
Abstract
We investigated the effects of GT collagen (Geltech low-molecular-weight fish collagen, FC) on skin moisturization in ultraviolet B (UVB)-irradiated HaCaT cells and SKH-I hairless mice. In vitro, we measured the expression of mRNA genes and proteins related to the skin moisturizing mechanism, hyaluronic acid concentrations, and sphingomyelin concentrations. As a result, FC increased the expression of LCB1, DEGS1, elastin, UGTrel7, and GlcNAc mRNA in UVB-irradiated HaCaT cells. Also, hyaluronic acid level, sphingomyelin level, and protein expressions of hyaluronan synthase (HAS)2 and CerS4 were increased compared to those in the UVB-irradiated control group. In vivo, we measured skin hydration through the expression of mRNA genes and proteins related to the skin moisturizing mechanism and found that the protein expression of HAS2 and CerS4 was increased in the groups taking FC. Moreover, FC intake increased the expression of LCB1, DEGS1, fibrilin-1, UGTrel8, and GlcNAc mRNA in UVB-irradiated SKH-I hairless mice. These results suggest that FC can be utilized to develop products aimed at improving skin moisturization.
Introduction
Exogenous skin aging caused by the environment, such as the recent increase in the ultraviolet (UV) index, is a subject of enormous interest, and research on foods and cosmetics as natural materials is being conducted to address this. 1 Among the factors of skin aging, photoaging due to chronic exposure to UV is common and causes symptoms such as decreases in skin moisture, loss of elasticity, formation of wrinkles, and pigmentation. 2
Skin is composed of the epidermis, dermis, and subcutaneous fat layer. Among them, melanocytes and keratinocytes are distributed in the epidermis, fibrous components and matrix components are included in the dermis, and collagen is present as a fiber component in 90% of the dermis. 3 When the skin's epidermis, which acts as a skin barrier by preventing the loss of moisture from the body, is damaged by UV rays, and skin moisture decreases, the tissues becomes dry, wrinkled, and rough. Therefore, it is important to replenish sufficient moisture to maintain healthy skin. 4,5
Hyaluronic acid in the stratum corneum of the skin moisturizes the skin, and it is important to maintain the level to inhibit skin aging. 6 Hyaluronic acid is distributed in connective tissue, epithelial tissue, and nervous tissue, and one-third of the total is degraded and synthesized daily. Because of this, the concentration must be maintained by hyaluronan synthase (HAS) activity. 7 It has been reported that UV irradiation induces skin inflammation and inhibits hyaluronic acid synthesis. 8 In addition, when the skin is exposed continuously to UV rays, reactive oxygen species are generated in the epidermis along with damage to the skin barrier, increasing the production of proinflammatory cytokines and causing an inflammatory reaction. As a result, skin connective tissues such as collagen, elastin, and fibrillin in the dermal layer are reduced, causing wrinkles and moisture loss. Therefore, both the epidermis and the dermis play a role in moisturization of the skin. 9
Recently, it has been shown that ingestion of collagen peptides isolated from fish scales improves moisturization and inhibits wrinkles in the skin. 10 –12 We focused on the evaluation of the efficacy of GT collagen (Geltech low-molecular fish collagen, FC), which has a large low-molecular-weight fraction and an average molecular weight less than 1000 Da.
Therefore, in our study, we evaluated the efficacy of GT collagen for skin moisturization in ultraviolet B (UVB)-irradiated HaCaT cells and SKH-I hairless mice, and assessed the feasibility of its utilization in products for skin health.
Materials and Methods
Preparation of materials
The GT collagen was supplied by Geltech (Busan, Korea) as purified fish scale collagen originating from tilapia (Oreochromis genus) gelatin. GT collagen purification was accomplished through hydrolysis followed by filtration, sterilization, drying, and sieving, as modified from a patented manufacturing process and preparation of collagen peptide (KR100647033B1). L-ascorbic acid (AA) and arbutin (Sigma-Aldrich, St. Louis, MO, USA) were used as the positive controls.
Liquid chromatography-mass spectrometry analysis
FC (100 mg) was weighted and 50 mL of deionized water was added. Zorbax Eclipse plus C18 column was used. The column temperature was 40°C, with flow rate of 0.3 mL/min, and injection volume of 3 μL. The mobile phase consisted of A (0.1% formic acid/H2O = 0.1/100 v/v) and B (0.1% formic Acid/Acetonitrile = 0.1/100 v/v). The gradient conditions were as follows: 0–1.0 min (2% B), 1–4.0 min (70% B), 4.0–6.0 min (70% B), 6.0–6.1 min (95% B), 6.1–10.0 min (95% B), and 10.0–10.1 min (2% B), 10.1–15.0 min (2% B). Mass spectrometry (MS) analysis was performed with the TSQ Altis™ (Thermo Fisher Scientific, Waltham, MA, USA).
