
Abstract
Immunosuppression occurs in response to a variety of external antigens. However, various immune cells and cytokines can activate the immune system. In this study, it was found that fermented deer velvet (FD) and fermented Eleutherococcus senticosus (FE) extract (FDE) mixtures regulated the immunity of animals that underwent induced immunosuppression through forced swimming exercise (FSE). Seven mouse treatment groups were included in the experiment: normal controls, FSE controls, positive controls (FSE+red ginseng 300 mg/kg body weight), FD200 (FSE+FD 200 mg/kg body weight), FE200 (FSE+FE 200 mg/kg body weight), FDE50 (FSE+FDE 50 mg/kg body weight), and FDE200 (FSE+FDE 200 mg/kg body weight). Oral intake of experimental and control substances lasted for 2 weeks. Oral FDE intake increased cell counts for major histocompatibility complex (MHC) I, MHC II, CD4(+) T cells, and CD8(+) T cells compared with controls. Moreover, FDE increased Th1 (interleukin [IL]-2 and interferon gamma) cytokine proliferation, T cell proliferation, IL-12 and IL-15 production, and natural killer cell activity compared with controls. In addition, FDE inhibited Th2 cytokines (IL-4, IL-6, IL-10, and tumor necrosis factor alpha) and nitric oxide production, increased B cell proliferation and leukocyte count, and promoted immunoglobulin A and G serum levels compared with controls. Thus, the finding that FDE increased immune function in an immunosuppression model suggests that FDE has immunomodulatory capacity.
Introduction
People living in industrialized societies show a strong willingness to invest in health improvement, regardless of economic conditions, as evidenced by increasing sales of dietary supplements to boost immunity and counteract decreased immunity because of air pollution, and other concerns. 1,2 When external antigens or stimuli, such as bacteria and viruses, enter the body, antigen-presenting cells (APCs) recognize them and immediately activate the immune defense system. 3
Viruses typically enter APCs through major histocompatibility complex (MHC) I molecules, and bacteria or other toxins enter APCs through MHC II. MHC I binds to the T cell receptor of CD8(+) T cytotoxic cells and MHC II binds to CD4(+) T helper cells. 4 CD4(+) T helper cells release T helper-1 (Th1)-type cytokines, such as interleukin (IL)-2 and interferon gamma (IFN-γ), as well as T helper-2 (Th2)-type cytokines, such as IL-4, IL-6, IL-10, and tumor necrosis factor alpha ( (TNF-α). An imbalance in Th1-type cytokines and Th2-type cytokines can negatively affect the immune system by causing immunosuppression. 5,6 This decreases T cell and B cell function, and decreases cytotoxic T cells and natural killer (NK) cell activity, which are activated by Th1-type cytokines; inhibiting these immune cell activities interferes with antigen removal, promotes abnormal production of immunoglobulin (Ig), and alters leukocyte numbers. 7 –10
Factors beyond external antigens that can affect the immune system include aging, nutritional status, exercise, and mental stress, among which, exercise is closely associated with daily life and physical stress, thus, greatly contributing to a person's immune function. 11 Regular, individualized exercise regimens have positive effects on immunity, but irregular or one-time exercise can lead to decreased immunity. 12 The forced swimming exercise (FSE) used in this study was designed for muscle facilitation experiments and despair behavior experiments, among others. It has also recently been used to evaluate immune function enhancement. 13 Exercise was classified into three levels according to maximum oxygen intake, and swimming was classified as high-intensity exercise because it requires a maximum oxygen intake of 70–85 VO2 (%). 14 Thus, forced swimming is an appropriate exercise for an immunosuppressive model.
Deer velvet has been widely used as a therapeutic agent in Korea and China for 2000 years. Particularly, fermented deer velvet (FD) has been reported to have an osteoclast differentiation inhibitory effect, an immunostimulating effect, and anti-inflammatory activity. 15 –19 Eleutherococcus senticosus, also called Siberian ginseng, is used as sedative and tonic, and is effective for treating rheumatism and diabetes. 20 –23 However, little is known about the immunomodulatory capacity of fermented Eleutherococcus senticosus (FE), FD, and FE extract (FDE) mixtures.
In this study, we investigated the immunomodulation effects of FDE mixtures in C57BL/6N mice in which immunosuppression was induced by FSE.
Materials and Methods
Preparation of plant materials
FD and FE hot water extracts were obtained from Green Store, Inc., R&D Center (Seongnam, Korea). The FDE mixture ratio of FD to FE was 3:7, which was determined after confirming that this was the most effective ratio through in vitro screening experiments. The FD and FE extracts and the FDE mixtures were sealed from light and air and stored at −20°C until use.
