
Abstract
We evaluated the effect of artichoke leaf extract (ALE) on the livers of mice with non-alcoholic fatty liver disease (NAFLD) induced by high-fat/high-fructose diet and H2O2-treated HepG2 cells, as well as the mechanism underlying its hepatoprotective effects. Supplementation with ALE suppressed the NAFLD-induced increases in serum lipids, bilirubin, gamma-glutamyl transferase, aspartate transaminase (AST), and alanine aminotransferase. In addition, we observed that supplementation with ALE attenuated the increases in antioxidant enzyme activity, mRNA levels of proinflammatory cytokines, and apoptosis signaling pathways caused by a high-fat/high-fructose diet. We found that ALE treatment suppressed inflammation and apoptosis caused by H2O2-induced oxidative stress in HepG2 cells. These findings suggest that ALE supplementation directly suppresses inflammation and apoptosis in hepatocytes during the development of NAFLD. Based on these results, we suggest that supplementation with ALE may be useful for preventing the progression of liver diseases, including hepatic steatosis and non-alcoholic steatohepatitis.
Introduction
Non-alcoholic fatty liver disease (NAFLD) is the most important cause of chronic liver disease and has an increasing prevalence worldwide. 1 The pathogenesis of NAFLD is a multistep process involving fatty liver with fat accumulation exceeding 5% along with non-alcoholic steatohepatitis (NASH) and cirrhosis, without excessive alcohol consumption. 2 Because Western diets, including high-calorie, high-fat, high-fructose, and high-sucrose diets, are associated with NAFLD development, nutrition plays a key role in the pathogenesis or prevention of NAFLD. NAFLD is strongly associated with metabolic dysfunction, including insulin resistance, dyslipidemia, and cardiovascular disease. 3,4
The liver is a major organ in lipid homeostasis responsible for fatty acid synthesis, export, and transfer to non-liver tissues by very-low-density lipoprotein as well as their utilization. 5,6 Lipid homeostasis in the liver is tightly regulated; however, the disruption of lipid metabolism pathways results in the development and progression of hepatic steatosis. 5,6 In addition, NASH is caused by several factors related to increased reactive oxygen species (ROS), including inflammation, oxidative stress, and toxicity. 7,8 Steatosis conditions are considered relatively benign, while conditions that promote the development of NASH are associated with a higher risk of cirrhosis and hepatocellular carcinoma. 9 Because NASH is characterized by multifactorial disorders, treatment options are limited, and individually tailored approaches need to be considered. 8 In addition, several studies have suggested alternative approaches, such as the use of phytochemicals with multipronged mechanisms of action. 10,11
We investigated the effects of supplementation with artichoke leaf extract (ALE) on mice with high-fat/high-fructose diet-induced NAFLD. Artichoke (Cynara scolymus) belongs to the family Asteraceae and is a typical component of a traditional Mediterranean diet. Several studies have reported beneficial health effects of artichokes, which contain various phenolic compounds, including antioxidant, anti-inflammatory, anticancer, antibacterial, and hepatoprotective effects. 12 –14 However, there are few mechanistic studies that have investigated the hepatoprotective effects of artichokes; accordingly, we evaluated the effects of ALE on lipid accumulation, antioxidant enzyme activity, and inflammation/apoptosis signaling in livers of mice with NAFLD and H2O2-treated HepG2 cells.
Materials and Methods
Artichoke dry extract preparation and standardization
Fresh artichoke leaves were milled, followed by extraction with hot water (1:5, w/w). The extract was dried and concentrated to 65–85%. The concentrated extract was dried at 45–60°C in a fluid bed dryer. The dried extract was milled to obtain a homogeneous particle size (Cynarol; Niak Pharmaceuticals, Gorgan, Iran). The artichoke extract was standardized using 1.66 ± 0.03 mg/g of cynarin (Fig. 1) by high performance liquid chromatography (HPLC) (Santa Clara, CA).