Cell viability
Cell viability was analyzed by using the MTT assay method. HaCaT cells were seeded at a density of 1 × 104 cells/well into a 96-well plate. After the cells were stabilized at the bottom of the plate, FC was added to each well at a dose of 0–1000 μg/mL. After incubation for 24 h, 20 μL of MTT solution prepared from thiazolyl blue tetrazolium bromide (Sigma-Aldrich) was added to each well for 4 h. After removing the supernatant, 100 μL of dimethyl sulfoxide was added to each well and the optical density was measured at 560 nm (enzyme-linked immunosorbent assay [ELISA] reader; Bio-Rad Laboratories, Hercules, CA, USA).
Cell and animal treatment
HaCaT cells, the keratinocytes of human skin used in this experiment, were received from Professor Hwang of the College of Life Sciences at Kyung Hee University. HaCaT cells seeded for each experiment were washed, and exposed to 50 mJ/cm2 of UV ray using a UVB lamp (5 Sankyo Denki G5T5 lamps; Sankyo Denki Co., Yokohama, Japan) and treated with AA (100 μg/mL) and FC (50, 100, 200, and 400 μg/mL).
This study was approved by the Institutional Animal Care and Use Review Committee of Kyung Hee University (KHGASP-20-346). Male SKH-I hairless mice (5 weeks old, 20 ± 2 g) were purchased from Saeronbio, Inc., (Uiwang, Korea). The housing condition was maintained at 23°C ± 2°C and 50% ± 5% relative humidity with a 12-h light/12-h dark cycle. A total of 56 mice were assigned randomly to the following 7 groups with 8 animals per group: normal control (NC), UVB-irradiated control (UVB-C), L-ascorbic acid 200 mg/kg b.w. (AA, positive control 1), arbutin 200 mg/kg b.w. (arbutin, positive control 2), and GT collagen (FC) 200, 400, and 600 mg/kg b.w. All experimental diets were based on the AIN93G diet and oral administration was conducted. We used a UVB lamp (Sankyo Denki Co.) for repeated UVB exposure of mice under the same conditions as in our previous study, which induced skin dryness. 13 The animal experiment was carried out for 8 weeks, following that, skin tissues of the mice were collected for analysis.
Real-time polymerase chain reaction
HaCaT cells were seeded with 5 × 105 cells/well in six-well-plates, animal skin tissues were lysed, and the total RNA was extracted using an RNeasy extraction kit (Qiagen, Gaithersburg, MD, USA). Subsequently, all processes of real-time polymerase chain reaction were performed with our previous protocol. 14 The gene sequences were designed using an applied biosystem program (Primer express; Thermo Fisher Scientific) (Table 1). All data were calculated through the Bio-Rad CFX manager 3.1 (Bio-Rad).
Primer Sequences Used in Real-Time Polymerase Chain Reaction-Based Quantification of mRNA
UGTrel, UDP-glucuronic acid; GlcNAc, UDP-N-acetylglucosamine; LCB1 (SPT), long chain base 1 (Serine palmitoyltransferase); DEGS1, delta 4-desaturase, sphingolipid 1; GAPDH, glyceraldehyde-3-phosphate dehydrogenase.
Western blotting
HaCaT cells seeded with 5 × 105 cells/well in a six-well-plate and animal skin tissues were homogenized using the CelLytic™ MT Cell Lysis Reagent (Sigma-Aldrich) with Halt™ Protease & Phosphatase inhibitor Cocktail (Thermo Fisher Scientific). Subsequently, protein quantification, sample loading, and protein transfer were performed as in our previous method. 15 Primary antibodies (HAS2, CerS4, and ß-actin) and horseradish peroxidase-conjugated secondary antibodies were purchased from Cell Signaling Technology and applied to the experiment. The visualization and analysis steps were conducted with the conventional method. 15
Enzyme-linked immunosorbent assay
HaCaT cells seeded with 1 × 104 cells/well in a 96-well plate were homogenized using each lysis buffer of the Hyaluronic Acid ELISA Kit (Biovision, Inc., CA, USA) and the Sphingomyelin Assay Kit (Abcam, Cambridge, UK) according to the manufacturer's protocol. The optical density was measured at 540 and 595 nm, respectively (ELISA reader; Bio-Rad).
Measurement of skin hydration
Skin hydration was tested with a Howskin instrument (Innoinsight, Inc., Yongin, Korea) immediately before sacrifice of the animal. The moisture value was measured in such a way that the electronic terminal of the device was read from the back of the mice and transmitted to an application on a mobile phone.
Statistical analysis
All experimental data are shown as means ± standard deviations. We analyzed the significance of values with Duncan's multiple-range test after conducting a one-way analysis of variance using the SPSS statistical program (SPSS PASW Statistic 23.0; SPSS, Inc., Chicago, IL, USA). Statistical significance was noted at P < .05.
Results
Liquid chromatography-mass spectrometry analysis of FC
The liquid chromatography-mass spectrometry analysis of FC revealed a peak matching those of the commercial standard VGPPGPAG with retention times of ∼3.79 min (Fig. 1). The FC contained VGPPGPAG from a minimum of 0.93 g/g (Fig. 1.).