Experimental animals and treatment
The animal protocols were approved by the Institutional Animal Care and Use Review Committee of Kyung Hee University (KHUASP-19-409). Male C57BL/6N mice (N = 56, 4-week-old mice, mean weight: 17.5 ± 0.3 g) were obtained from Saeronbio, Inc. (Uiwang, Korea). The animal care condition was maintained at 24°C ± 1°C, 55–60% relative humidity, and 12-h light/12-h dark cycles. The experimental animals were acclimated for 1 week and provided with standard pellet chow and water ad libitum.
Mice were divided into seven groups: normal control (NC), FSE control (exercise-C, C), positive control (PC; FSE+red ginseng 300 mg/kg body weight), 24 FD200 [FSE+FD 200 mg/kg body weight (FD 4 mg/20 g of mouse/day)], FE200 [FSE+FE 200 mg/kg body weight (FE 4 mg/20 g of mouse/day)], FDE50 [FSE+FDE 50 mg/kg body weight (FDE 1 mg/20 g of mouse/day)], and FDE200 [FSE+FDE 200 mg/kg body weight (FDE 4 mg/20 g of mouse/day)]. Oral intake of experimental and control substances lasted for 2 weeks. After killing, spleens and blood samples were separated for analyses.
Forced swimming exercise
The FSEs were conducted 3 days before (first swim, for 15 min), 1 day before (second swim, for 50 min), and on the day (third swim, for 2 h) the animals were killed. The swimming facility was an acrylic water tank (W 40 cm × L 25 cm × H 18 cm), and the experiment was carried out maintaining a flow rate of 14 L/min at a temperature of 25°C. Each mouse was forced to swim until it could no longer float in the water by itself, at which point, the exercise was ended. 13
Flow cytometry
Separated splenocytes were seeded at 3 × 106 cells/tube and were reacted with each of the following: anti-mouse CD8a FITC, anti-mouse CD4 PE, anti-mouse CD45-PE/CY7 (SouthernBiotech, Birmingham, AL, USA), MHC class I monoclonal antibody, and MHC class II monoclonal antibody (Thermo Fisher Scientific, Waltham, MA, USA), with light blocked for 30 min on ice. Next, the washing step was performed two times using a flow cytometry staining buffer (Thermo Fisher Scientific) and the supernatant was removed. Subsequently, the pellet was suspended in flow cytometry staining buffer and measured with CytoFLEX (Beckman Coulter, Brea, CA, USA). Results were analyzed using the CytExpert 2.2 program provided by Beckman Coulter.
T and B cell proliferation of splenocytes
Separated splenocytes were seeded at 5 × 105 cells/well in 96-well tissue culture dishes. Next, cells were treated with concanavalin A (Con A; 5 μg/mL; Sigma-Aldrich, St. Louis, MO, USA) or lipopolysaccharide (LPS; 5 μg/mL; Gibco-BRL; Grand Island, NY, USA) to promote T and B cell proliferation. After a 48-h incubation (37°C, 5% CO2), T and B cell proliferation of splenocytes was measured using EZ-CyTox (Daeil Lab Service, Seoul, Korea) following the manufacturer's protocol.
Enzyme-linked immunosorbent assay for cytokines in splenocytes
Separated splenocytes were seeded at 5 × 105 cells/well in 96-well tissue culture dishes. Next, the splenocytes were treated with Con A (5 μg/mL) to stimulate IL-2, -4, -10, -12, -15, and IFN-γ production. They were also treated with LPS (5 μg/mL) to stimulate IL-6 and TNF-α production. After a 24-h incubation (37°C, 5% CO2), supernatants were assessed for IL-2, -4, -6, -10, and TNF-α levels. IL-12 and IL-15 levels were measured after a 48-h incubation, and IFN-γ levels were measured after a 72-h incubation. R&D Duoset ELISA Development kits (R&D Systems, Minneapolis, MN, USA) were used to measure cytokine levels according to the manufacturer's protocol.
Determination of NK cell activity
Separated splenocytes were seeded at 5 × 105 cells/well in 96-well tissue culture dishes and were then cocultured with Yac-1 cells (ATCC, Rockville, MD, USA) at 1 × 104 cells/well. After a 4-h incubation (37°C, 5% CO2), supernatants were transferred to a new plate, and NK cell activity was measured using a CytoTox 96 Non-Radioactive Cytotoxicity Assay kit (Promega Corp., Madison, WI, USA) according to the manufacturer's protocol.
