
Abstract
Low-grade inflammation might be a link between obesity and obesity-associated metabolic dysfunction, including diabetes, hepatic steatosis, and other health complications. This study investigated whether the supplementation of high hydrostatic pressure extract of mulberry (Morus alba L.) leaves (HML) to obese rats could counteract obesity-related inflammation. Three-week-old male Sprague-Dawley rats were separated into three groups as follows: (a) a normal diet, (b) 45% high-fat (HF) diet, and HF diet containing 0.4% HML (c) or 0.8% HML (d) (IACUC No. 17-033). After 14 weeks of HML supplementation, adipose tissue mass, mRNA expression of adipogenic genes, such as aP2, peroxisome proliferator-activated receptor γ (PPARγ), and sterol regulatory element binding protein 1c (SREBP1c), and macrophage recruitment were significantly decreased in HF-fed obese rats. Serum concentrations of nitric oxide and mRNA levels of arginase1 (Arg1), CD11c, and inducible nitric oxide synthase (iNOS) involved in adipose tissue macrophage M1 polarization were also significantly reduced by HML. Moreover, HML alleviated the serum and hepatic lipid profiles and reduced hepatic lipogenic gene expression of acetyl-CoA carboxylase (ACC), cluster of differentiation 36 (CD36), CPT1, fatty acid synthase (FAS), stearoyl-CoA desaturase (SCD1), and SREBP1c, and inflammation-associated genes, including IL1β, interleukin 6 (IL6), and tumor necrosis factor α (TNFα). Serum IL6 and TNFα levels were remarkedly suppressed in the 0.8% HML group. These results suggested that the favorable effect of HML on obesity-associated inflammation might be related in part to the decrease in adipose tissue and hepatic fat deposition and inflammation.
Introduction
Amain feature of obesity is fat accumulation in adipose tissue due to the chronic energy imbalance between excessive energy intake and energy expenditure. 1 The enlargement of adipose tissue induces the production and secretion of pro-inflammatory cytokine such as interleukin 6 (IL6), monocyte chemoattractant protein 1 (MCP1), and tumor necrosis factor α (TNFα). These pro-inflammatory cytokines lead to the macrophage M1 polarization that infiltrate the adipose tissue and the subsequent increase of local and systematic inflammation, responsible for the pathogenesis of obesity-associated metabolic diseases. 2 –5 Thus, inhibiting obesity-associated inflammation has been considered as a good tool for the prevention/treatment of its related comorbidities.
Mulberry (Morus alba L.) leaves have been medicinally used for diabetes, inflammation, and obesity. 6 –10 Mulberry leaves contain plenty of bioactive chemicals having potential antioxidant, antidiabetic, or anti-inflammatory properties. 11 –13 Mulberry leaf powder containing chlorogenic acid, gallic acid, and tricetin has been proposed to regulate insulin receptor substrate 1, IL6. peroxisome proliferator-activated receptor γ (PPARγ), and TNF. 14 Furthermore, chlorogenic acid derivatives found in mulberry leaf extract inhibits hepatic lipid infiltration, oxidative stress, and IL6 and TNFα release, offering promise as a nutraceutical agent for preventing obesity and its related nonalcoholic fatty liver disease (NAFLD). 9,15
High hydrostatic pressure (HHP) processing was purported as a favorable nonthermal processing method for high-quality mulberry juice. 16 Compared with the thermal processing, the antioxidant activity of mulberry juice and contents of resveratrol, total flavonoid, and total phenolic were significantly higher when using the HHP method. The use of low temperature HHP helps to preserve heat-sensitive biologically active compounds. 17 As described in our previous study, polyphenols such as rutin, isoquercitrin, nicotiflorin, morkotin, and astragalin in HHP-treated mulberry leaf extract (HML) were identified by ultra performance liquid chromatography-photodiode detector-quadrupole/time of flight-mass spectrometry. 18 The specific aim of this study was to investigate the beneficial effects of HML on obesity and its associated inflammation in obese rats.
Materials and Methods
Preparation of mulberry leaf extract
HML was provided by the Korea Food Research Institute (KFRI, Wanju, Korea). Mulberry leaves were harvested from the Sangju area (Korea) in May 2017, then ground and mixed with water and enzymes, such as Viscozyme, Celluclast, and Termamyl. The mixture was poured into plastic bags and treated in a TFS-2L high-pressure machine (Toyo Koatsu Co. Ltd., Hiroshima, Japan) under 90 MPa at 50°C for 24 h. After extraction, the extracts were boiled to inactivate the enzymes. After centrifuging the cooled extracts at 11,000 g for 10 min, the collected supernatant was filtered, and freeze-dried to form a powder.