LC-MS of VGPPGPAG in the FC.
Effects of FC on mRNA expressions related to skin moisturization in UVB-irradiated HaCaT cells
The mRNA expression of LCB1 was significantly reduced in the UVB-C group (63.7%) compared to the NC group. Compared with the UVB-C group, the AA (131.7%) and FC (50.6% minimum to 114.2% maximum) groups showed significantly increased mRNA expression of LCB1 (P < .05; Fig. 2A). The mRNA expression of DEGS1 was significantly reduced in the UVB-C group (53.3%) compared to the NC group. Compared with the UVB-C group, the AA (77.7%) and FC (49.7% minimum to 99.7% maximum) groups showed significantly increased mRNA expression of DEGS1 (P < .05; Fig. 2B). The mRNA expression of elastin was significantly reduced in the UVB-C group (59.9%) compared to the NC group. Compared with the UVB-C group, the AA (85.0%) and FC (37.4% minimum to 101.2% maximum) groups showed significantly increased mRNA expression of elastin (P < .05; Fig. 2C). The mRNA expression of UGTrel7 was significantly reduced in the UVB-C group (58.4%) compared to the NC group. Compared with the UVB-C group, the AA (87.5%) and FC (25.0% minimum to 80.8% maximum) groups showed significantly increased mRNA expression of UGTrel7 (P < .05; Fig. 2D). The mRNA expression of GlcNAc was significantly reduced in the UVB-C group (65.1%) compared to the NC group. Compared with the UVB-C group, the AA (139.0%) and FC (50.0% minimum to 117.2% maximum) groups showed significantly increased mRNA expression of GlcNAc (P < .05; Fig. 2E).
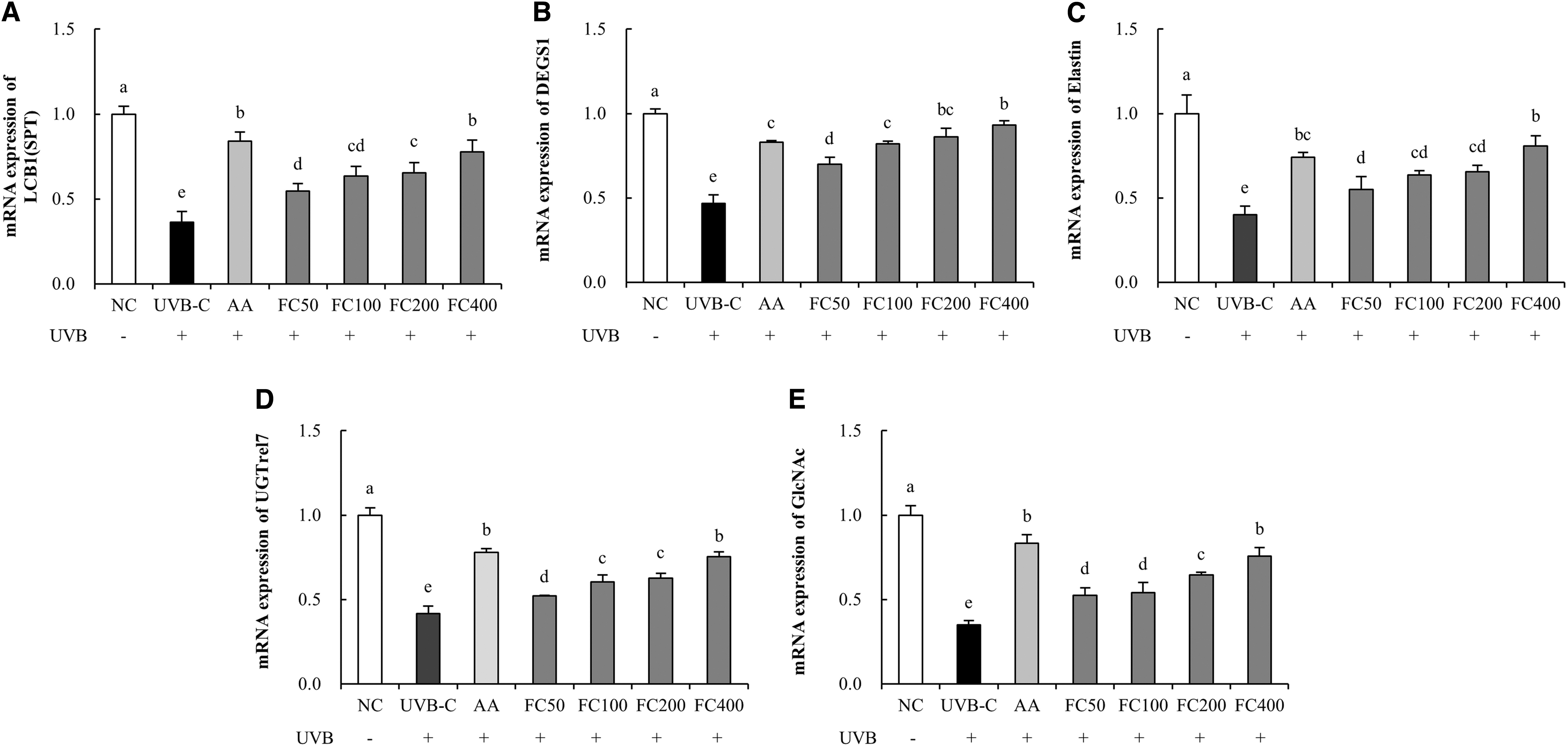
Effects of FC on the expression of mRNAs related to skin moisturization in UVB-irradiated HaCaT cells.
