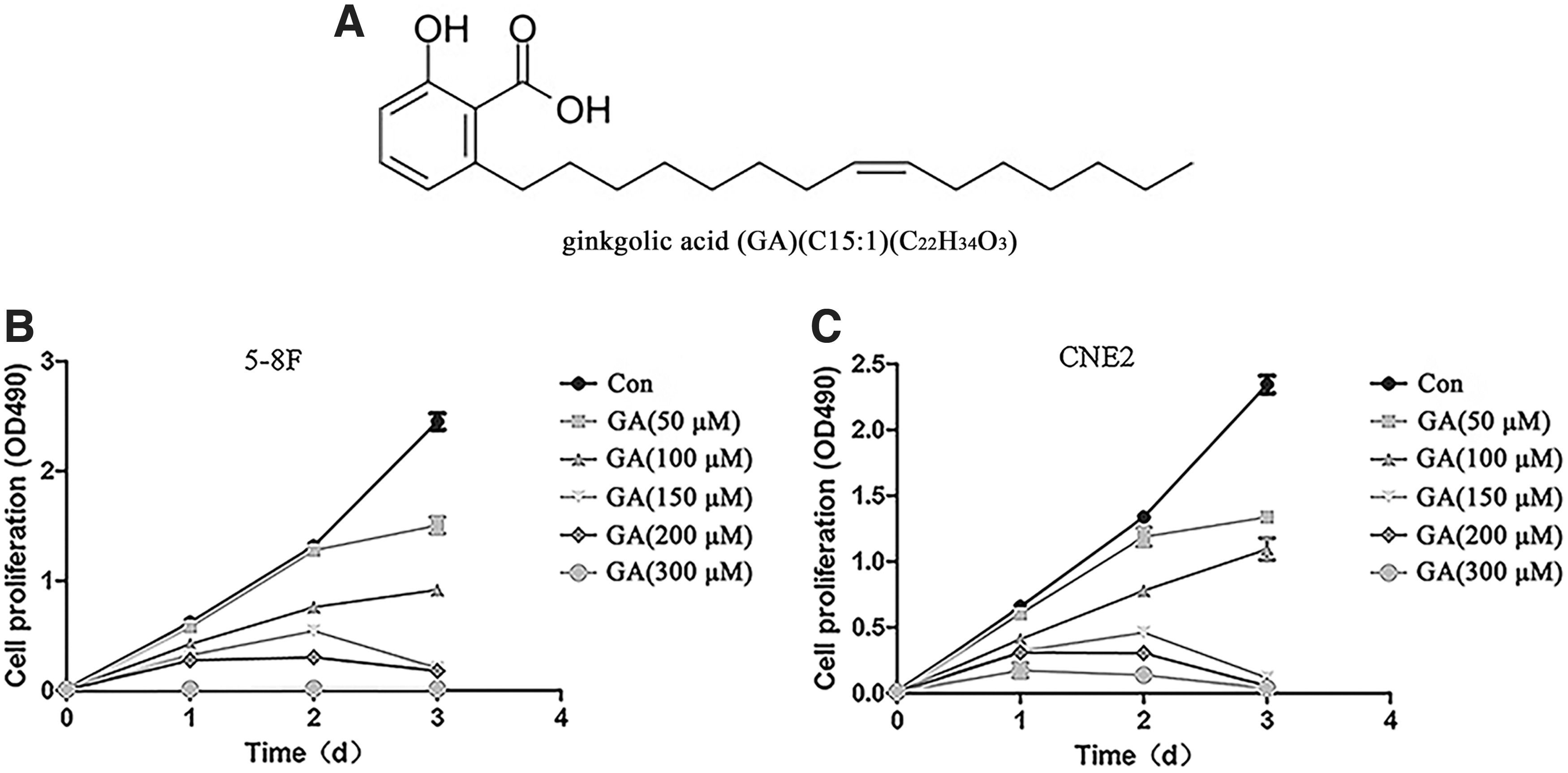
GA inhibits NPC cell proliferation.