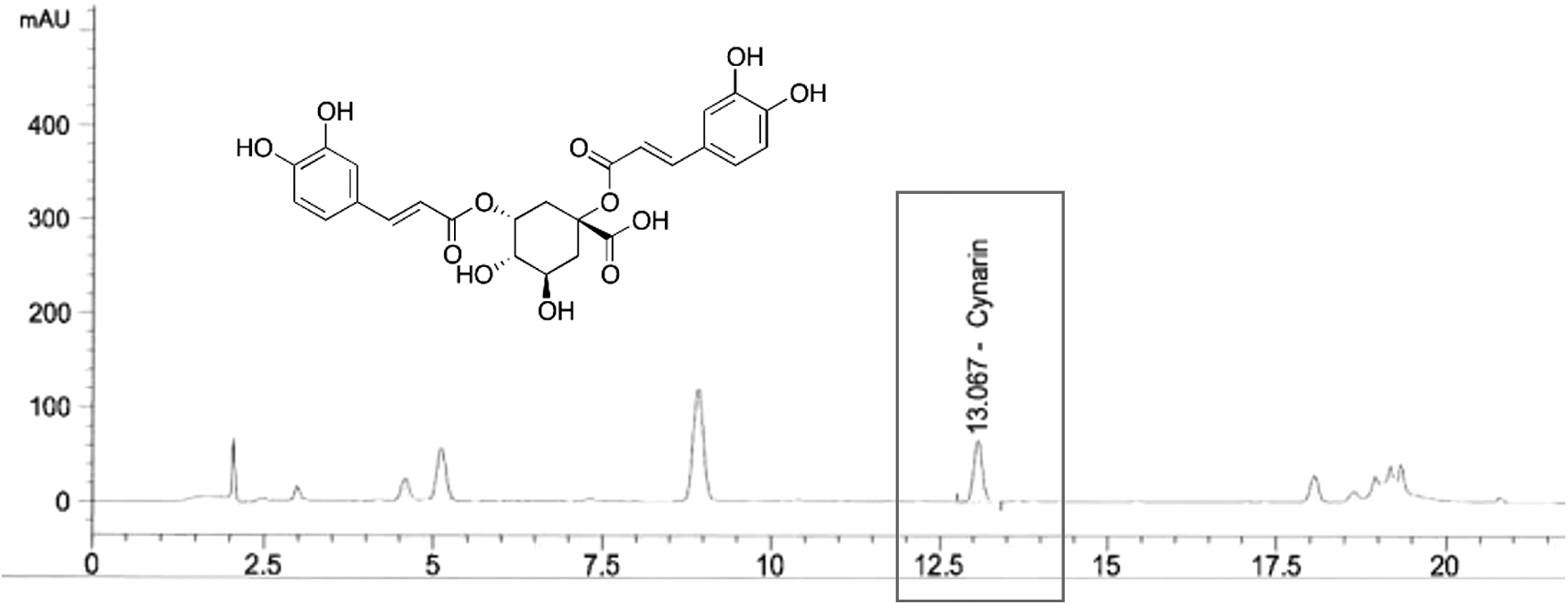
High-performance liquid chromatography analysis of cynarin in artichoke extract.
Animals
C57BL/6 mice (6-week-old male) were nurtured in a controlled environment at 22–25°C with alternating 12-h light and 12-h dark cycles. The mice were fed a commercial pelleted chow and were provided water ad libitum. The Institutional Animal Care and Use Committee of Kyung Hee University (KHUASP-20-194) approved the protocol for the animal study. Mice were divided into five groups (n = 8): normal diet (NC); 60% high-fat diet (HFD) +10% fructose in water (control, C); 60% HFD +10% fructose in water +30 mg/kg b.w. silymarin extract (positive control); 60% HFD +10% fructose in water + ALE 50 mg/kg b.w. (ALE 50); and 60% HFD +10% fructose in water + ALE 150 mg/kg b.w. (ALE150). At the end of 14 weeks, mice were sacrificed, and the liver and blood (by orbital venipuncture) were collected for analysis.
Hematoxylin and eosin staining
The liver tissues were fixed and embedded in paraffin. The paraffin blocks were sliced into 4 μm sections. The sections were stained with hematoxylin and eosin (H&E) for the measurement of lipid droplets.
Serum biomarker analyses
Serum was collected, and then levels of lipid profile were measured using quantification kits from Biomax, Inc., (Seoul, Korea). Aspartate transaminase (AST), alanine aminotransferase (ALT), gamma-glutamyl transferase (GGT), and bilirubin levels were measured using quantification kits from Bio-Rad Laboratories (Hercules, CA).