Animals and diets
Three-week-old male Sprague-Dawley rats were purchased from Doo-Yeol Biotech (Seoul, Korea) and were individually housed in a stainless steel cage with a 12-h light and dark cycle. All animal experimental protocols were approved by the Institutional Animal Care and Use Committee (IACUC) of Ewha Womans University (IACUC No. 17-033). Rats were randomized and fed a normal diet (NOR), a 45% high-fat (HF) diet, HF premixed with 0.4% HML (ML), and HF supplemented with 0.8% HML (MH), respectively, for 14 weeks. The NOR diet consisted of 70% carbohydrate, 20% protein, and 10% fat, based on D12450H (Research Diets, New Brunswick, NJ, USA).
The HF diet (45% of energy) contained 21% lard, 2.9% soybean oil, 23% casein, 20% sucrose, 8.5% starch, 12% dextrose, 6.0% cellulose, 1.2% minerals, and 1.2% vitamins (w/w), based on a D12451 diet (Research Diets). Body weight and food intake were weekly measured. Feces were collected on the last 3 days of the experiment and stored at −40°C. At the end of 14 weeks, rats were fasted overnight and anesthetized with Zoletil (Virbac Laboratories, Carros, France)/Rompun (Bayer Korea, Seoul, Korea). Blood samples were collected by cardiac puncture and liver, epididymal white adipose tissue (eWAT), and mesenteric and perirenal WAT were dissected, weighed, and snap frozen until further analysis.
Serum biochemical profiles
Serum levels of alanine transaminase (ALT), aspartate transaminase (AST), high-density lipoprotein-cholesterol (HDL-C), total cholesterol (TC), and triglyceride (TG) were carried out by using commercial kits (Asan Pharmaceutical Co., Ltd., Seoul, Korea). In brief, 20 μL of serum was added to 100 μL prewarmed ALT or AST substrate buffer, mixed, and incubated at 37°C for 30 or 60 min for ALT or AST determination. The reaction was carried out by adding 100 μL colorimetric reaction buffer, incubated at room temperature (RT) for 20 min and terminated by 1 mL of 0.4 N NaOH solution. After 10 min incubation at RT, an UV/visible spectrophotometer (ultrospec 2100 pro; Amersham Biosciences, NJ, USA) was used to measure the absorbance at a wavelength of 505 nm. Serum ALT and AST levels were calculated by using their standard curve.
For serum TC and TG measurements, 10 μL of serum was mixed with 1.5 mL of TC or TG enzyme solution. After 5 min incubation at 37°C for 5 min for TC determination or 10 min for TG assay, the absorbance of the mixture was measured at 500 nm against a blank (Amersham Biosciences, NJ, USA) and the serum concentrations of TC and TG levels were determined by their respective standard curves.
For serum HDL-C analysis, the precipitation buffer was mixed with serum, incubated at RT for 10 min, and then centrifuged at 841 g for 10 min. The enzyme solution was added to the supernatant and incubated at 37°C for 5 min. The absorbance of the mixture was measured at a wavelength of 500 nm (Amersham Biosciences, NJ, USA).
Serum low-density lipoprotein-cholesterol (LDL-C) level was calculated by the Friedewald equation: (LDL-C = TC−HDL-C−[TG/5]). 19 Nonesterified fatty acid (FFA) concentrations in serum were measured using a commercially available kit (Wako Pure Chemical Corporation, Osaka, Japan).
Hepatic and fecal lipid analyses
Fecal samples (0.5 g) and liver samples (0.1 g) were used for lipid extraction using a modified method developed by Bligh and Dyer. 20 For homogenization, 1.5 mL of 0.9% saline and 7.5 mL of methanol:chloroform (2:1, v/v) were added to liver tissues or feces. After the addition of 2.5 mL chloroform, the mixture was shaken horizontally for 10 min. The lower phase was collected and dried after centrifugation at 2000 g for 10 min. After complete evaporation, total hepatic lipid was weighed, dissolved using a mixture of n-hexane/isopropanol (3/2, v/v), and stored in fresh tube at −40°C for analysis. TC and TG concentrations were determined as described earlier.
Histological analysis
Dissected liver tissue or eWAT was fixed overnight in 10% formalin. Fixed tissues were then embedded in paraffin, sliced into 5-μm-thick sections, and stained with hematoxylin and eosin (H&E). Digital images were acquired with a microscope (Olympus, Tokyo, Japan).
