
Abstract
Ancient traditions showed that fermented enzyme foods have beneficial health effects on the body. However, only a few studies have reported on its impact on weight loss and metabolic syndrome. Therefore, it is necessary to verify whether diet supplementation with fermented enzyme foods can have a beneficial functional impact on the body. We examined the antiobesity properties of fermented mixed grain (FMG) with digestive enzymes (FMG) in diet-induced obese mice. Sixty C57BL/6J mice were randomly assigned to six dietary groups: (1) normal diet (ND), (2) high-fat diet (HFD), (3) Bacilus Coagulans, (4) steamed grain, (5) low-dose FMG (L-FMG), and (6) high-dose FMG (H-FMG) supplement for 12 weeks. The results showed that H-FMG supplement dramatically decreased body weight and fat mass with simultaneous decreases in plasma lipid contents. Furthermore, H-FMG significantly lowered fasting blood glucose concentrations and improved glucose tolerance compared with the HFD group. Also, the concentrations of inflammatory cytokines secreted from adipocytes in H-FMG–supplemented mice decreased dramatically. Taken together, our findings indicated that H-FMG can ameliorate HFD-induced obesity and its associated complications and could be used as a potential preventive intervention for obesity.
Introduction
Obesity has emerged as the most important health epidemic of modern society, which is one of the highest risk factors for increased metabolic diseases and other associated health complications. 1 Disorders of adipose tissue metabolism in obesity and metabolic syndrome are very important in the initiation of inflammatory response. Inflammation increases the secretion of the cytokine, monocyte chemotactic protein-1 (MCP-1), which causes macrophage infiltration in adipose tissue. 2,3 Activated macrophages in the adipose tissue, both resident and immigrant, further initiate the secretion of inflammatory cytokines through nuclear factor-κB (NF-κB) signaling. 4 Metabolic syndrome, caused by obesity, leads to its complications such as type nonalcoholic fatty liver disease (NAFLD), type 2 diabetes mellitus (T2DM), cardiovascular disease, and certain obesity-related cancers. 5 Many studies have indicated that Toll-like receptors (TLRs), the innate immune receptors, activate the innate immunity, regulate cytokine expression, and indirectly activate the adaptive immune systems. 6,7 Especially, TLRs lead to the chronic inflammatory state derived from obesity and its complications. 8
One of the best approaches to weight management is to inhibit dietary lipid absorption with Orlistat being prescribed to treat obesity for the past 10 years. 9 Even if body weight loss due to Orlistat remains moderate, it meaningfully reduces the risk factors for obesity and its complication. 10 Furthermore, Orlistat has no serious side effects associated with appetite-suppressing drugs. 9 These results have prompted investigation into new generation food and pharmaceutical agents that disrupt the digestion of dietary carbohydrates and fats in the gut.
The weight loss is triggered by carbohydrate digestion inhibitors with the improvement of glycemic control. 11 However, side effects from colon fermentation, such as as gas production, remain a problem.
In contrast, fermented enzyme foods have recently been suggested as a solution to improve obesity in Eastern populations; however, there is no scientific evidence to support this suggestion. Therefore, we investigated whether fermented enzyme foods, including digestive enzymes, have antiobesity effects in high-fat diet (HFD) in mice.
Materials and Methods
Animal study
Sixty male C57BL/6J mice of age 4 weeks (Jackson Laboratory, Bar Harbor, ME, USA) were used in our study. All mice were housed at ∼24°C and a 12-h light/dark cycle and fed a standard chow diet for acclimation and, then, randomly assigned to six groups. Each group was fed one of the following diets for 12 weeks: normal diet (ND, 10% kcal from fat, n = 10); HFD (40% of kcal from fat, n = 10); Bacilus Coagulans [BC, HFD with 0.3% Bacilus Coagulans (w/w), n = 10]; 0.3% steamed grain [SMG, HFD with 0.3% steamed grain (w/w), n = 10]; low-dose fermented mixed grain [L-FMG, HFD with 0.3% fermented mixed grain (w/w)]; and high-dose fermented mixed grain [H-FMG, HFD with 0.9% fermented mixed grain (w/w)]. This study is approved by the Kyungpook National University Industry Foundation (KNU-2019-0059).
Preparation of samples
To prepare BC, Bacillus Coagulans KCTC13284BP was cultivated in Luria Bertani broth at 37°C and centrifuged. The pellet was mixed with maltodextrin and dried using a freeze dryer (LP100; IlShin BioBase Co. LTD., Dongducheon, Korea). For SMG, the six kinds of grains (Sanlim global Corp., Yongin, Korea), brown rice, barley, soybean, mixed grains, including wheat, job's tears, and corn were steamed with water at 120°C and dried at 60°C. To prepare FMG, the same six kinds of grains used for SMG were fermented separately in the solid and liquid state with Bacillus Coagulans in an aseptic ventilation facility at 35°C and dried at 60°C. The dried fermented grains were then mixed at a constant ratio (brown rice 35%, barley 25%, soybean 20%, and mixed grains 20%), and the Multibiozyme (Bision Corp., Seongnam, Korea) 1.8% was added to supplement digestive enzyme activity.