Effects of FC on hyaluronic acid and sphingomyelin levels in UVB-irradiated HaCaT cells
The concentration of hyaluronic acid was significantly reduced in the UVB-C group (27.8%) compared to the NC group and significantly increased in the AA group (23.8%) compared to the UVB-C group. Compared with the UVB-C group, the FC group (16.9% minimum to 38.4% maximum) showed significantly increased hyaluronic acid concentrations in a dose-dependent manner (P < .05; Fig. 3A). The concentration of sphingomyelin was significantly reduced in the UVB-C group (37.6%) compared to the NC group and significantly increased in the AA group (38.4%) compared to the UVB-C group. Compared with the UVB-C group, the FC group (24.8% minimum to 46.5% maximum) showed significantly increased sphingomyelin concentrations in a dose-dependent manner (P < .05; Fig. 3B).
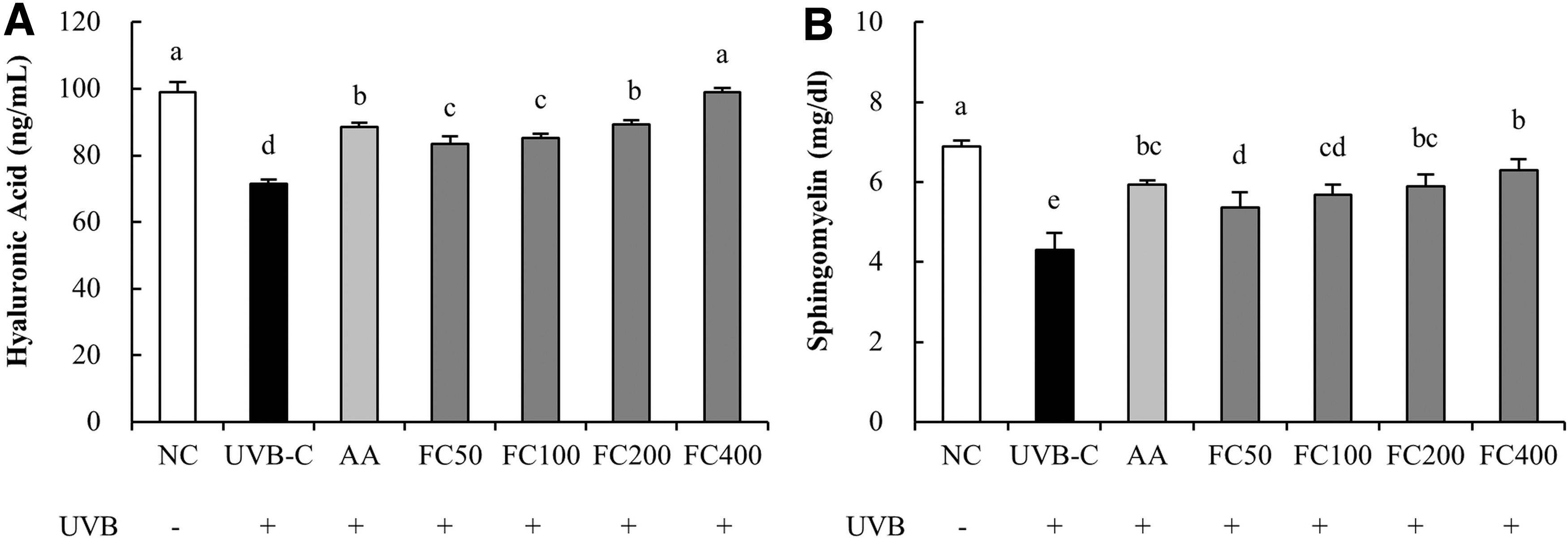
Effects of FC on levels of hyaluronic acid and sphingomyelin in UVB-irradiated HaCaT cells.
Effects of FC on protein expression related to skin moisturization in UVB-irradiated HaCaT cells
The protein expression of HAS2 was significantly reduced in the UVB-C group (53.2%) compared to the NC group and significantly increased in the AA group (89.2%) compared to the UVB-C group. Compared with the UVB-C group, the FC groups (29.9% minimum to 73.3% maximum) showed significantly increased protein expression of HAS2 (P < .05; Fig. 4A). The protein expression of CerS4 was significantly reduced in the UVB-C group (37.2%) compared to the NC group and significantly increased in the AA group (34.9%) compared to the UVB-C group. Compared with the UVB-C group, the FC groups (6.4% minimum to 57.2% maximum) showed significantly increased protein expression of CerS4 (P < .05; Fig. 4B).