Immunohistochemistry analysis
Formalin-fixed paraffin-embedded eWAT sections were deparaffinized, rehydrated, and incubated with polyclonal anti-rabbit F4/80 antibody solution (GeneTex, Inc., Irvine, CA, USA). After the subsequent incubation with Polink-2 Plus HRP anti-rat DAB detection kit (Golden Bridge International, Inc., Irvine, CA, USA), the eWAT sections were then stained with diaminobenzidine (Golden Bridge), and counterstained with Mayer hematoxylin (ScyTek Laboratories, Logan, UT, USA). Crown-like structures (CLSs) were identified as single adipocytes surrounded by F4/80-positive macrophages and captured with an Olympus microscope.
RNA isolation, reverse transcription, and real-time quantitative polymerase chain reaction
Total RNA isolation from eWAT or liver tissue was carried out by RiboEx (GeneAll Biotechnology Co., Daejeon, Korea) according to the manufacturer's introduction. cDNA was generated from isolated RNA using MMLV reverse transcriptase (Bioneer Corp., Daejeon, Korea) and real-time quantitative polymerase chain reaction (qPCR) was performed using AccuPower 2X Greenstar qPCR Master Mix (-ROX dye) (Bioneer Corp.) and a Rotor-Gene 2000 (Corbett Research, Mortlake, Australia). Fold change was calculated as the 2−ΔΔCt method 21 compared with the housekeeping gene, β-actin. Supplementary Table S1 demonstrated used primers in this study.
Serum IL6 and TNFα measurement
Serum or IL6 or TNFα levels were determined using the LEGEND MAX(TM) rat IL-6 enzyme-linked immunosorbent assay (ELISA) kit or rat TNFα ELISA MAX™ deluxe kit (BioLegend, Inc., San Diego, CA, USA) using quantitative sandwich enzyme immunoassay technique with rat IL6 or TNFα antibodies. Biotinylated polyclonal biotinylated anti-rat IL6 antibody or anti-rat TNF-α detection antibody was added and followed by the addition of avidin-horseradish peroxidase and tetramethylbenzidine (TMB) substrate solution to produce antibody-antigen-antibody “sandwich” or a blue color in proportion to the concentration of IL6 or TNFα present in the samples. After the addition of stop solution, the absorbance was measured at a wavelength of 450 nm using a Varioskan plate reader (Thermo Scientific, Waltham, MA, USA).
Serum levels of nitric oxide
Serum nitrite oxide concentration was quantified to measure serum nitric oxide (NO) production using a commercially available kit (Thermo Scientific). To evaluate serum nitrite and nitrate in serum, a colored azo dye product of the Griess reaction using the enzyme nitrate reductase to convert nitrate to nitrite was detected at 540 nm using a microplate reader (Thermo Scientific).
Statistical analysis
Data are expressed as means ± standard errors (SEs). One-way analysis of variance, followed by Tukey's multiple comparison test, was employed to compare the statistical difference among the groups. Statistical significance was defined as P < .05 using SPSS software (version 23; IBM Corp., Armonk, NY, USA).
Results
Effect of HML on body weight
As shown in Table 1, the initial body weights were not statistically different. After 14 weeks on the experimental diets, rats fed the MH diet had a significantly reduced final body weight and body weight gain by 10.94%, respectively, compared with animals that consumed the HF diet, without changes in food intake, energy intake, and energy efficiency ratio.
Effect of High Hydrostatic Pressure Mulberry Leaf Extract on Body Weight and Food Intake
Values are expressed as mean ± SEM (n = 7). Different letters (a, b, c) show significant difference (P < .05).
Energy efficiency = body weight gain (g/day)/energy intake (kcal/day).
ALT, alanine transaminase; AST, aspartate transaminase; EER, energy efficiency ratio; HF, 45% high fat diet; MH, HF +0.8% high hydrostatic pressure mulberry leaf extract; ML, HF +0.4% high hydrostatic pressure mulberry leaf extract; NOR, normal diet.
Effect of HML on serum, hepatic, and fecal lipid profiles
Figure 1A illustrates that HF-fed rats had higher serum lipid levels than rats fed a NOR diet. Serum concentrations of TG, TC, FFA, and LDL-C in the MH group were significantly reduced to 58.0, 91.4, 46.0, and 56.1%, respectively, when the HF group was regarded as 100%.