Alpha-amylase, protease, and lipase activity (unit/g) were measured according to the general test method of the Korea food standard codex from the ministry of food and drug safety (MFDS) and were shown in Table 1. The α-amylase activity was examined as the amount of enzyme that releases 1 μmol of glucose from starch per minute. Protease activity was examined as the amount of enzyme that releases 1 μmol of
Digestive Activities in Samples
SMG (HFD +0.3% steamed mixed grain, wt/wt); L-FMG (HFD +0.3% fermented mixed grain (wt/wt); H-FMG (HFD +0.9% fermented mixed grain (wt/wt).
Plasma, hepatic, and fecal lipid profile
Plasma triglyceride (TG), total cholesterol (TC), and high-density lipoprotein cholesterol (HDL-C) concentrations were determined by a commercial kit (Asan, Seoul, Republic of Korea). The plasma free fatty acid (FFA) content was measured by a commercial kit (Wako Chemicals, Richmond, VA, USA). The levels of plasma apolipoprotein A-I (Apo A-I) and apolipoprotein B (Apo B) were determined by a commercial kit (Nittobo Medical Co., Ltd.). We extracted hepatic and fecal lipid contents according to Folch's methods. 12 Hepatic and fecal lipid contents were determined with the same commercial kits as for plasma lipid profile analysis.
Activities of lipid-regulating enzymes in liver and epididymal white adipose tissue
Samples were prepared and analyzed with the method developed by Hulcher and Oleson. 13 Fatty acid synthase (FAS) activity was analyzed according to the method by Nepokroeff et al. 14 The activities of glucose-6-phosphate dehydrogenase (G6PD), 15 malic enzyme (ME), 16 carnitine palmitoyl transferase (CPT), 17 and fatty acid β-oxidation 18 were measured using the previously described method.
Measurement of metabolic rate
Indirect calorimetry analysis was performed for measuring the energy expenditure 2 weeks before sacrifice using the OxyletProTM System (PANLAB, Cornellà, Spain). The mice were transferred to chambers, with free access to water and food at 25°C.
Plasma adipokine and inflammatory cytokine
Plasma leptin, adiponectin, resistin, and plasminogen activator inhibitor-1 (PAI-I) levels were measured using a multiplex detection kit (Bio-Rad Laboratories, Inc., Hercules, CA, USA). We used the Luminex 200 LabMAP system (Luminex, Austin, TX, USA) and the Bio-Plex Manager software version 4.1.1 (Bio-Rad Laboratories, Inc.) for analysis.
White adipose tissue and hepatic morphology
Fixed in 10% buffered formalin, white adipose tissue (WAT) and liver samples were embedded in paraffin, sectioned at 4 μm, stained with hematoxylin and eosin and Masson's trichrome, and observed under an optical microscope (Nikon, Tokyo, Japan) with a magnifying power of 200 × .
mRNA expression analysis
The mRNA extraction method has previously been described. 19 Total mRNA was reverse transcribed into cDNA using the QuantiTect Reverse Transcription kit (Qiagen, Hilden, Germany). mRNA expression was quantified by real-time quantitative PCR using the QuantiTect SYBR Green PCR kit (Qiagen) and SDS7000 sequence-detection system (Applied Biosystems, Foster City, CA, USA). Primers were designed to detect carnitine palmitoyl transferase 1A (cpt1a, 12894), PR domain containing 16 (PRDM16, 70673), peroxisome proliferative activated receptor, gamma, coactivator 1 alpha (pgc1a, 19017), cell death inducing DFFA-like effector a (cidea, 12683), sirtuin 1 (sirt1. 93759), CCAAT enhancer binding protein beta (c/ebpb, 12608), and peroxisome proliferator activated receptor alpha (ppara, 19013). Amplification was performed as previously described. 20
Microarray analysis
To determine expression changes between models, mRNA was utilized for microarray analysis (Affymetrix Mouse Gene 2.0 ST Array; Affymetrix, Santa Clara, CA, USA). All steps of the arrays were performed on a GeneChip Fluidics 450 workstation (Affymetrix) and GeneChip scanner 3000 7G, according to the manufacturer's instruction. We uploaded the raw files on the Gene Expression Omnibus database (GSE157822). Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway mapping tool was used to analyze the transcriptional biological pathways related to H-FMG. Differentially expressed genes were set based on the following criteria: fold change ≥1.5 and P value <.05.
Statistical analysis
Values are represented as the mean standard error or standard deviation. We used the SPSS software version 11.0 (SPSS, Inc., Chicago, IL, USA) for statistical analysis. The statistical differences between the ND and HFD results were calculated by the Student's t-test. The significant differences among the HFD groups were analyzed using one-way analysis of variance followed by Tukey's multiple-range with P < .05.
Results
Fermented mixed grain supplementation reduces body fat mass, body weight, and plasma lipid levels
At the initial time point, in comparison with body weight, there was no significant difference among any of the groups. In contrast, the H-FMG supplement significantly lowered the final body weight compared with the HFD group (Fig. 1A). In comparison with food and energy intake (Fig. B), there was no significant difference among the HFD groups. The HFD induced an increase in adipose tissue weight, whereas H-FMG supplementation significantly decreased perirenal, retroperitoneal, mesenteric, and subcutaneous WAT weight compared with the HFD group. Consistent with our results, lipid formation in the epididymal WAT (eWAT) indicated that the H-FMG group had smaller eWAT adipocytes than the HFD group (Fig. D). Furthermore, there was excessive accumulation of fibrosis in eWAT of the HFD group than in the ND group. Besides, H-FMG supplementation lowered the accumulation of fibrosis in comparison with the HFD group.