Measurement of antioxidant enzyme activity
The antioxidant enzyme activity in the liver was measured using OxiTec™ Superoxide dismutase (SOD), OxiTec catalase (CAT), and OxiTec glutathione peroxidase (GPx) assay kits (Biomax, Inc.).
Cell culture and measurement of hepatoprotective activity
HepG2 cells (ATCC HB-8065) were obtained from the American Type Culture Collection (Rockville, MD). HepG2 cells were cultured with ALE at various concentrations. After incubation for 24 h, cells were treated with 500 μM H2O2. After 4 h, 500 μg/mL 3-(4,5-dimethylthiazol-2yl-)-2,5-diphenyl tetrazolium bromide (MTT) was added to each well, and then the supernatants were removed after 4 h. Dimethylsulfoxide (DMSO) 200 μL was added to each well, and the results are expressed as a percentage of values for the control group.
Real-time polymerase chain reaction
Total RNA from cells and livers of mice was isolated using TRIzol reagent. cDNA synthesis was performed using an iScript™ cDNA Synthesis Kit (Bio-Rad). PCR amplification consisted of 40 cycles of 95°C for 15 sec, 58°C for 15 sec, and 72°C for 30 sec with SYBR Green PCR Master Mix (Bio-Rad) and primer pairs (Table 1). The data analysis was performed using CFX Maestro (Bio-Rad).
Primer Sequences Used for mRNA Quantification by Real-Time Polymerase Chain Reaction
GAPDH, glyceraldehyde 3-phosphate dehydrogenase; IL, interleukin; TNF-α, tumor necrosis factor-alpha.
ELISA assay for proinflammatory cytokines
Levels of interleukin-1β (IL-1β), IL-6, and tumor necrosis factor-alpha (TNF-α) in the serum from mice were measured by using the R&D Duoset ELISA development kits (R&D Systems, Minneapolis, MN).
Western blot analysis
The liver tissues and cells were lysed using CelLytic MT cell lysis reagent. Fifty micrograms protein was separated using 10% Mini-PROTEAN®TGX™Precast Protein Gel (Bio-Rad Laboratories) and electrotransferred onto polyvinylidene difluoride (PVDF) membranes (Bio-Rad Laboratories). Membranes were blocked with 5% skimmed milk in tris-buffered saline with 0.1% tween 20 (TBST) for 1 h, and then incubated for 12 h at 4°C with antibodies against p-JNK, p-p53, p-p38, CD95, cleaved-caspase 3, cleaved-caspase 8, COX-2, iNOS, IκB, p-p65, and beta-actin (1:1000; Cell Signaling, Danvers, MA). The membranes were incubated with a secondary antibody (Cell Signaling Technology, Inc.) for 1 h at room temperature. The protein bands were marked with EzWestLumi Plus (ATTO, Tokyo, Japan) detection reagents and developed using Ez-Capture II (ATTO). The bands were quantified using CS Analyzer 3.0 (ATTO).
Statistical analysis
All data are expressed as mean ± standard deviation. Differences among groups were evaluated by one-way analysis of variance (ANOVA) and Duncan's multiple range tests implemented in SPSS for Windows (SPSS PASW Statistic 22.0, SPSS, Inc., Chicago, IL). Differences were considered significant at P < .05.
Results
Effects of artichoke on lipid profiles and liver function in mice with NAFLD
We found that levels of triglyceride (TG), total cholesterol (TC), and low-density lipoprotein (LDL) cholesterol were higher in the control group (C) than in the group fed the normal diet (NC). However, ALE supplementation resulted in significantly lower serum levels of TG, TC, and LDL cholesterol and higher serum levels of HDL cholesterol than those in the group fed a high-fat/high-fructose diet (Fig. 2A–D) (P < .05). Bilirubin, GGT, AST, and ALT levels are useful markers of NAFLD and liver damage. Levels of these liver function markers were higher in the control group than in the group fed the normal diet. ALE supplementation resulted in significantly lower levels of bilirubin, GGT, AST, and ALT than those in the group fed a high-fat/high-fructose diet (Fig. 2E–H) (P < .05). These results suggest that supplementation with ALE has protective effects on the livers of NAFLD-induced mice.