Inducing tumor cell apoptosis is a commonly used therapeutic strategy for blocking tumor progression. 15,16 The phosphoinositide 3-kinase (PI3K)/protein kinase B (AKT) axis is most frequently altered in human cancers. Single-nucleotide polymorphisms in the gene encoding AKT were found in a phase II clinical trial of patients with head and neck squamous cell carcinoma (HNSCC), which are associated with poor prognosis. 17 Phosphorylated (p-)AKT (Ser473) can be used to predict poor radiotherapy outcomes of advanced HNSCC patients. 18 On the one hand, nuclear factor (NF-κB) can regulate the expression of inflammation-related genes. On the other hand, NF-κB also has an important influence on cell cycle control, cell proliferation, apoptosis, and other biological behaviors. 19 –21 In addition, increased activation of NF-κB can be directly related to tumor cell resistance to ionizing radiation and chemotherapy. 22,23 It is also reported in NPC.
This study mainly investigated the effects of GA on NPC progression as well as its internal mechanism. GA inhibited the proliferation and invasion of NPC cells. It also induced their apoptosis by activating the caspase signaling. In addition, GA acted synergistically with 5-FU not only in NPC cells through suppression of AKT/NF-κB activation but also in vivo. Our results provide evidence that GA has an inhibitory effect on NPC.
Materials and Methods
Cells and culture
The 5–8F,CNE2, and NP69 are maintained in our laboratory, which were cultured in Glasgow's minimum essential medium (Gino Hangzhou Biomedical Technology Co., Hangzhou, China). Ten percent fetal calf serum, penicillin (100 U/mL), and streptomycin (100 μg/mL) were added. GA (C15:1) and 5-FU were purchased from MedChemExpress (Monmouth Junction, NJ, USA) and Beijing Zhongshuo Pharmaceutical Technology Development Corporation, respectively.
Cell viability analysis
In 96-well plates, cells were treated with different concentrations of GA. Cell Counting Kit (CCK)-8 solution was added in corresponding wells and then incubated for 2 h at 37°C. The absorbance at 490 nm was measured with a microplate analyzer (PerkinElmer, Waltham, MA, USA).
Wound-healing assay
5–8F and CNE2, which were grown in six-well plates, were serum-starved overnight before pretreatment with GA. A 1-mL sterile pipette tip was used to scribe line on the cell monolayer. Then the plates were washed once with phosphate-buffered saline (PBS). Serum-free medium was added and images of the same fields were obtained at three time points (0, 24, and 48 h) with an inverted microscope at 40 × magnification (BX51; Olympus, Tokyo, Japan).
Cell invasion assay
Transwell inserts (Corning Inc., Corning, NY, USA) with Matrigel were used for the cell invasion assay. Treated with GA (0, 50, and 100 μM) for 24 h, 5–8F and CNE2 cells were seeded onto the inserts. After being immobilized with 4% paraformaldehyde solution (Wuhan Aspen Biotechnology Co., Wuhan, China) and stained with crystal violet (Wuhan Seville Biological Company, Wuhan, China), transwell inserts were observed by a digital camera (Nikon, Tokyo, Japan) at 200 × magnification. ImageJ v1.48 software was used to determine the number of cells that had penetrated the insert.
Flow cytometry analysis
The fluorescein isothiocyanate (FITC)/Propidium Iodide (PI) apoptosis detection kit (Lianke Biotech Co., Hangzhou, China) was used to detect apoptotic cells. Briefly, 1 × 105 cells were resuspended in 500 μL binding buffer. Then, 5 μL Annexin V-FITC and 10 μL PI were added, followed by incubation in the dark for 20 min. The apoptotic fraction was sorted with a FACSCalibur flow cytometer (BD Biosciences, NJ, USA). The experiment was repeated three times.