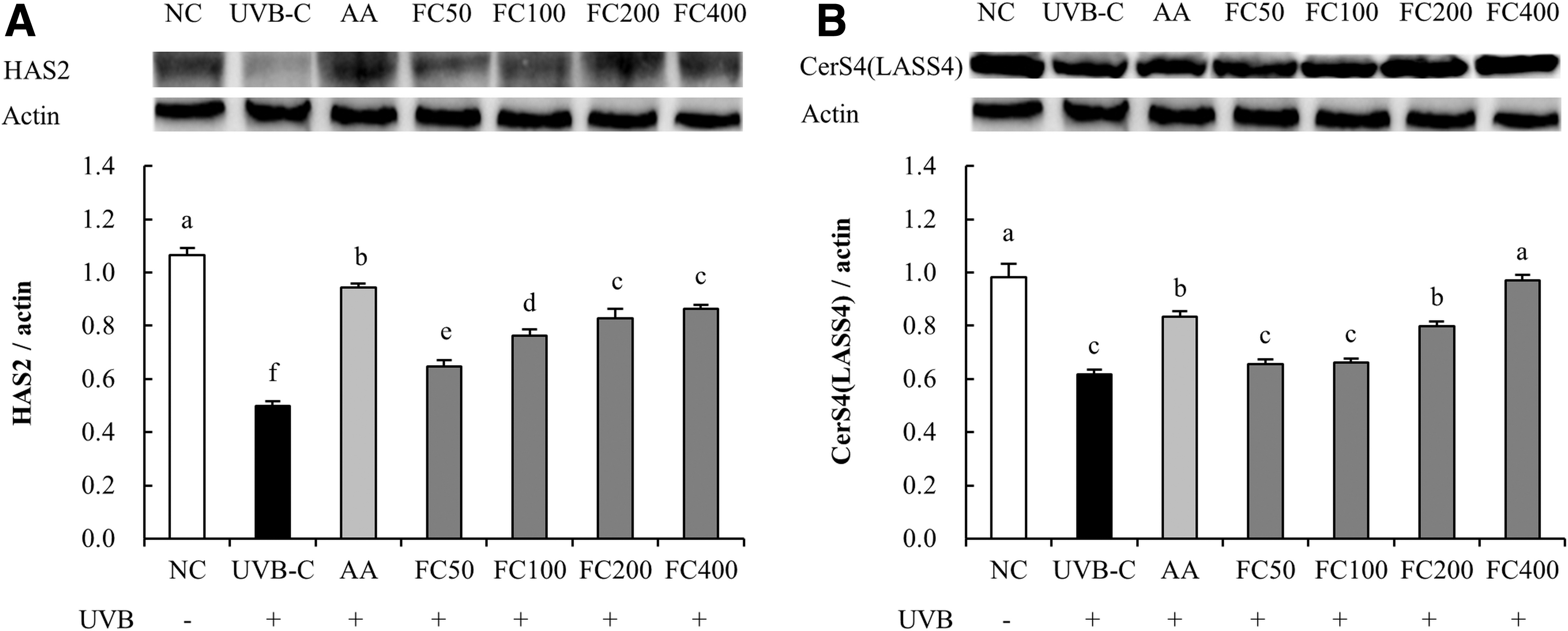
Effects of FC on protein expression related to skin moisturization in UVB-irradiated HaCaT cells.
Effects of FC on skin hydration in UVB-irradiated SKH-I hairless mice
The water content of skin was significantly reduced in the UVB-C group (37.3%) compared to the NC group and significantly increased in the AA and arbutin groups (37.2% and 15.1%, respectively) compared to that of the UVB-C group. Compared with the C group, the FC groups showed significant increase in water content of the skin in a dose-dependent manner (FC200: 10.9%, FC400: 24.3%, and FC600: 28.5%; P < .05; Fig. 5F).