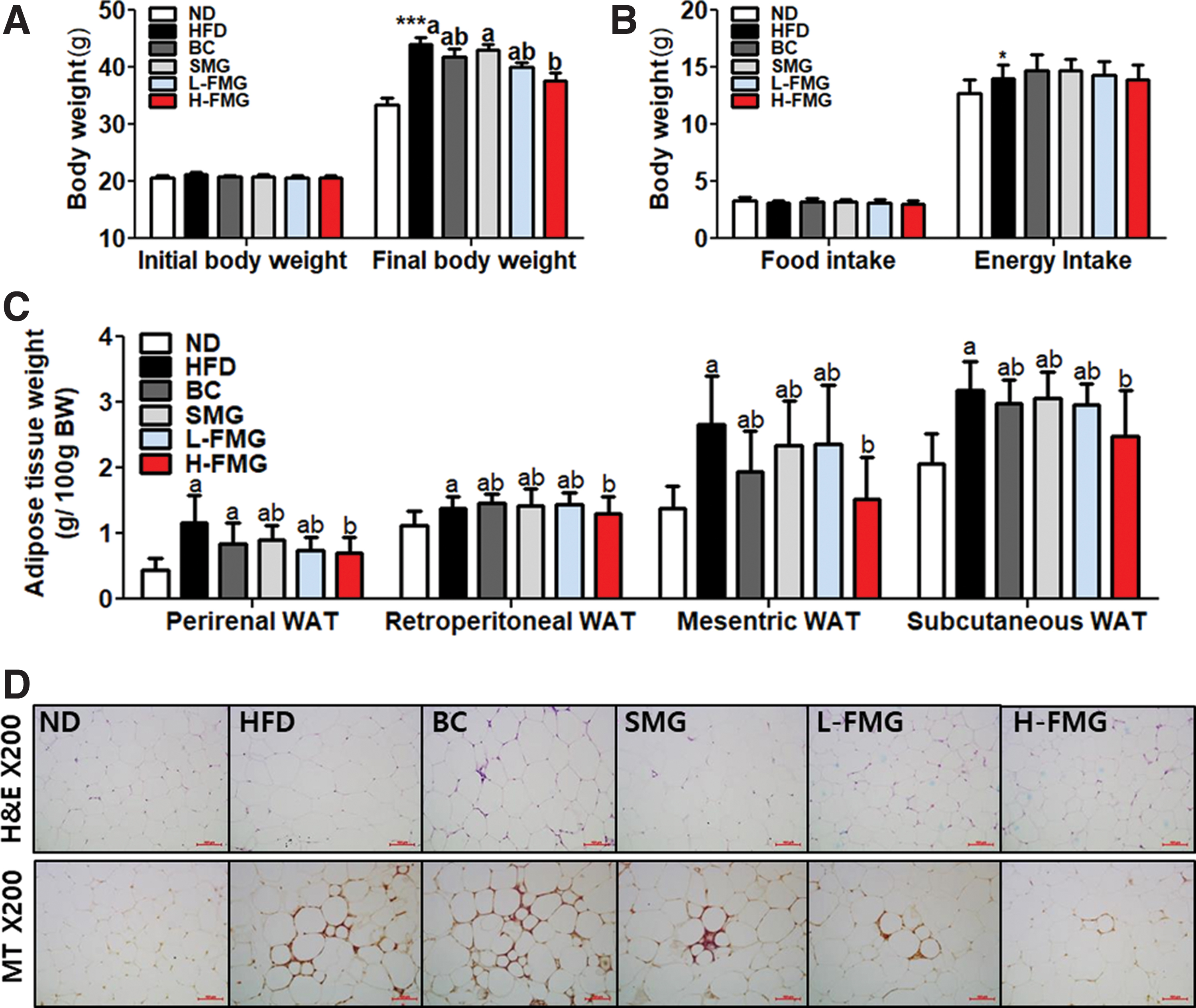
Dose-dependent effect of FMG food for 12 weeks on
Plasma lipid contents are given in Table 2. The HFD group demonstrated significantly increased levels of TC, FFA, HDL-C, non-HDL-C, apoA-1, and low-density lipoprotein (LDL) cholesterol compared with those of the ND group. However, low-dose FMG markedly lowered the concentrations of non-HDL-C, apoA-1, and LDL. H-FMG greatly lowered the concentrations of TC, non-HDL-C, apoA-1, and LDL.
Dose-Dependent Effect of Fermented Mixed Grain Food for 12 Weeks on Plasma Lipid Profiles in Diet-Induced Obese Mice
Data are mean ± SD.
ND (normal diet, low-fat diet (LFD) containing 10 kcal% fat); HFD (HFD containing 40 kcal% fat); BC (HFD+Bacillus Coagulans 0.3% (wt/wt); SMG (steamed mixed grain, HFD+SMG 0.3% wt/wt); L-FMG (low-dose fermented mixed grain, HFD +0.3% (wt/wt); H-FMG (high-dose fermented mixed grain, HFD+fermented mixed grain 0.9% (wt/wt).