Cell cycle arrest was examined by using the PI/RNase Staining Buffer kit (BD Biosciences). Cells were washed with precooled PBS, and then fixed with a fixation buffer at 4°C for 25 min. A 5 mL volume of 75% cold ethanol was added and the samples were placed at −20°C for at least 2 h. After washing with PBS, the cells were resuspended in 500 μL staining buffer at room temperature and sorted within 1 h.
Western blotting analysis
Cell total protein was extracted by radio immunoprecipitation assay cell lysis buffer (Beyotime Biotechnology, Shanghai, China) containing a protease and phosphatase inhibitor mixture (Roche, Indianapolis, IN, USA). After centrifugation at high speed and low temperature for 20 min, the protein concentration was measured and the protein was denatured. The denatured protein was separated by sodium dodecyl sulfate-polyacrylamide gel electrophoresis and transferred to a polyvinylidene fluoride membrane (EMD Millipore, MA, USA). After blocking with 5% skimmed milk powder and adding a primary antibody, it was incubated overnight at 4°C. The next day, it was incubated with horseradish peroxidase-goat anti-rabbit/mouse IgG (1:3000; Thermo Scientific, MA, USA). The protein was detected with an enhanced chemiluminescent substrate (ChemiDoc MP; Bio-Rad, CA, USA). The primary antibody was purchased from Cell Signaling Technology, and ImageJ software was used to analyze protein bands.
In vivo studies
Five-week-old male BALB/c-nu mice were purchased from Vital River Laboratory Animal Technology (Beijing, China). The experiments were approved by the Institutional Animal Care and Use committee at Wuhan University (Protocol No. 20201116). After 1 week of acclimation, these nude mice were implanted subcutaneously into the right armpit with 1 × 106 5–8F cells in 0.1 mL culture medium to obtain NPC xenografts. The mice were then divided into five groups randomly (5/group): PBS, Solvent group (10% dimethyl sulfoxide +90% corn oil), 5 mg/kg 5-Fu, 15 mg/kg GA, and 15 mg/kg GA +5 mg/kg 5-Fu. Mice were administered with drugs via gastric gavage (3 times/week) for 3 weeks. The equation V = a × b 2 /2 (mm3) was used to calculate the volume.
Statistical analysis
Mean ± standard error of the mean was used to present data and Prism v5.0 Software (GraphPad, La Jolla, CA, USA) was used for statistical analysis. Differences between groups were evaluated by two-way analysis of variance (ANOVA) test or one-way ANOVA test. The t-test of two independent samples was used to compare the mean between two groups of measurement data. A P < .05 was considered statistically significant.
Results
GA inhibits the proliferation, migration, and invasion of NPC cells
GA's effect on NPC growth was investigated by CCK-8. Results demonstrated that GA reduced the viability of 5–8F and CNE2 cells in a time- and dose-dependent manner (Fig. 1B, C). In the wound-healing assay, we found that GA also inhibited their migration (Fig. 2A–C). The number of cells passing through the insert was dose dependently reduced in the presence of GA by the transwell invasion assay (Fig. 2D, E). These results demonstrate that GA inhibits the proliferation, migration, and invasion of NPC cells.

GA inhibits the migration and invasion of 5–8F and CNE2 human gastric cancer cells.
GA induces cell cycle arrest by inhibiting cyclin-dependent kinase 6 expression
Flow cytometry analysis revealed that GA treatment increased the percentage of 5–8F and CNE2 cells in G0/G1 phase dose dependently, while reducing the G2/M and S fractions (Fig. 3A, B). To determine the mechanism of cell cycle arrest, we examined cell cycle-associated proteins by Western blotting. The levels of cyclin-dependent kinase (CDK)6 and its regulatory factors cyclin D2 and cyclin D3 were downregulated by GA treatment (Fig. 3C). Thus, GA inhibits NPC growth by inducing cell cycle arrest in tumor cells.

GA induces cell cycle arrest in human gastric cancer cells by regulating CDK6 expression.