Blood sampling and analysis
Serum samples were collected from centrifuged (556 g at 4°C for 20 min) whole blood. Nitric oxide (NO), IgE, IgA, and IgG levels were measured using Mouse ELISA kits (Abcam, Massachusetts, United Kingdom) according to the manufacturer's protocols.
Leukocytes were isolated from whole blood that was collected through a syringe containing 2% EDTA (Sigma-Aldrich). Whole blood mixed with DPBS was floated slightly on Histopaque®-1077 (Sigma-Aldrich) and then was centrifuged for 20 min (556 g at 4°C). Next, the supernatant was removed and mixed with DPBS to count the number of leukocytes.
Statistical analyses
All results are given as mean ± standard deviation. We analyzed the significance of experimental results with Duncan's multiple range test after conducting a one-way analysis of variance using PASW Statistics version 22.0 (SPSS, Inc., Chicago, IL, USA). Results were considered statistically significant at P < .05.
Results
Effects of FDE on body weight and organ weight in an immunosuppression mouse model
Table 1 shows the effects of FDE on body and organ weight, food intake, and food efficiency rate (FER). There were no significant differences in weight gain, food intake, or FER among all groups. Spleen and liver weights in the exercise-C group were higher than those in the NC group, and scores were lower in the FDE groups compared with the exercise-C group. However, there were no significant differences in organ weights among all groups.
Effects of FDE on Weight Gain, Food Intake, FER**, and Organ Weight of C57BL/6N Mice with Induced Immunosuppression by Forced Swimming Exercise
Values are presented as mean ± SD (n = 8), and different superscript letters indicate significance at P < .05. NC, normal control; C, control, induced immunosuppression by FSE; PC, positive control (FSE+red ginseng 300 mg/kg body weight); FD200 (FSE+FD extract 200 mg/kg body weight); FE200 (FSE+FE extract 200 mg/kg body weight); FDE50 (FSE+FD and FE 3:7 50 mg/kg body weight); FDE200 (FSE+FD and FE 3:7 200 mg/kg body weight).
Weight gain (g/2 weeks) = final body weight (g) − initial body weight (g) **FER (FER) = weight gain (g)/total food consumption (g) × 100.
FD, fermented deer velvet; FDE, fermented deer velvet, and fermented Eleutherococcus senticosus extract; FE, fermented Eleutherococcus senticosus; FER, food efficiency rate; FSE, forced swimming exercise; SD, standard deviation.
Effects of FDE on the number of MHC I, MHC II, CD4(+), and CD8(+) T cells in primary splenocytes in an immunosuppression mouse model
The MHC I count was significantly depressed in the exercise-C group compared with the NC group, and was significantly elevated in the PC group compared with the exercise-C group. Compared with the exercise-C group, the FE200 and FDE200 groups showed significantly elevated MHC I counts, with the FDE groups showing a significant dose-dependent trend. MHC II counts were significantly depressed in the exercise-C group compared with the NC group and were significantly elevated in the PC group compared with the exercise-C group. Compared with the exercise-C group, the PC, FD200, and FDE groups had significantly elevated MHC II counts (P < .05) (Fig. 1A, C).