Mean values with different superscript letters are significantly different among HFD-fed groups (P < .05); mean values are significantly different for ND from those of HFD, * P < .05, ** P < .01.
Apo A-I, apolipoprotein A-I; Apo B, apolipoprotein B; FFA, free fatty acid; HDL-C, high-density lipoprotein cholesterol; LDL-C, low-density lipoprotein cholesterol; SD, standard deviation; TC, total cholesterol; TG, triglyceride.
Table 3 gives the enzyme activities associated with lipid metabolism. The H-FMG supplement induced increases in β-oxidation and decreased FAS activity in comparison with HFD-fed mice.
Effect of Consuming Fermented Mixed Grain Food for 12 Weeks on Lipid Metabolism Regulating Enzyme Activity in Diet-Induced Obese Mice
Data are mean ± SD.
Mean values with different superscript letters are significantly different among HFD-fed groups (P < .05); Mean ND values are significantly different from those for HFD, ** P < 0.01. ND (normal diet, AIN-93); HFD (high-fat diet, AIN-93 containing 40 kcal% fat); BC (HFD+Bacillus Coagulans 0.3% (wt/wt); SMG (HFD +0.3% steamed mixed grain, wt/wt); L-FMG (HFD +0.3% fermented mixed grain (wt/wt); H-FMG (HFD +0.9% fermented mixed grain (wt/wt).
FAS, fatty acid synthase; G6PD, glucose-6-phosphate dehydrogenase; ME, malic enzyme; eWAT, epididymal white adipose tissue.
Fermented mixed grain supplementation increases energy expenditure and its related mRNA expression