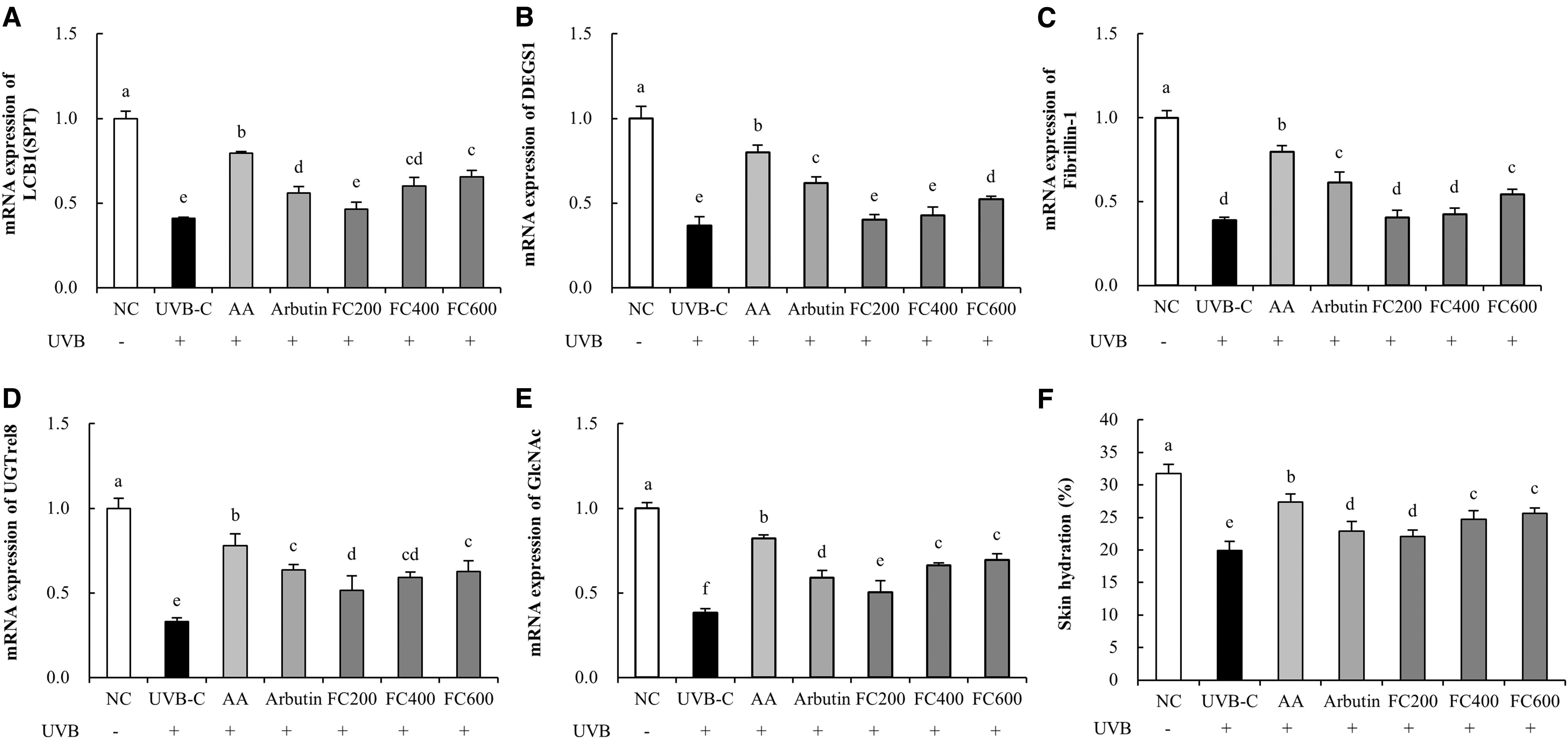
Effects of FC on the expression of mRNAs related to skin moisturization and hydration of skin from the back of UVB-irradiated SKH-I hairless mice.
Effects of FC on mRNA expressions related to skin moisturization in UVB-irradiated SKH-I hairless mice
The mRNA expression of LCB1 was significantly reduced in the UVB-C group (59.0%) compared to the NC group. Compared with the UVB-C group, the AA (94.0%), arbutin (36.8%), and FC (13.6% minimum to 59.8% maximum) groups showed significantly increased mRNA expression of LCB1 (P < .05; Fig. 5A). The mRNA expression of DEGS1 was significantly decreased in the UVB-C group (63.2%) compared to the NC group. Compared with the UVB-C group, the AA (117.0%), arbutin (68.2%), and FC (9.1% minimum to 41.9% maximum) groups showed significantly increased mRNA expression of DEGS1 (P < .05; Fig. 5B). The mRNA expression of fibrilin-1 was significantly reduced in the UVB-C group (61.1%) compared to the NC group. Compared with the UVB-C group, the AA (104.6%), arbutin (57.4%), and FC (3.9% minimum to 39.3% maximum) groups showed significantly increased mRNA expression of fibrilin-1 (P < .05; Fig. 5C). The mRNA expression of UGTrel8 was significantly decreased in the UVB-C group (66.9%) compared to the NC group. Compared with the UVB-C group, the AA (136.1%), arbutin (92.5%), and FC (55.8% minimum to 89.5% maximum) groups showed significantly increased mRNA expression of UGTrel8 (P < .05; Fig. 5D). The mRNA expression of GlcNAc was significantly decreased in the UVB-C group (61.8%) compared to the NC group. Compared with the UVB-C group, the AA (115.3%), arbutin (54.5%), and FC (32.1% minimum to 81.4% maximum) groups showed significantly increased mRNA expression of GlcNAc (P < .05; Fig. 5E).