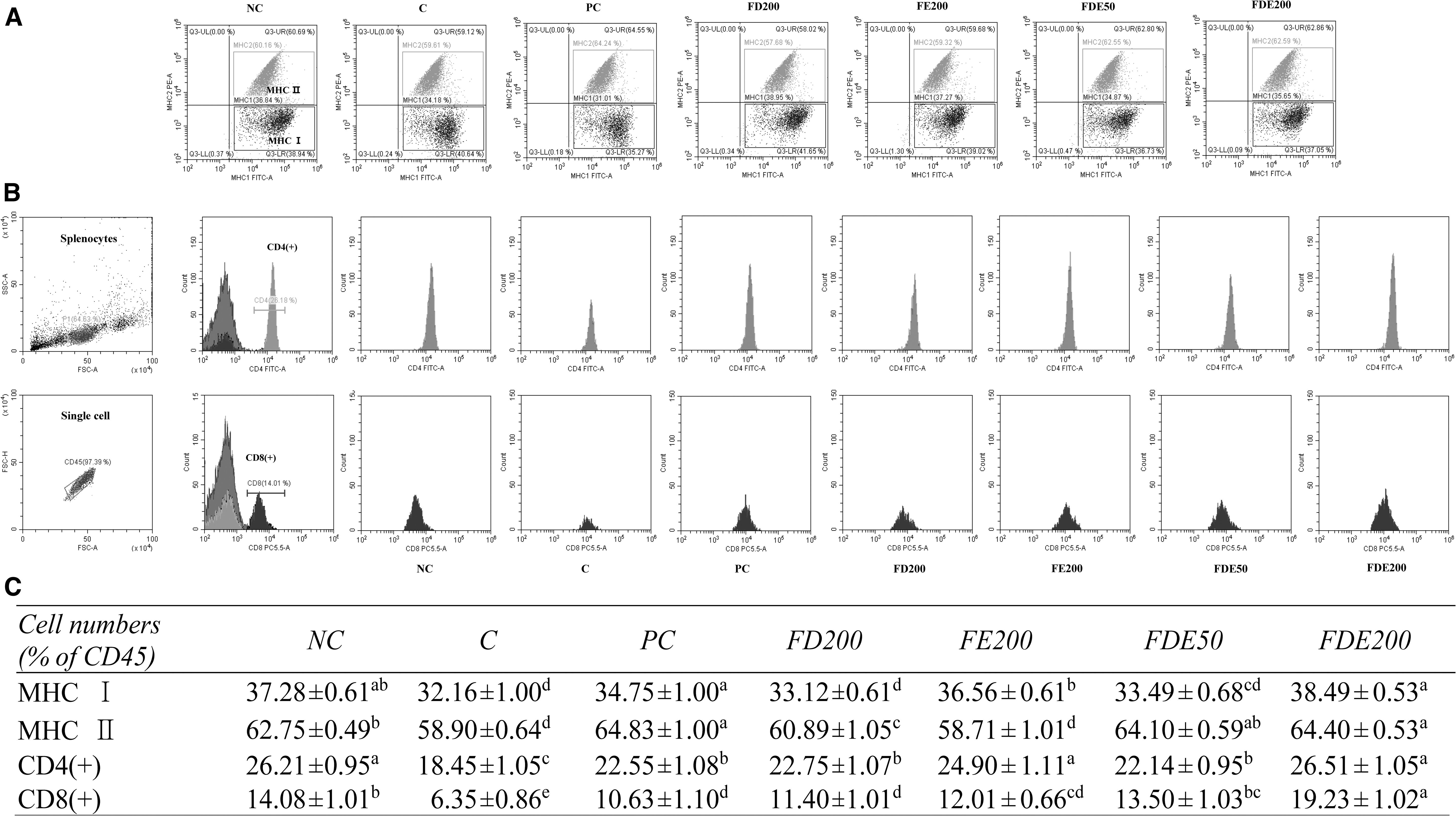
A flow cytometry analysis of MHC I, MHC II, CD4(+), and CD8(+) in splenocytes from C57BL/6N mice with induced immunosuppression by FSE.
CD4(+) T cell counts were significantly decreased in the exercise-C group compared with the NC group and were significantly increased in the PC, FD200, FE200, and FDE groups compared with the exercise-C group. Specifically, CD4(+) T cells were significantly elevated in FDE groups compared with the exercise-C group in a dose-dependent manner. CD8(+) T cells were significantly depressed in the exercise-C group compared with the NC group and were significantly elevated in the PC, FD200, FE200, and FDE groups compared with the exercise-C group. Moreover, CD8(+) T cells of the FDE groups were significantly increased compared with the exercise-C group in a dose-dependent manner (P < .05; Fig. 1B, C).
Effects of FDE on T and B cell proliferation in primary splenocytes in an immunosuppression mouse model
T cell proliferation from Con A-treated splenocytes was significantly depressed in the exercise-C group compared with the NC group and was significantly elevated in the PC, FD200, FE200, and FDE groups compared with the exercise-C group. Specifically, T cell proliferation was significantly elevated in the FDE groups compared with the exercise-C group in a dose-dependent manner. B cell proliferation in splenocytes after LPS treatment was significantly depressed in the exercise-C group compared with the NC group and significantly elevated in the PC group compared with the control group. B cell proliferation was significantly elevated in the FE200 and FDE groups compared with the exercise-C group. However, there was no significant difference between the FDE50 and FDE200 groups (P < .05; Table 2).
Effects of FDE on Splenocytes and Blood Parameters of C57BL/6N Mice with Induced Immunosuppression by Forced Swimming Exercise
Values are presented as mean ± SD (n = 8), and different superscript letters indicate significance at P < .05.