H-FMG supplement significantly increased in carbon dioxide production (VCO2) and energy expenditure (EE) per day compared with the HFD and SMG groups. The H-FMG group demonstrated significantly upregulated mRNA expression levels of pgc1a and sirt1, factors related to fatty acid oxidation, in eWAT compared with the HFD group (Fig. 2B). We analyzed the expression of thermogenic genes in eWAT, as well as mRNA expression of prdm16, cidea, and c/ebpb in the H-FMG group compared with the HFD group.
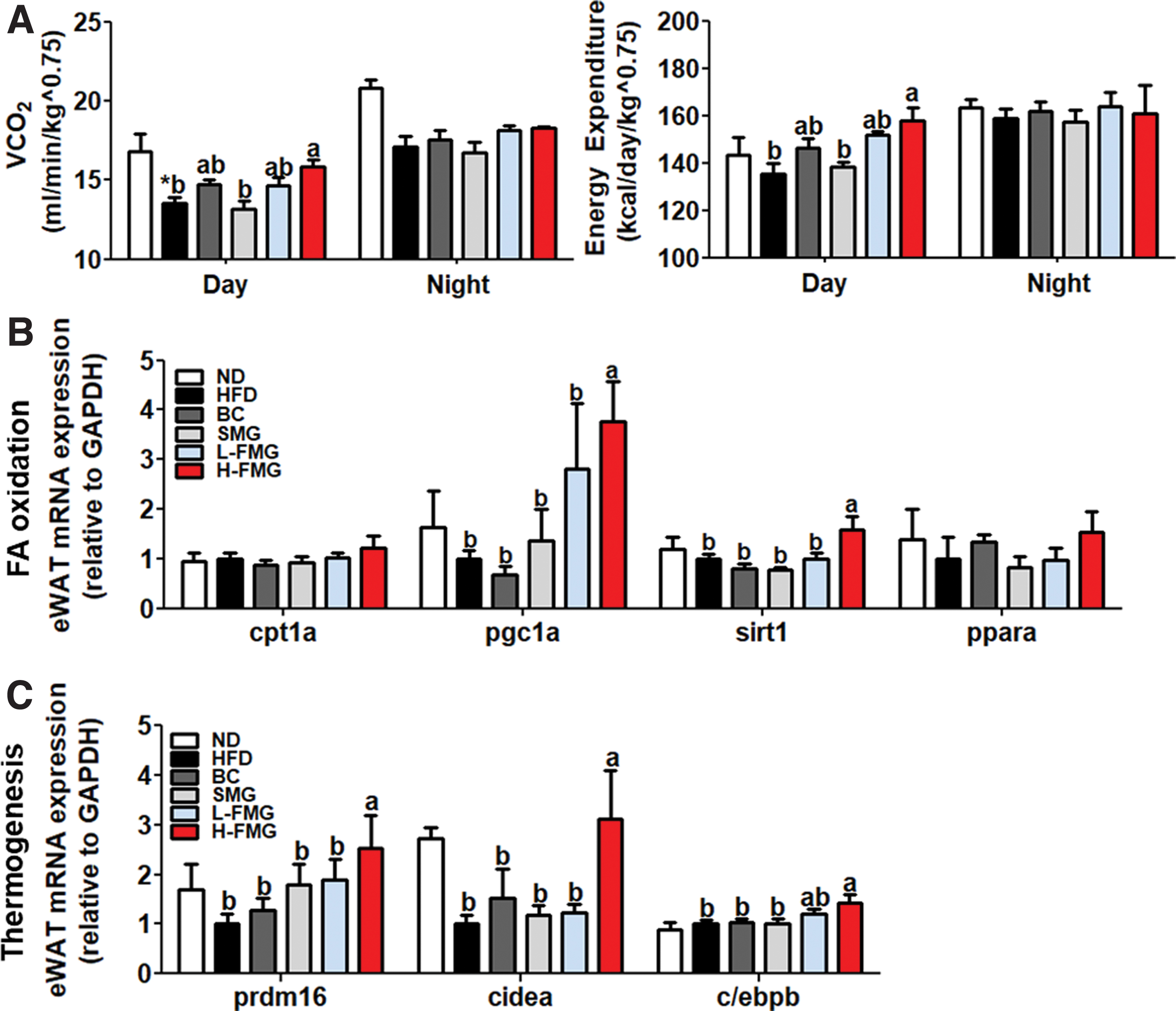
Dose-dependent effect of FMG food for 12 weeks on
Fermented mixed grain supplementation improves the impaired glucose metabolism
The FBG in H-FMG supplement was significantly lower than the HFD group from 3rd week to 12th week (Fig. 3A). The intraperitoneal glucose tolerance test and area under the curve results indicate that FMG improved glucose intolerance (Fig. 3B). Even if there was no significant difference in plasma insulin and homeostatic model assessment-insulin resistance (HOMA-IR) (Fig. 3C), hepatic PEPCK activity was significantly decreased by H-FMG supplement in diet induced obese (DIO) mice.
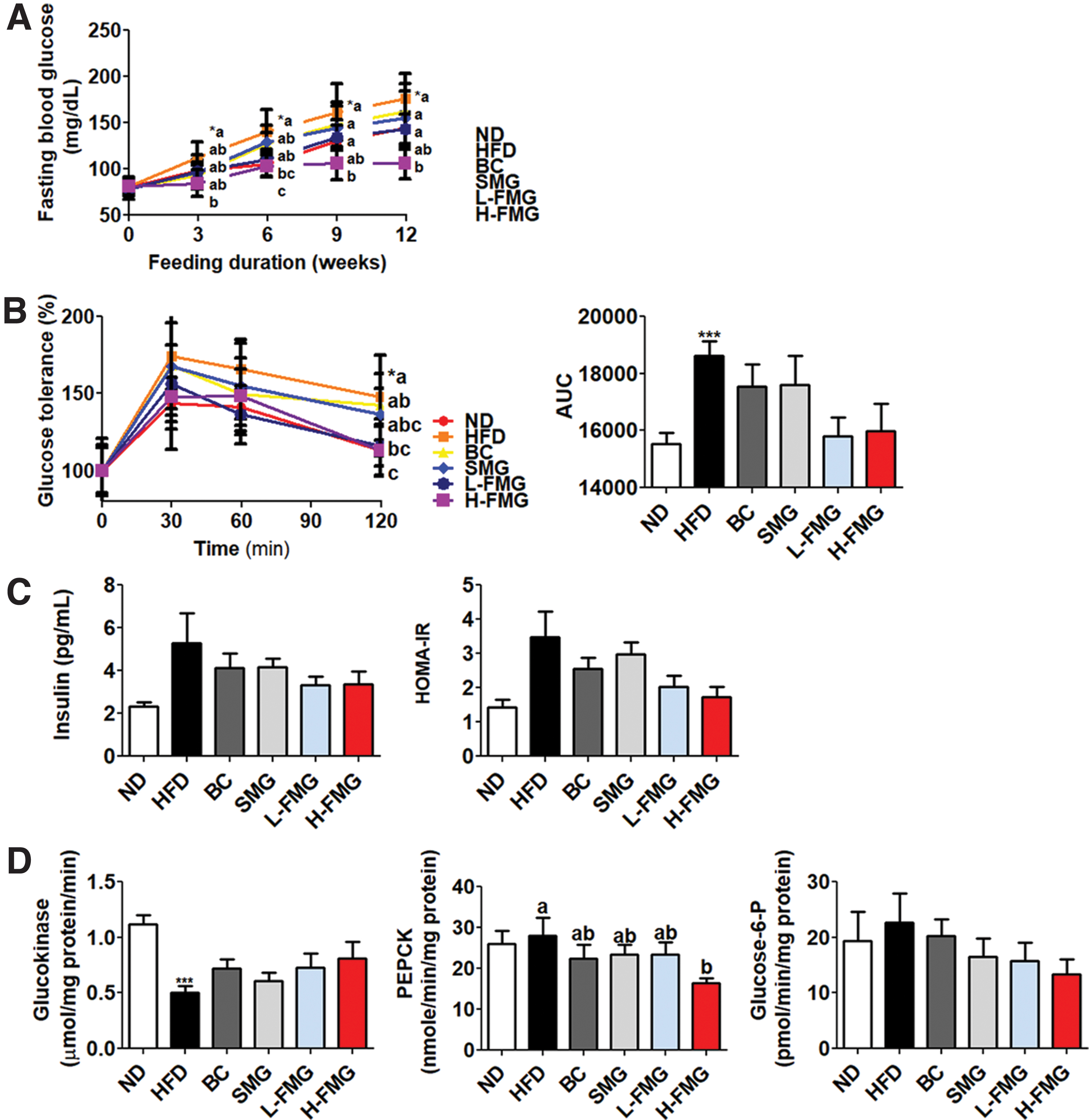
Dose-dependent effect of FMG food for 12 weeks on
Fermented mixed grain supplementation improves the hepatic steatosis
Although there was no significant difference in liver weight, hepatic TG and hepatic C were markedly lowered compared with the HFD groups (Fig. 4A). Consistent with these findings, FMG supplementation decreased the eWAT adipocyte size and lipid droplets and improved hepatic fibrosis (Fig. 4B). Moreover, FAS and ME activities in the H-FMG group were significantly decreased with the FMG supplement (Table 3).