Serum triglyceride
Effects of artichoke on lipid accumulation, antioxidant enzymes, and proinflammatory cytokines in livers from mice with NAFLD
We measured the liver weight and lipid accumulation in the livers of mice with NAFLD. Figure 3 shows that liver weight and lipid accumulation in the liver were higher in the control group than in the normal control group. We found that there is no significant difference in liver weight between NC group and ALE150 group, and silymarin and ALE supplementation significantly decreased lipid accumulation in the livers of mice with high-fat/high-fructose diet-induced NAFLD. In addition, activities of the antioxidant enzymes SOD, GPx, and CAT were significantly lower in groups with ALE supplementation than in the control group, indicating that ALE suppressed oxidative stress (Fig. 3C–E) (P < .05). When compared with levels in the control group, the groups with ALE supplementation had significantly lower mRNA levels of IL-1β, IL-6, and TNF-α and serum levels of IL-1β, IL-6, and TNF-α (Fig. 3F–K) (P < .05). These findings suggest that ALE can suppress lipid accumulation, oxidative stress, and inflammation in the liver of mice with NAFLD.
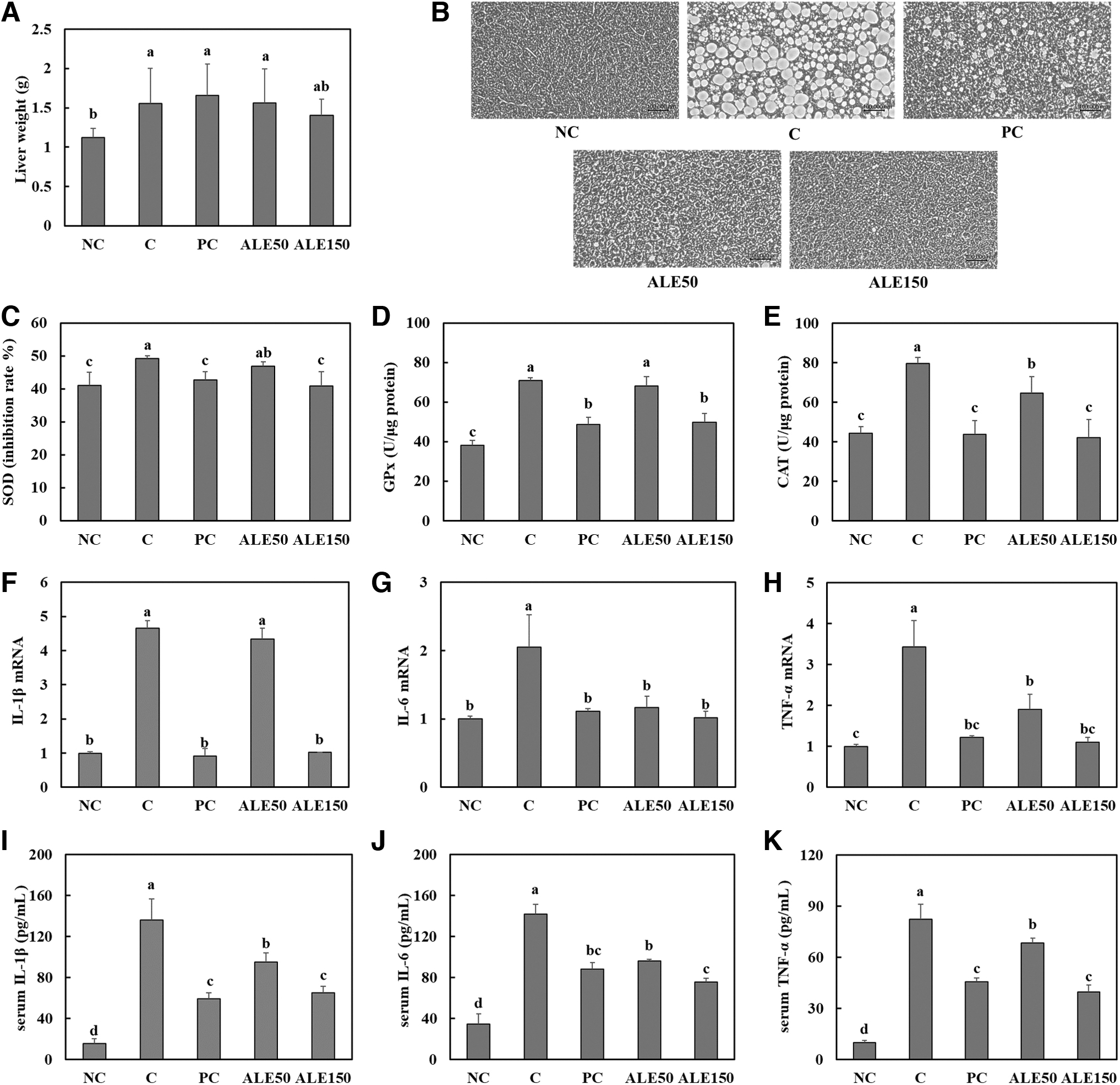
Liver weight
Effects of artichoke on apoptosis signaling pathways in livers of mice with NAFLD
We found that the high-fat/high-fructose diet stimulated apoptosis signaling pathways, including the JNK-p53-caspase3 pathway and p38-Fas/CD95-caspase8-caspase3 pathway. When compared with levels in the control group, the groups with silymarin and ALE supplementation showed significantly lower levels of proteins in apoptosis signaling pathways (Fig. 4) (P < .05).