NC; C, induced immunosuppression by FSE; PC (FSE+red ginseng 300 mg/kg body weight); FD200 (FSE+FD extract 200 mg/kg body weight); FE200 (FSE+FE extract 200 mg/kg body weight); FDE50 (FSE+FD and FE 3:7 50 mg/kg body weight); FDE200 (FSE+FD and FE 3:7 200 mg/kg body weight); IFN-γ, interferon gamma; Ig, immunoglobulin; IL, interleukin; NO, nitric oxide; Th, T helper; TNF-α, tumor necrosis factor alpha.
Effects of FDE on Th1-type cytokine production in primary splenocytes in an immunosuppression mouse model
Th1-type cytokine (IL-2 and IFN-γ) production levels are given in Table 2. IL-2 and IFN-γ levels from Con A-treated splenocytes were significantly depressed in the exercise-C group compared with the NC group and were significantly elevated in the PC, FD200, FE200, and FDE groups compared with the exercise-C group. Specifically, the FDE groups had significantly elevated Th1-type cytokines in a dose-dependent manner (P < .05).
Effects of FDE on Th2-type cytokine production in primary splenocytes in an immunosuppression mouse model
Th2-type cytokine (IL-4, IL-6, IL-10, and TNF-α) production levels are given in Table 2. IL-4 and IL-10 levels from Con A-treated splenocytes were significantly elevated in the exercise-C group compared with the NC group and were significantly depressed in the PC, FD200, FE200, and FDE groups compared with the exercise-C group. Specifically, IL-10 levels were significantly depressed in a dose-dependent manner in the FDE groups. IL-6 and TNF-α levels in LPS-treated splenocytes were significantly elevated in the exercise-C group compared with the NC group and were significantly depressed in the PC, FD200, FE200, and FDE groups compared with the exercise-C group. Specifically, IL-6 and TNF-α levels were significantly depressed in a dose-dependent manner in the FDE groups (P < .05).
Effects of FDE on IL-12 and IL-15 cytokines and NK cell activity in primary splenocytes in an immunosuppression mouse model
IL-12 and IL-15 levels from Con A-treated splenocytes were significantly depressed in the exercise-C group compared with the NC group, and significantly elevated in the PC group compared with the C group. Specially, FDE groups had significantly elevated IL-12 and IL-15 production in a dose-dependent manner compared with the exercise-C group (P < .05; Fig. 2B, C).
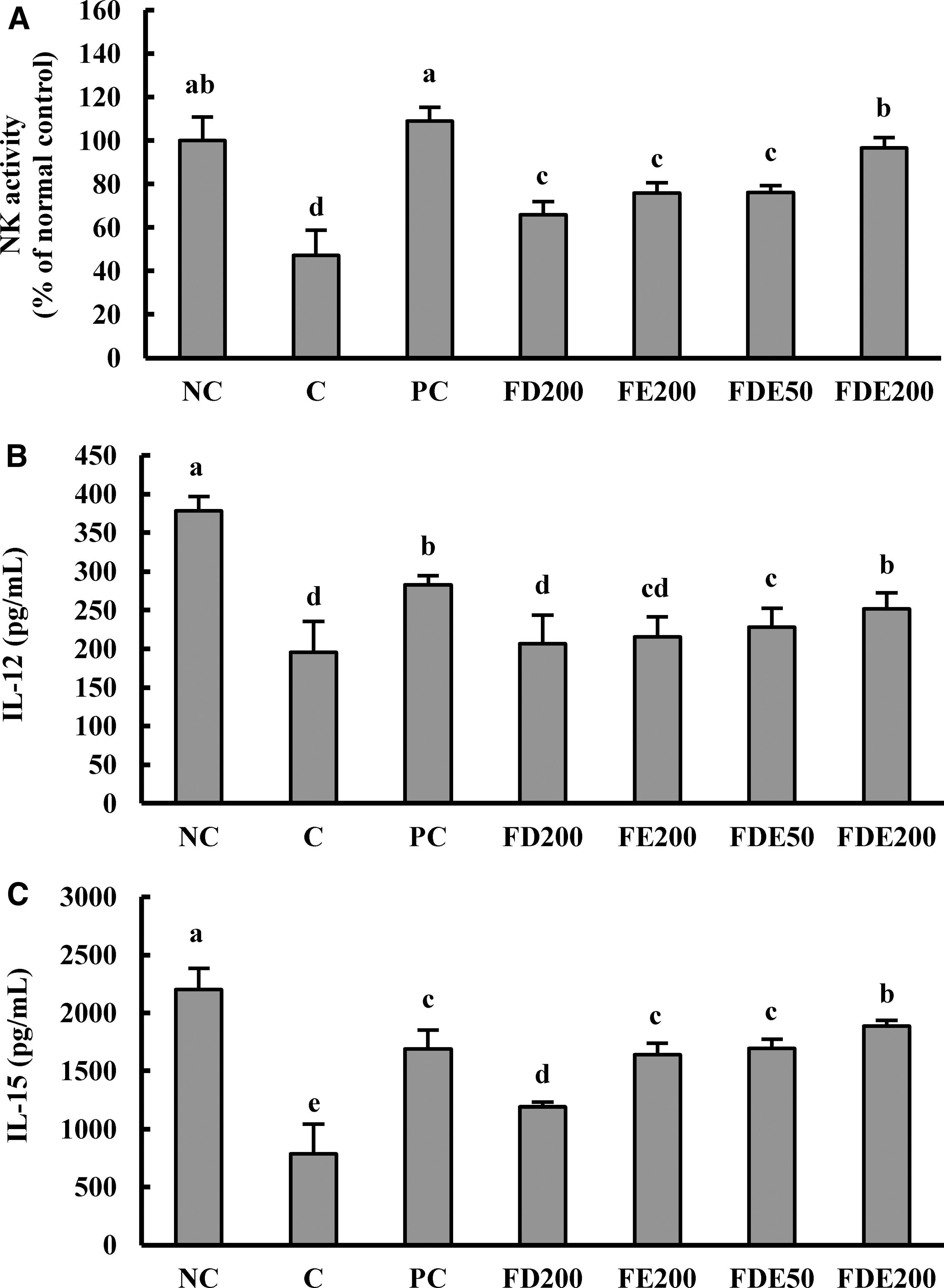
Effects of FDE on NK cell activity
NK cell activity was significantly decreased in the exercise-C group compared with the NC group, and significantly elevated in the PC, FD200, FE200, and FDE groups compared with the exercise-C group. Moreover, FDE groups had significantly increased NK-cell activity in a dose-dependent manner compared with the exercise-C group (P < .05; Fig. 2A).
Effects of FDE on serum NO and Ig levels in an immunosuppression mouse model
Serum NO levels were significantly elevated in the exercise-C group compared with the NC group and were significantly decreased in the PC, FD200, FE200, and FDE groups compared with the exercise-C group. Specifically, FDE groups had significantly depressed NO production in a dose-dependent manner compared with the exercise-C group (P < .05).