GA induces apoptosis in NPC cells
Apoptosis is targeted by many antitumor drugs. 24 Flow cytometry analysis showed that GA enhanced apoptosis in 5–8F and CNE2 cells in a dose-dependent manner (Fig. 4A, B). However, there was no obvious proapoptotic effect in NP69 nasopharyngeal epithelial cells. Western blotting revealed that the expression of the antiapoptotic protein B cell lymphoma (Bcl)2 was downregulated, whereas that of the proapoptotic factor Bcl2-associated X protein (Bax) was upregulated by GA treatment (Fig. 4C). In addition, cleaved poly-ADP ribose polymerase (PARP) and caspase-3/-9 protein expression were increased by GA in contrast with untreated control cells in 5–8F and CNE2 (Fig. 4C). Thus, caspase-dependent apoptosis was induced by GA in NPC cells.

Apoptosis of human gastric cancer cells induced by GA treatment.
Antitumor effects of GA in NPC are mediated via inhibition of AKT/NF-κB signaling
The AKT/NF-κB signaling has been shown to modulate tumor progression by regulating apoptosis. 25 Given that NF-κB positively regulates survival genes in cancer cells 26,27 and is also a trigger for cancer cell resistance to treatment, 28 we speculated that GA may exert antitumor effects in NPC by suppressing NF-κB signaling. To investigate such a possibility, we evaluated the protein expression of p-AKT, AKT, NF-κB p65, IκBα, p-IκBα, and NF-κB p-p65 in GA-treated cells by Western blotting. Those groups that were treated with 50 and 100 μM GA showed reduced levels of IκBα, p-IκBα, p-AKT, and NF-κB p-p65 (Fig. 5). These data indicate that GA inhibits the activation of the AKT/NF-κB pathway.