Protein expression levels of apoptosis signaling pathway components determined by Western blotting (band images;
Effects of artichoke on inflammation in H2O2-treated HepG2 cells
H2O2 led to an increase in the protein concentrations of inflammatory mediators NF-κB (p65), COX-2, and iNOS. However, ALE treatment suppressed the protein expression of inflammatory mediators in H2O2-induced inflammation (Fig. 5A, B) (P < .05). In addition, we found that ALE treatment induced significant decreases in mRNA expression levels of IL-1β, IL-6, and TNF-α and the secreted levels, compared with levels in the control (treatment with H2O2) (Fig. 5C–H) (P < .05).
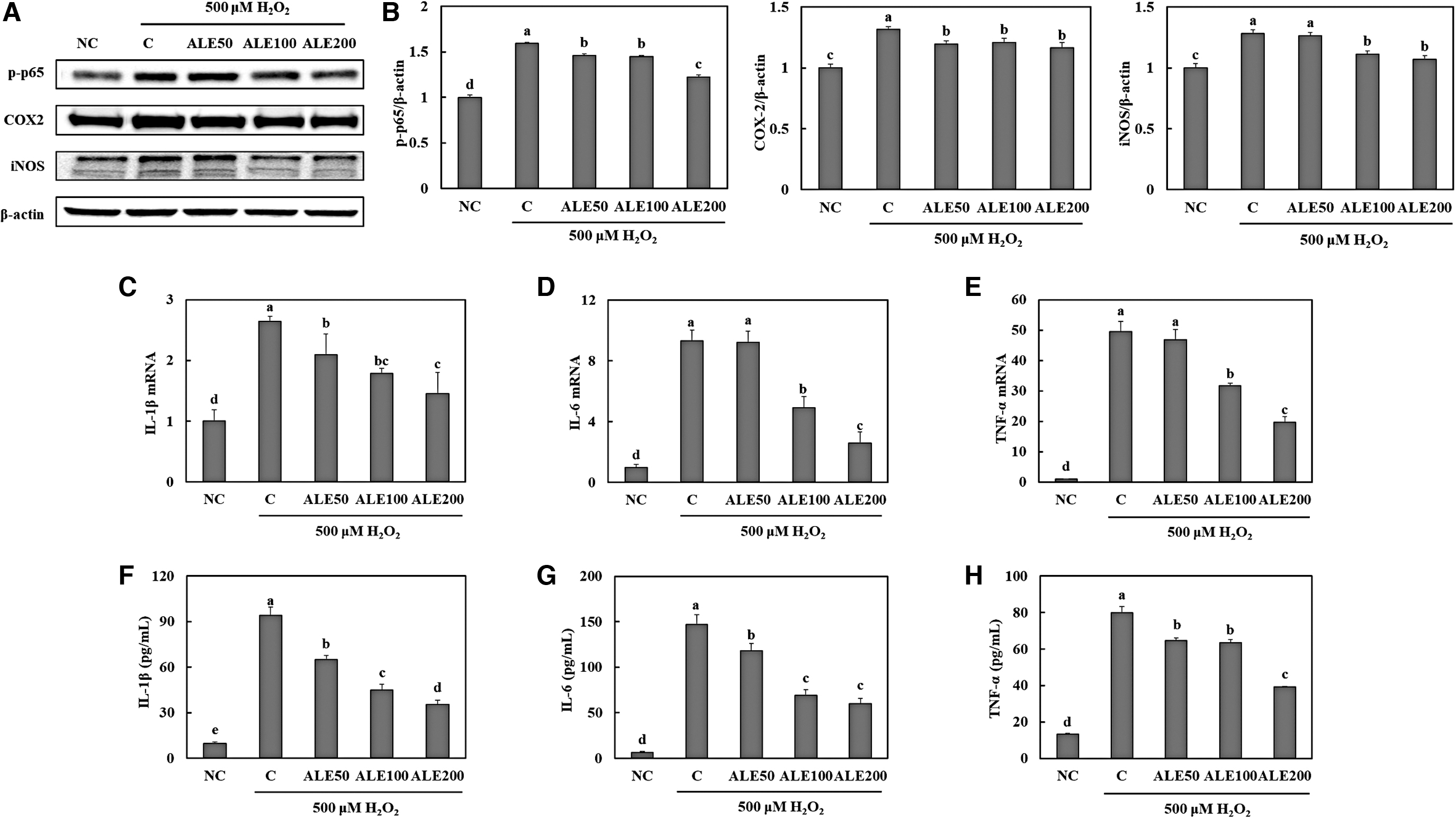