IgE, IgA, and IgG levels were significantly decreased in the exercise-C group compared with that in the NC group and were significantly promoted in the PC, FD200, FE200, and FDE groups compared with the exercise-C group. Specifically, FDE groups had significantly promoted IgA and IgG production in a dose-dependent manner compared with the exercise-C group (P < .05; Table 2).
Effects of FDE on blood leukocyte count in an immunosuppression mouse model
Leukocyte numbers (Table 2) were significantly depressed in the exercise-C group compared with the NC group and were significantly elevated in the PC, FD200, FE200, and FDE groups compared with the exercise-C group. Specifically, FDE groups had significantly elevated leukocyte numbers in a dose-dependent manner compared with the exercise-C group (P < .05; Table 2).
DISCUSSION
The aim of this study was to examine the effect of FDE on immunomodulation in C57BL/6M mice that underwent immunosuppression via FSE. At present, health improvement efforts have a strong focus on immunity, and research on natural products that improve immunity is being carried out widely. 2,25,26
Positive factors enhance the body's adaptive immune system, which is a cell-mediated and humoral response. 27 In contrast, negative effects, such as irregular exercise, inhibit lymphocyte proliferative responses, aggravate inflammatory responses, and cause hyper-immune reactions like asthma and allergic reactions. 12,28,29 One study showed that lymphocyte proliferation in mitogen-stimulated splenocytes was significantly reduced compared with the control group after 2 h of forced swimming in BALB/c mice without preliminary exercise. 30
Many natural substances are known to improve immune function, including deer velvet, which has an anti-inflammatory effect and promotes hematopoietic function. 31,32 In addition, the fatty acid chains of phosphatidylcholine found in deer velvet affect lymphocyte proliferation to regulate immune responses and promote macrophage phagocytosis. 33,34 Deer velvet loses many of its active ingredients during the chopping process, and research on FD was initiated to explore whether a different processing approach could preserve its active ingredients. 35 Recent studies have shown that fermenting antler inhibits osteoclast differentiation and enhances splenocyte survival rates and IL-12 production. 17,18
Eleutherococcus senticosus is another substance related to immune enhancement owing to eleutheroside and lignin active ingredients. 36,37 It was reported to have various pharmacological effects such as anti-diabetes, anti-obesity, and antioxidant effects; however, research on FE is still lacking. 38 –40 Furthermore, no studies have yet explored the immunopromoting effects of an extract mixture of FD and FE (FDE). Therefore, the aim of this study was to examine the effect of FDE on immunomodulation in C57BL/6M mice in which immunosuppression was induced through FSE.