Effects of FC on expression of proteins related to skin moisturization in UVB-irradiated SKH-I hairless mice
The protein expression of HAS2 was significantly reduced in the UVB-C group (70.8%) compared to the NC group. Compared with the UVB-C group, the AA (183.3%), arbutin (164.2%), and FC groups (111.8% minimum to 202.3% maximum) showed significantly increased protein expression of HAS2 (P < .05; Fig. 6A). The protein expression of CerS4 was significantly decreased in the UVB-C group (31.1%) compared to the NC group. Compared with the UVB-C group, the AA (45.8%), arbutin (5.4%), and FC groups (22.0% minimum to 42.6% maximum) showed significantly increased protein expression of CerS4 (P < .05; Fig. 6B).
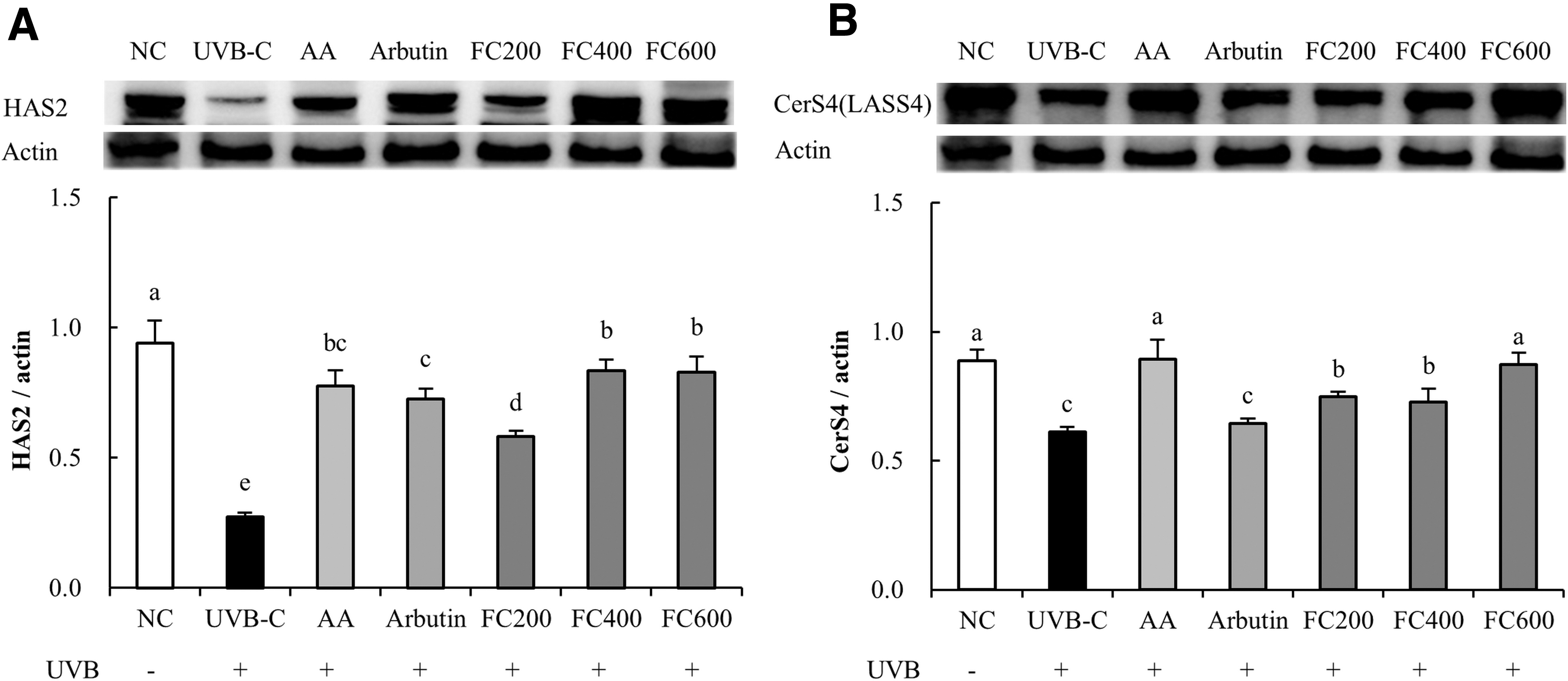
Effects of FC on protein expression related to skin moisturization in UVB-irradiated SKH-I hairless mice.