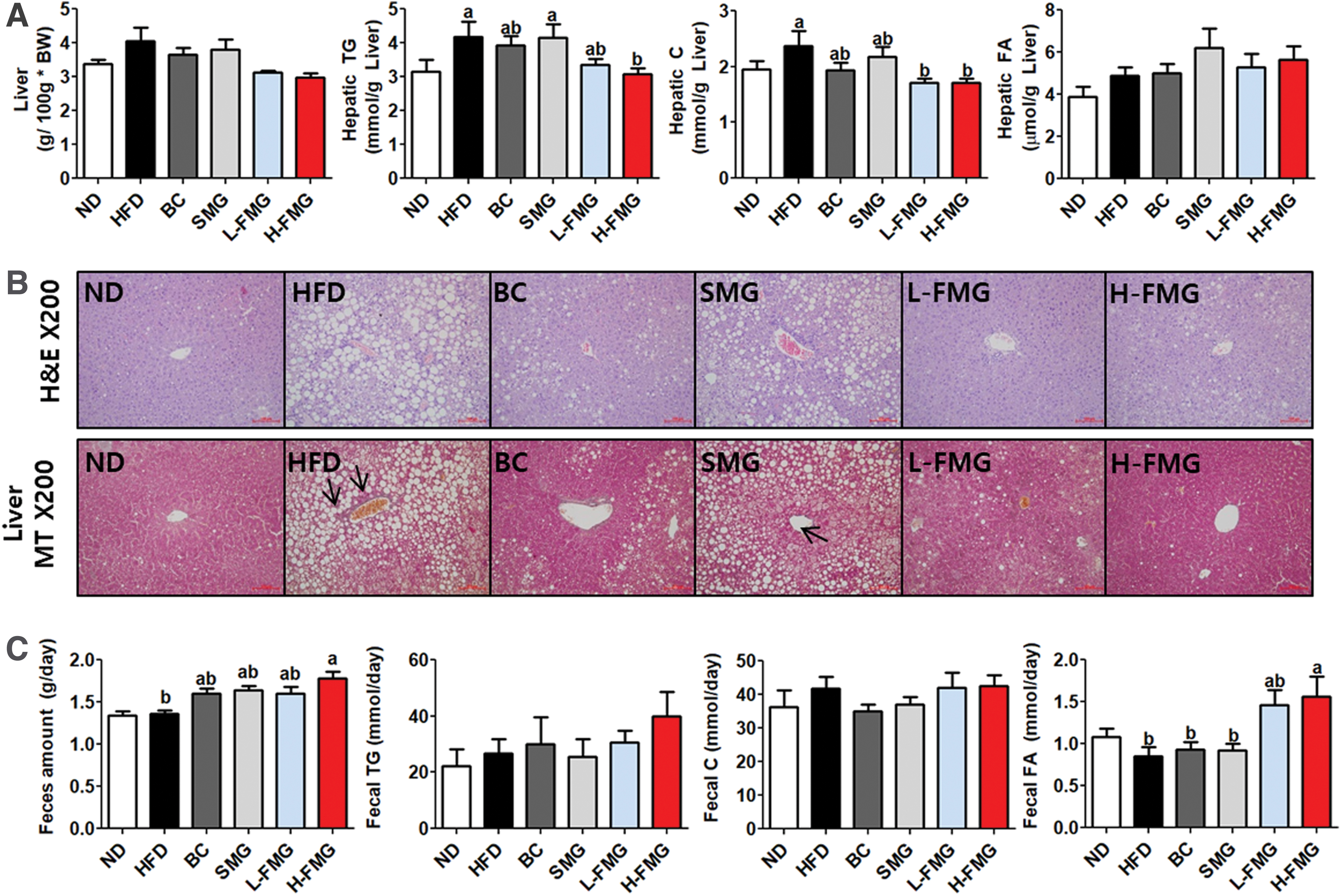
Dose-dependent effect of FMG food for 12 weeks on
The fecal lipid contents analysis (Fig. 3C) revealed that FMG supplement significantly increased the amount of feces as well as the fecal FA levels compared with the HFD group.
FMG supplementation reduces plasma adipokine and inflammatory cytokine levels
Plasma leptin concentrations were markedly decreased whereas adiponectin concentrations were dramatically increased in the H-FMG group compared with those in the HFD group (Fig. 5A, B). H-FMG supplement dramatically decreased the keratinocyte chemoattractant (KC), interleukin (IL)-1β, IL-6, tumor necrosis factor-α (TNF-α), and MCP-1 concentration in comparison with the those in the HFD group (Fig. 5A, B).