In this experiment, oral FDE intake induced a significant increase in the number of MHC I, MHC II, CD4(+), and CD8(+) T cells as measured by flow cytometry compared with the swim control mice (P < .05). T cells were classified as CD8(+) cytotoxic T cells and CD4(+) T helper cells, which cause apoptosis in adaptive immunity. When antigens are observed in APCs, MHC molecules play an important role in T cell selection and interact with CD8(+) cytotoxic T cell or CD4(+) T helper cell coreceptors. 41 MHC I molecules are consistently observed in all nucleated cells, whereas MHC II molecules are only present in APCs, such as B cells, dendritic cells, macrophages, and thymic epithelial cells. Although the molecular functions of MHC I and II are similar, they bind to CD8(+) cytotoxic T cells and CD4(+) T helper cells, respectively, to respond to specific antigens. 42
Acute exhaustive exercise is known to decrease NK cell and macrophage function, and to decrease the ratio of CD4(+), a T helper cell co-factor for T lymphocytes, and CD8(+), a co-factor for cytotoxic T cells. 43 In this study, oral FDE intake also induced a significant increase in the NK cell activity compared with swim control mice (P < .05). CD8(+) cytotoxic T cells directly attack abnormal cells, such as NK cells, tumor cells, and virus-infected cells. 44
CD4(+) T helper cells are classified as Th1 cells or Th2 cells. Th1 cells produce Th1-type cytokines, which stimulate phagocytosis mainly by increasing macrophage activity, and Th2 cells produce Th2-type cytokines, which stimulate B cells and induce antibody production. Immunity function is affected by the balance of Th1-type and Th2-type cytokines and their complementary regulation. 45 Th1-type cytokines, such as IL-2 and IFN-γ, stimulate CD8(+) cytotoxic T cell proliferation, CD4(+) T helper cell differentiation, and macrophages leading to antigen elimination. 46,47 Th1-type cytokines, such as IL-12 and IL-15, stimulate NK cell activity and eliminate the APCs found in abnormal cells in endogenous immunity. NK cells bind to virus-infected or abnormal cells through active or inhibitory signals and attack APCs by receiving active signals. 48 In this study, oral FDE intake induced a significant increase in Th1-type, IL-12, and IL-15 cytokine levels compared with swim control mice (P < .05). These results suggest subsequent CD8(+) cytotoxic T cell activation, NK cell activation, and antigen elimination. Th2-type cytokines such as IL-4, IL-10, IL-6, and TNF-a are produced at abnormally high levels in immunosuppression models. 49 In particular, inflammatory cytokines such as IL-6 and TNF-α activate nuclear factor kappa B and regulate immune cell activity. 5,6 In this study, oral FDE intake induced a significant decrease in IL-4, IL-6, IL-10, and TNF-α cytokine levels and NO production compared with the swim control mice (P < .05). This result is consistent with the finding that high intensity short-term exercise increased NO production. 50 In addition, these results suggest a subsequent decrease in macrophage activation, inflammatory response, and apoptosis. Therefore, we hypothesized that FDE has immunomodulation ability by regulating the balance between Th1 and Th2 cytokines.
High-intensity exercise is known to reduce leukocyte numbers as well as B cell and T cell function, which induce antibody production and affect Ig production, 51,52 including IgA, IgD, IgG, IgE, and IgM. Their basic structure and function are similar, but their size, composition, and specific functions are different. IgG is the major serum Ig, IgM is predominant in primary immunity and has very large-sized molecules. IgA is also responsible for body surface immunity, whereas IgD is involved in cell binding, and IgE is involved in allergic reactions. 53,54 In this study, oral FDE intake induced a significant increase in leukocyte numbers and significantly promoted IgA and IgG levels compared with the swim control mice.
In summary, we investigated the immunomodulatory effects of FDE mixtures in C57BL/6N mice that underwent immunosuppression through FSE. Specifically, we presume that eleutheroside and lignin, which are active FE ingredients, improved CD8(+) cytotoxic T cell and CD4(+) T helper cell function in this FSE model. In particular, we found that oral FDE intake increased MHC I, MHC II, CD4(+) T cell, and CD8(+) T cell counts. Moreover, FDE increased Th1 type cytokines, IL-12 and IL-15 levels, NK cell activity, B cell proliferation, and leukocyte counts compared with the swim control mice. FDE also changed Ig production levels (Fig. 3). Therefore, we conclude that FDE is effective for immunomodulation and may be developed as a natural functional substance.