GA inhibits the activation of the AKT/NF-κB signaling pathway in human gastric cancer cells.
GA acts synergistically with 5-FU to induce apoptosis of NPC cells
5-FU is a chemotherapy drug that is frequently used to treat NPC. We investigated whether 5-FU can be used as an adjuvant chemotherapy for NPC by treating 5-8F and CNE2 cells with different concentration of GA (50 and 100 μM) and 5-FU. GA dose dependently enhanced the effects of 5-FU chemotherapy on NPC cell proliferation and apoptosis (Fig. 6A, B). The antiproliferative and proapoptotic effects of GA involved suppression of AKT/NF-κB signaling (Fig. 6C, D). GA treatment also increased the 5-FU-mediated inhibition of this pathway.
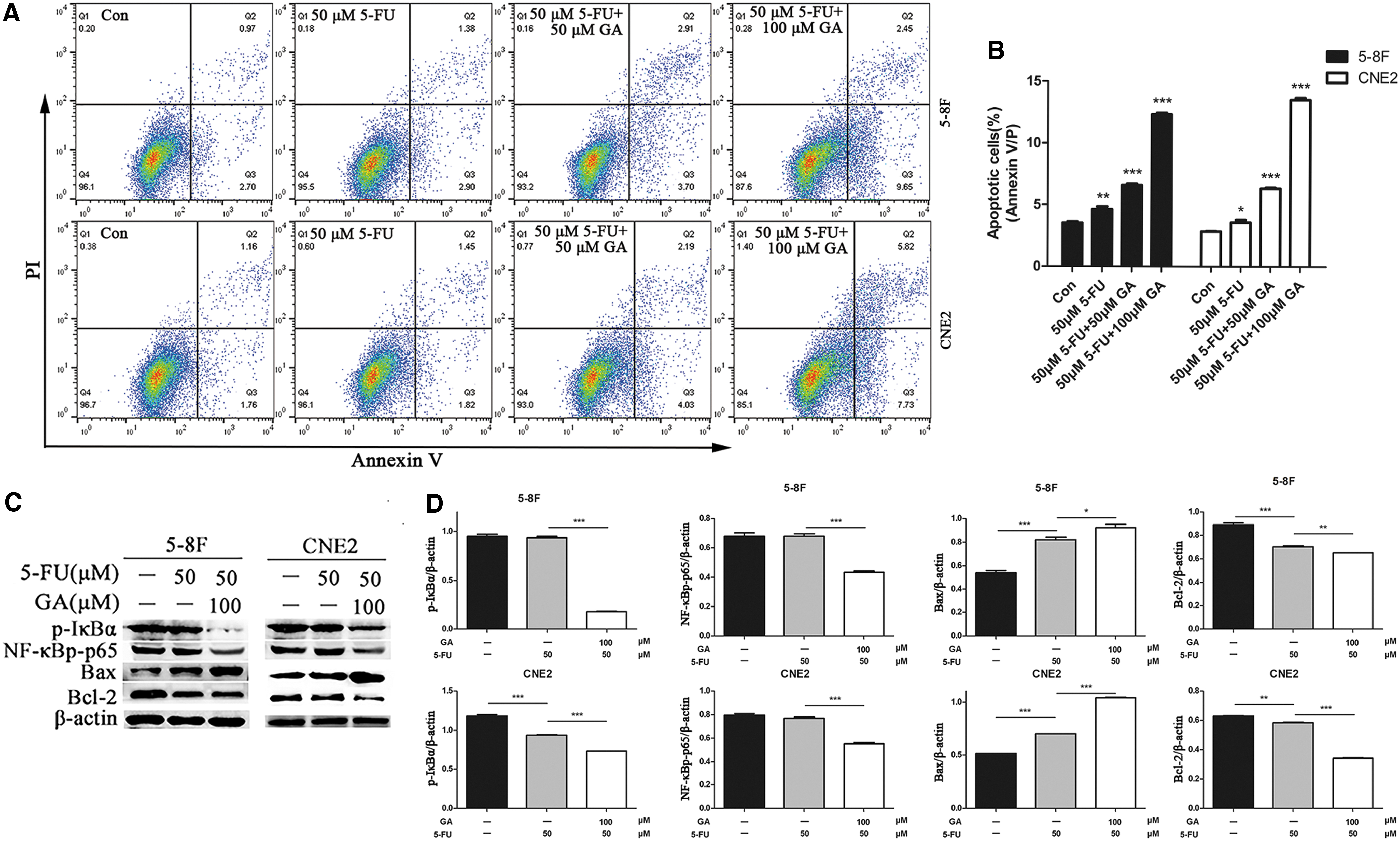
GA and 5-FU act synergistically to induce apoptosis of 5–8F and CNE2 cells.
Synergistic inhibitory effect of GA and 5-FU on the growth of NPC
We established xenograft models using 5–8F in this part. The results were as follows (Fig. 7): there was no difference between the normal saline control group and the solvent group, while the xenograft volume and weight in the GA-treated group were significantly smaller compared with the solvent group. These results showed the inhibitory capability of GA on the development and growth