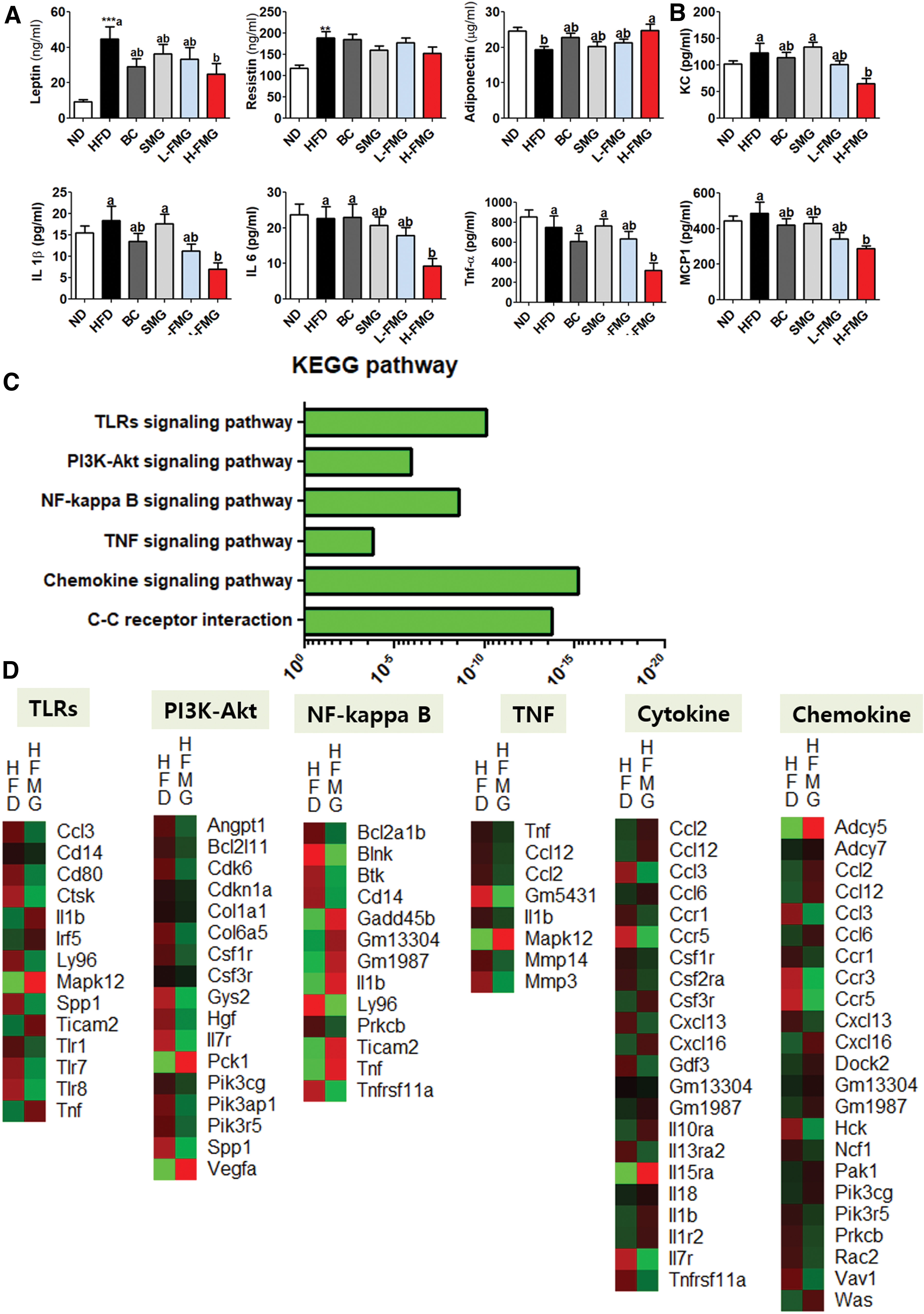
Dose-dependent effect of FMG food for 12 weeks on plasma
The transcriptomic profile of eWAT was investigated through microarray analysis. The KEGG mapper analysis is shown in Figure 5C. H-FMG supplement significantly decreased the mRNA expression related to inflammatory response.
Discussion
We investigated the dose-dependent antiobesity effects of FMG to regulate lipid and glucose metabolism in DIO mice along with its impact on overall metabolic regulation.
A positive imbalance is characterized by excess energy intake, decreased EE, and eventual obesity, which induces metabolic disorders and chronic inflammation. 21,22 Previous studies have shown that digestive enzyme inhibitors have been reported to improve obesity by interfering with the absorption of dietary carbohydrates and lipids. 17,18,23,24 However, we show that dietary intake of digestive enzymes alleviates obesity. H-FMG supplementation significantly decreased the overall body fat weight (Fig. 1A, C), without any change in food and energy intake (Fig. 1B). These results suggest that FMG reduced body fat weight through metabolic regulation, without dietary intake reduction. Consistent with these results, we determined that EE and VCO2 in the H-FMG group were significantly higher than those in the HFD group (Fig. 2A). To support these results, mRNA expression related to fatty acid oxidation (CPT1a, PGC1a, SIRT1, and PPARa) and thermogenesis (PRDM, Cidea, and C/EBPb) was markedly increased in the H-FMG group (Fig. 2B, C). H-FMG supplement significantly induced increased β-oxidation activity and alsodecreased FAS activity significantly in DIO mice. Taken together, H-FMG might increase EE through the regulation of genes associated with the fatty acid oxidation and thermogenesis in eWAT.