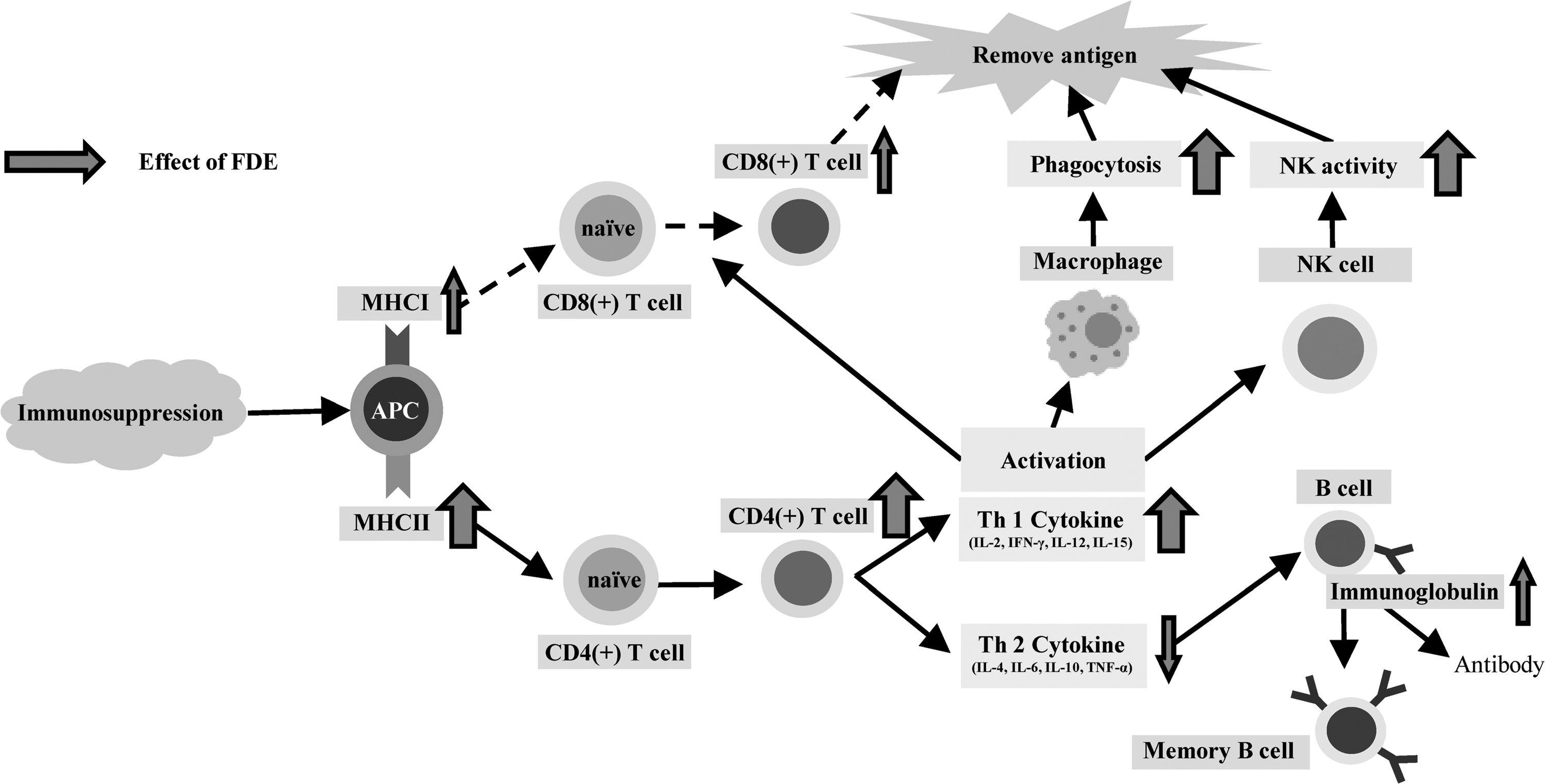
Effects of FDE on immunosuppression. CD, cluster of differentiation; IFN-γ, interferon gamma; Th, T helper; TNF-α, tumor necrosis factor alpha.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by the collaborative R&BD program (2018) of the Agency for Korea National Food Cluster (AnFC).