of NPC. The size of transplanted tumors in the 5-FU group also decreased significantly, which was more obvious than in those treated with GA alone. The inhibitory effect was further improved with combined GA and 5-FU, which indicated that GA and 5-FU had synergistic effects in the treatment of NPC. GA enhances the sensitivity of NPC cells to 5-FU treatment
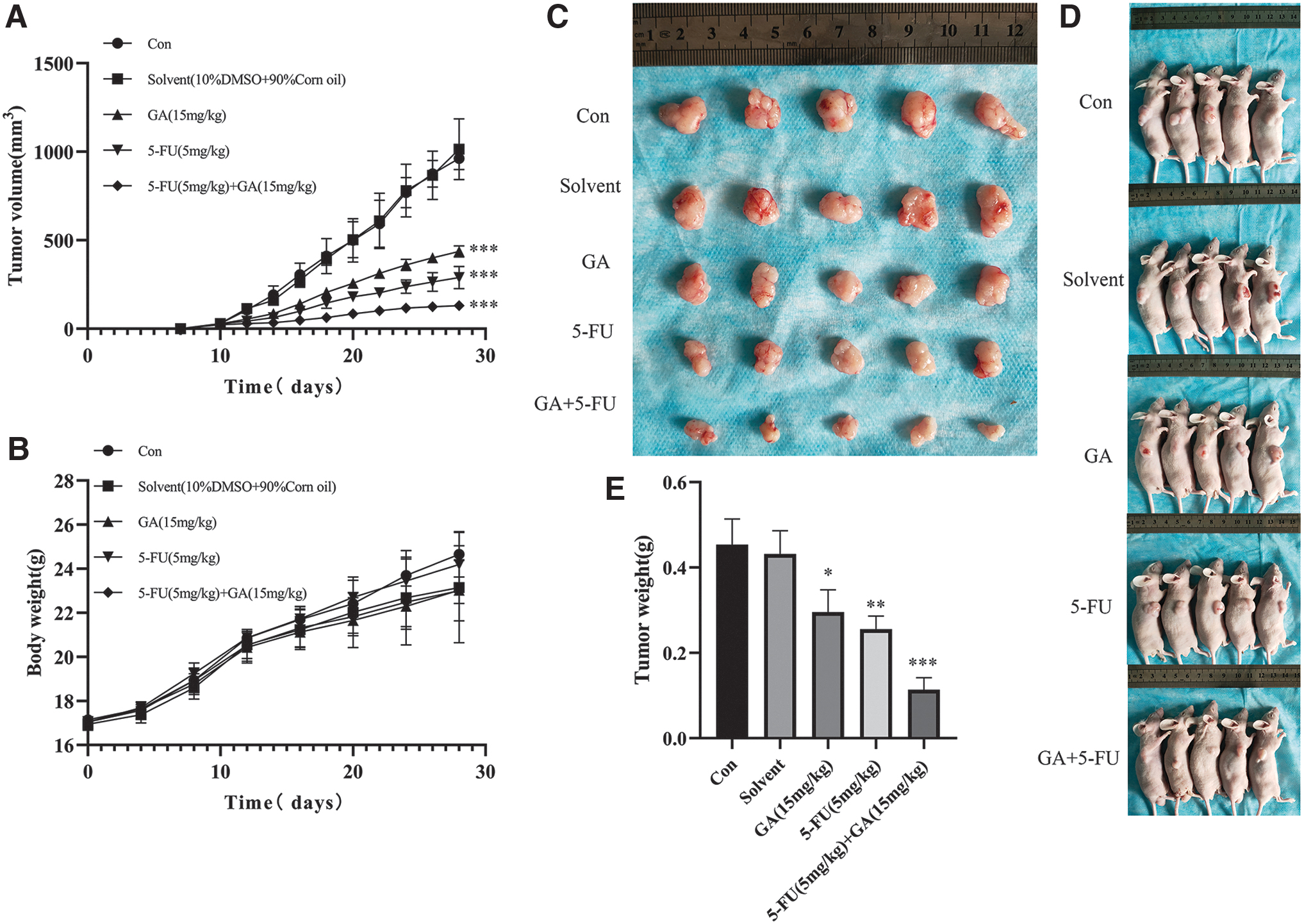
GA inhibited the growth of NPC transplanted tumor and had a synergistic effect with 5-FU. The 5–8F cells were xenografted onto BALB/c nude mice flank.
Discussion
NPC is a highly aggressive head and neck tumor that develops resistance to radiotherapy and chemotherapy, leading to distant metastasis and poor prognosis. 3,29 New therapeutic strategies are needed to improve clinical outcomes. GA, a compound extracted from G. biloba leaves, has a variety of biologically relevant properties, including antitumor activity. 30 –33 This study proved for the first time that GA can block the progression of NPC through inhibiting cell proliferation, invasion, and improving apoptosis of tumor cells. These effects were mediated, in part, via suppression of AKT/NF-κB signaling. Moreover, GA enhanced the antitumor effects of 5-FU, including inducing apoptosis and negatively regulating the AKT/NF-κB pathway. In addition, we have demonstrated the inhibiting influence of GA on the growth of NPC and its synergistic therapeutic effect with 5-FU in vivo.