Expression levels of proteins in inflammation pathways determined by Western blotting (band images;
Effects of artichoke on apoptosis signaling pathways in H2O2-treated HepG2 cells
We found that treatment with H2O2 stimulated apoptosis signaling pathways, including the JNK-p53-caspase3 pathway and p38-Fas/CD95-caspase8-caspase3 pathway, in HepG2 cells. However, ALE treatment suppressed the expression of proteins involved in the apoptosis pathways in H2O2-treated HepG2 cells (Fig. 6) (P < .05).
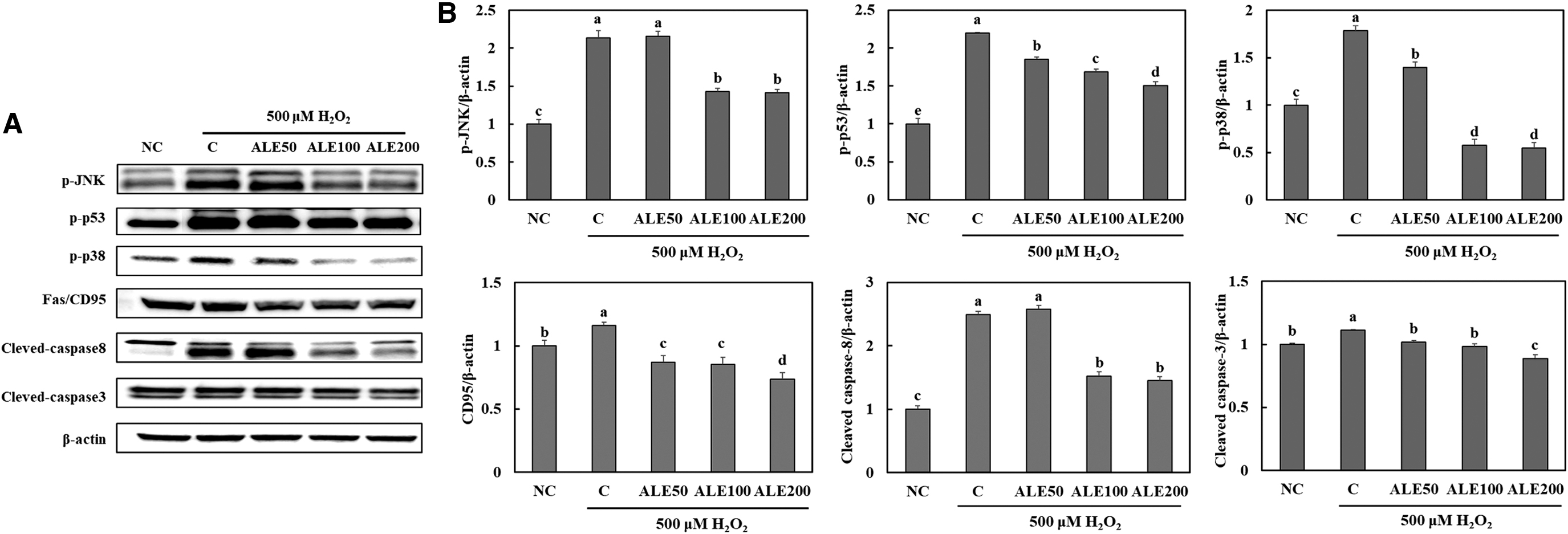
Expression levels of proteins in apoptosis signaling pathways determined by Western blotting (band images;
These data indicate that ALE supplementation directly suppresses inflammation and apoptosis in the liver during the development of NAFLD.