Discussion
The change in climate and destruction of the ozone layer surrounding the earth increase the amount of UV rays that reach the earth's surface. As a result, concerns about the health of human skin are increasing. 15 UV exposure reduces moisture in the stratum corneum and fibrous layers of the skin, which can lead to serious skin lesions. 16 Prolonged exposure of the skin to UV can lead to itching due to dry skin and skin cancer such as melanoma. Some steroid drugs used to treat skin damage have side effects that cause secondary pain in patients. 17 Therefore, natural treatments, including heath foods and functional cosmetics, are of increasing interest to prevent skin damage caused by UV without side effects. 1
Collagen is a support protein found in both vertebrates and invertebrates and affects the formation of skin, bones, teeth, and tendons, which account for 30% of the protein in the human body, and is concentrated in the dermal layer of the skin. 18 Therefore, as a very useful biomaterial, it is widely used in the cosmetics and food industries and is biocompatible due to its properties such as biodegradability and low antigenicity. 19 –22 Many studies on collagen extracted from the skin and connective tissues of marine vertebrates/invertebrates such as squid, jellyfish, skates, and sea cucumbers have been reported. 23 –26
Collagen peptide is a substance obtained through hydrolysis after extracting and heating high-molecular-weight collagen from fish scales, pig skin, or cattle skin to convert it into gelatin. When this peptide enters the body, it is decomposed into amino acids by enzymes, but can be resynthesized into collagen peptides by vitamin C. It has been reported that collagen peptides are effective for moisturizing skin, improving elasticity, and preventing wrinkles, for which they are applied to the development of functional cosmetics. 27 However, in recent studies, the high molecular weight of certain collagen peptides is decomposed easily in the stomach when ingested and then they cannot affect the skin. For this reason, research on low-molecular collagen peptides has shown greater efficacy compared to that of high-molecular collagen peptides in moisturizing skin and inhibiting wrinkles. 28
This study used low-molecular fish collagen derived from tilapia gelatin, and VGPPGPAG, an amino acid peptide, was detected in FC. Tilapia fish of the Oreochromis genus are distributed widely across the wild and are characterized by robust properties, fast growth rates, and salt tolerance. 29 They can be used as a collagen source and are farmed widely. 30 Studies applying collagen extracted from tilapia have been conducted. 31 One such study reported that biomimetic tilapia collagen nanofibers are effective in skin regeneration by inducing differentiation of keratinocytes and collagen synthesis in skin fibroblasts. 32 In addition, the effect of collagen derived from tilapia fish on skin moisturization is known. 33 However, there is need for studies on the effects of low-molecular-weight collagen extracted from tilapia fish on gene and protein expressions affecting ceramide content in a moisturization-related mechanism.
HaCaT is a genetically abnormal human skin cell. However, since it differentiates similar to normal keratinocytes, it is used widely in research to model skin keratinocytes. In addition, the quality of these cells is maintained in a stable genetic balance even at high passage levels. 34 Recently, marine collagen derived from the skin of Nile tilapia showed a wound healing effect in the HaCaT cell model. 35 In an in vitro study, the cell viability test result for the HaCaT cells exposed to FC revealed cell viability of 90% or greater up to a dose of 1000 μg/mL (data not shown). Moreover, FC derived from tilapia increased the gene expression of LCB1 and DEGS1, factors related to ceramide production in UV-irradiated HaCaT cells. It also increased the protein expression of CerS4, one of the various ceramide synthases involved in the synthesis of ceramides mainly expressed in skin. 36 In addition, FC increased the gene expression of UGTrel7 and GlcNAc, factors involved in the production of hyaluronic acid, and increased the protein expression of HAS2. These effects resulted in an increase in the content of sphingomyelin and hyaluronic acid in the UV-irradiated HaCaT cells.
SKH-I hairless mice have no hair, making them convenient for UV irradiation and susceptible to tumors, so they are a commonly used model for tumor formation studies. 37 Since SKH-I hairless mice are sensitive to environmental conditions, UV-induced water loss was expected. In an in vivo study, there were no significant differences in body weight and organ weight among any of the groups (Table 2). Moreover, oral intake of FC derived from tilapia increased the gene expression of LCB1 and DEGS1, which had been decreased by UV exposure, and increased the protein expression of CerS4. In addition, FC increased gene expression of UGTrel8, GlcNAc, and fibrilin-1 and protein expression of HAS2, increasing skin hydration.
Effects of FC on Body Weight and Organ Weights of SKH-1 Hairless Mice With or Without Ultraviolet B Irradiation
Values are presented as mean ± SD (n = 8), and different superscript letters indicate significance at P < .05.
NC, normal control; UVB-C, ultraviolet B-irradiated control; AA, L-ascorbic acid 200 mg/kg b.w.; Arbutin, Arbutin 200 mg/kg b.w.; FC, low-molecular fish collagen 200, 400, and 600 mg/kg b.w.
Weight gain (g/8 weeks) = final body weight (g) – initial body weight (g).
FER = weight gain (g)/total food consumption (g) × 100.
FER, food efficiency rate; NC, normal control; SD, standard deviation.
In our study, we investigated whether FC plays a role in skin moisturization in UVB-irradiated HaCaT cells and SKH-I hairless mice. Skin exposure to UV light has been reported to inhibit the synthesis of hyaluronic acid and cause an inflammatory reaction. 38 We found that FC increased gene or protein expression of LCB1, DEGS1, CerS4, UGTrel, and GlcNAc, which are skin moisturizing parameters, in cells and animal models exposed to UV light. This is meaningful in that it confirmed the indicators involved in the production of ceramide and hyaluronic acid compared with previous studies.
The results of this study verified that FC helps improve moisturization of the skin in vitro and in vivo and can be used in the development of cosmetic and nutraceutical products aimed at improving general skin moisturization. However, more studies are needed to confirm the efficacy of FC in humans.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.