Cancer is characterized by aberrant cell cycling. Cyclins are often overactivated, which leads to uncontrolled cell proliferation in cancer cells. Many anticancer drugs are synthetic inhibitors of CDKs. 34 In our study, we found that GA induced G0/G1 arrest in NPC cells, thereby preventing G1/S-phase transition and inhibiting cell proliferation. Cyclin D, CDK4, and CDK6 have oncogenic functions, 35 –38 and the CDK4/CDK6 inhibitor palbociclib has been approved for breast cancer treatment. Here we demonstrated that GA dose dependently decreased the expression of CDK6, cyclin D2, and cyclin D3 in NPC cells, confirming its antitumor activity. The development of highly selective and potent compounds is the key to the success of cell cycle targeted cancer therapy. Our results proved that GA has the potential for NPC treatment that acts by targeting the cell cycle.
Apoptosis is a type of programmed cell death characterized by chromatin condensation, nucleus and DNA fragmentation, cell membrane blebbing, and cell shrinkage. 39,40 Most of the current cancer treatments function by inducing cancer cell apoptosis. 41 Thus, small-molecule compounds targeting apoptosis pathways are among the therapeutic options for cancer treatment. Apoptosis signaling comprises intrinsic and extrinsic pathways. 42 In the former, Bcl2 family proteins can regulate the permeabilization of the outer mitochondrial membrane. Bcl2 inhibits caspase activity by preventing Bax activation. As the main initiator of this pathway, it activated the effector caspase-3 and cleaved caspase-9, 43,44 inducing cell death. 45 The antiapoptotic activity of GA was shown to involve decreases in the Bcl2/Bax ratio and expression of Bcl2 and Bcl extralarge (Bcl-xl) in gastric cancer cells, as well as upregulation of the antiapoptotic factors Bax and Bcl2-associated death promoter (Bad), which enhanced the cleavage of caspase3 and caspase9 and increased the expression of PARP, resulting in cell death. 46 In our study, GA treatment enhanced apoptosis in 5–8F and CNE2 compared with NP69 and increased the levels of Bax, caspase-9/-3, and PARP, while decreasing that of Bcl2. These results indicate that GA promotes apoptosis in NPC cells via a caspase-dependent mechanism. However, additional studies are required to identify whether GA also targets the extrinsic apoptosis pathway in the future.
NF-κB transcription factors are involved in many aspects of tumorigenesis. Stimulating proliferation, inhibiting apoptosis, and improving angiogenesis are included. 27 As an inhibitor of programmed cell death, NF-κB can reduce the apoptotic response to ionizing radiation and genotoxic anticancer drugs. 47,48 AKT and NF-κB are constitutively active in HNSCC. 49,50 In the present work, GA blocked the activation of the AKT/NF-κB pathway in 5–8F and CNE2; moreover, it enhanced the inhibitory functions of 5-FU on NPC cell apoptosis and AKT/NF-κB signaling. Thus, GA can be used as an adjunct to traditional chemotherapy drugs to prevent the progression of NPC.
This study had some limitations. First, GA may be potentially toxic and its pharmacokinetic data in vivo need to be further supplemented. Second, while we focused on the Akt/NF-κB axis, it is possible that GA exerts antitumor effects via modulation of other signaling pathways. Nonetheless, our results demonstrate that GA has a therapeutic potential for NPC. GA acts by suppressing the proliferation, migration, and invasion of NPC cells both alone and in conjunction with 5-FU, which broadens the therapeutic options for this relatively rare but highly malignant neoplasm.
Footnotes
Acknowledgment
The authors would like to thank the Central Laboratory of Renmin Hospital of Wuhan University for experiment resources.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.