Discussion
In this study, we used a mouse model of high-fat/high-fructose diet-induced NAFLD to investigate the effect of ALE on the induction of NASH as well as hepatic steatosis. Abdelmalek et al. demonstrated that increased daily fructose consumption stimulates the development of hepatic fibrosis, but reduces hepatic steatosis in patients with NAFLD. 15 In addition, Liu et al. found that a high-fat/high-fructose diet significantly increases the levels of AST, ALT, TC, LDL cholesterol, and NASH-related histological parameters compared with levels in various controls (high fat-only diet, high fructose-only diet, and standard chow diet). 16 Consistent with these previous results, we confirmed that a high-fat/high-fructose diet caused hepatic steatosis, hepatic inflammation, and liver damage in mice. We observed that the high-fat/high-fructose diet increased the serum lipid profile, bilirubin, GGT, AST, and ALT, liver tissue weight, and lipid accumulation in the liver. However, supplementation with ALE suppressed the steatosis and the increased serum lipid profile, bilirubin, GGT, AST, and ALT induced by a high-fat/high-fructose diet. We investigated the mechanism of hepatoprotective effects of artichokes associated with oxidative stress, inflammation, and apoptosis.
Normal and healthy hepatocytes are equipped with an antioxidative defense system consisting of enzymatic (SOD, CAT, and GPx) and non-enzymatic factors. 17 In this study, we found that a high-fat/high-fructose diet stimulated the activity of SOD, CAT, and GPx, indicating that this diet triggered the activation of the antioxidative defense enzymatic system to cope with excessive ROS production. Cordero-Herrera et al. have reported that excessive ROS production can inhibit the expression of phosphorylated AMP-activated protein kinase (AMPK), which induces the development of hepatic steatosis. 18 In addition, Schuste et al. have reported that overproduction of ROS triggers the production of proinflammatory cytokines, which play an important role in NASH progression. 19 We observed that supplementation with ALE attenuated the increases in mRNA levels of proinflammatory cytokines caused by a high-fat/high-fructose diet. Thus, we suggest that supplementation with ALE inhibits the progression of NASH, a combination of hepatic steatosis and severe inflammation, in NAFLD by inhibiting oxidative stress and inflammation in the liver.
Several studies have shown that a characteristic feature in the liver of patients with NASH is apoptosis by a combination of ROS overproduction, inflammatory cytokines, and free fatty acids. 17,20,21 Feldstein et al. reported that apoptosis is induced in the liver of patients with NASH and is correlated with the severity of liver disease. 21 In addition, several studies have demonstrated that inhibition of apoptosis suppressed the development of fibrosis in the liver with NASH. 22,23 Lipotoxicity in the liver activates both the extrinsic pathway and intrinsic pathway of apoptosis, which play a key role in cell death during the progression of NASH. 24 The results of this study showed that supplementation with ALE inhibits both the JNK-p53-caspase3 and p38-Fas/CD95-caspase8-caspase3 apoptosis signaling pathways, involved in NASH-induced hepatocyte death. We suggest that supplementation with ALE may suppress NASH progression in NAFLD by antioxidant and anti-inflammatory effects.
We used H2O2-induced oxidative stress in HepG2 cells to evaluate whether ALE supplementation in NAFLD directly affects hepatocytes. We found that treatment with H2O2 increased the protein expression of inflammatory mediators, mRNA expression of proinflammatory cytokines, and expression of apoptosis-related proteins in HepG2 cells. However, ALE treatment suppressed inflammation and apoptosis under H2O2-induced oxidative stress in HepG2 cells, suggesting that ALE directly suppresses inflammation and apoptosis in hepatocytes during NAFLD development. Taken together, our results suggest that supplementation with ALE may be useful for preventing the progression of liver diseases, including hepatic steatosis and NASH.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.