The physiological mechanisms associated with HFD-induced obesity are low satiety responses, high efficiency of stored dietary fats in the body, and high intake of HFD due to changes in hormones involved in energy balance. These include HFD-induced hyperleptinemia and hyperinsulinemia with leptin and insulin resistance, and decreased inhibition of ghrelin release due to HFD. 25 –27 In our study, plasma leptin concentrations in the H-FMG group were drastically lowered, whereas plasma adiponectin concentration significantly increased in comparison with those in the HFD group (Fig. 5A). Moreover, obesity is a condition in which adipose tissue accumulates excessively. Adipose tissue, a metabolically dynamic endocrine organ, secretes various cytokines and chemokines. 2,22,28 As a risk factor, obesity turns the body into a proinflammatory state by increasing inflammatory cytokines, IL-6 and TNF-α, and decreased adiponectin that has anti-inflammatory properties. 29,30 TLRs are a family of pattern recognition receptors whose molecular compositions are preserved from various microorganisms. 31 After ligand recognition, TLRs select signal adapters to start proinflammatory signaling steps leading to the activation of several transcription factor families. 32 TLRs activate the PI3K/AKT and NF-κB signaling pathways, leading to high secretion of inflammatory cytokines. In this study, transcriptome profiling showed that H-FMG supplementation significantly downregulated the expression of inflammation-linked eWAT genes compared with their expression in the HFD group. In addition, KC, IL-1β, IL-6, TNF-α, and MCP-1 concentrations were significantly decreased with body fat weight reduction in H-FMG compared with those in the HFD group (Fig. 5B). Hepatic and WAT fibrosis, caused by chronic inflammation, were markedly improved with H-FMG supplementation (Fig. 1D, 3B).
Obesity, which can contribute to NAFLD and hepatic steatosis, is the highest risk element for the development of T2DM. 33,34 NAFLD was found in 50% of patients with T2DM who had normal aminotransferase levels. 35 The correlation between hepatic fat and insulin resistance is even relevant from the perspective of managing hyperglycemia in patients with T2DM. 36 First, the degree of hepatic fat accumulation is an important factor for determining the amount of insulin required to decrease glucose levels in T2DM patients. 37 The insulin requirement is closely related to hepatic fat content, it is expected that patients with NAFLD need more insulin to improve glycemic control than those without NAFLD. 38 Second, a decreased hepatic fat content through dieting has been shown to decrease insulin resistance. 39 In our study, although there was no significant difference in liver weight among the HFD groups, hepatic fatty acid concentration, hepatic TG, and hepatic cholesterol concentration were significantly decreased. Also, hepatic FAS and ME, which are related to fatty acid synthesis, were significantly decreased. Glucose metabolism analysis showed that H-FMG supplementation-induced FBG was significantly decreased, which improved the insulin resistance with markedly decreased PEPCK activity.
Obesity and T2DM are well known to accompany chronic low-grade inflammation along with an oxidative stress environment seen in the WAT expansion. 8,40 The TLR4 signaling pathway is associated with obesity and T2DM caused by chronic inflammation and insulin resistance. 41,42 In obese/T2DM patients, TLR2 and TLR4 have been considered as sensors of lipopolysaccharide and FFA. 43 TLR10 expression was regulated by ROS-mediated oxidative stress by not only ER stress but also NF-κB/MAPK signaling. Therefore, TLR10 could be an immune biomarker for inflammation. 44 In patients with T2DM, β cell dysfunction may be associated with a strong association between TLRs and inflammation. 45 TLR2 expression was increased in obese/T2DM patients and related to an inflammatory response, as assessed by increased serum levels of IL-18. 46 A high-glucose level could lead to increased TLR2 and TLR4 expression in retinal ganglion cells through increased release of proinflammatory factors in diabetic retinopathy. 47 In our study, although there were no significant differences in insulin levels and HOMA-IR, H-FMG supplement significantly decreased fasting blood glucose levels with decreases in plasma inflammatory cytokines and associated mRNA expression.
The limitations of this study are as follows. We found that FMG supplement improved obesity and its complications. However, it is still unclear whether the effect is due to digestive enzymes or FMG itself. Accordingly, another study is ongoing in which we will elucidate the active ingredient of FMG.
In conclusion, we demonstrated that FMG supplement can reduce body weight and body fat mass with improvement of the metabolic status in DIO mice through the TLR signaling pathway. Furthermore, this study suggested that the properties of FMG supplementation were dose dependent.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This research was supported by Basic Science Research Program (2020R1I1A1A01067942, 2020R1I1A3074694), through the National Research Foundation of Korea (NRF) funded by the ministry of education.