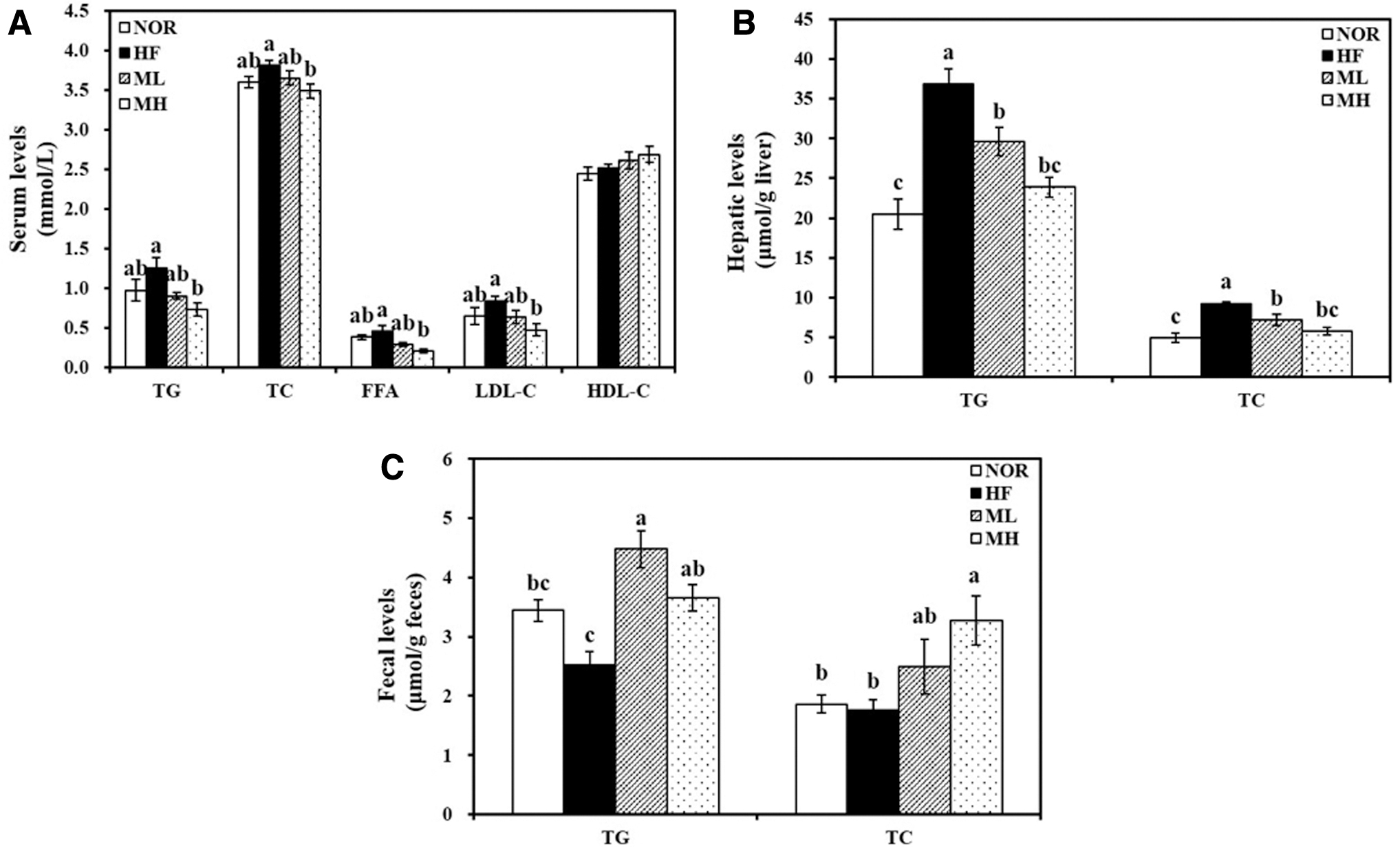
Effect of HHP extract of mulberry leaf on serum, hepatic, and fecal lipid profiles.
Animals fed with 0.4% or 0.8% HML showed a significant decrease in hepatic TG and TC (Fig. 1B). The ML and MH groups significantly inhibited HF-increased hepatic TG by 19.6% and 35.2%, respectively. Moreover, the hepatic TC level was significantly decreased with ML and MH treatment to 77.5% and 62.3%, respectively, when the HF group was 100%.
Fecal TG levels were increased by at least 1.45-fold in both HML-fed groups compared with the HF-fed group (Fig. 1C). Moreover, a dose-dependent increase in fecal TC concentrations was observed by the HF diet supplemented with HML (Fig. 1C).
Effect of HML on fat accumulation and mRNA levels involved in fatty acid oxidation and fat uptake and de novo synthesis in the liver tissue
Hepatic total lipid concentrations of the HML groups (ML and MH) were significantly reduced by 24% or 29% and was accompanied by mitigated HF-induced hepatic morphology (Fig. 2A, B). Fat levels in the diet or dose of HML did not change the serum ALT and AST activities (Table 1).
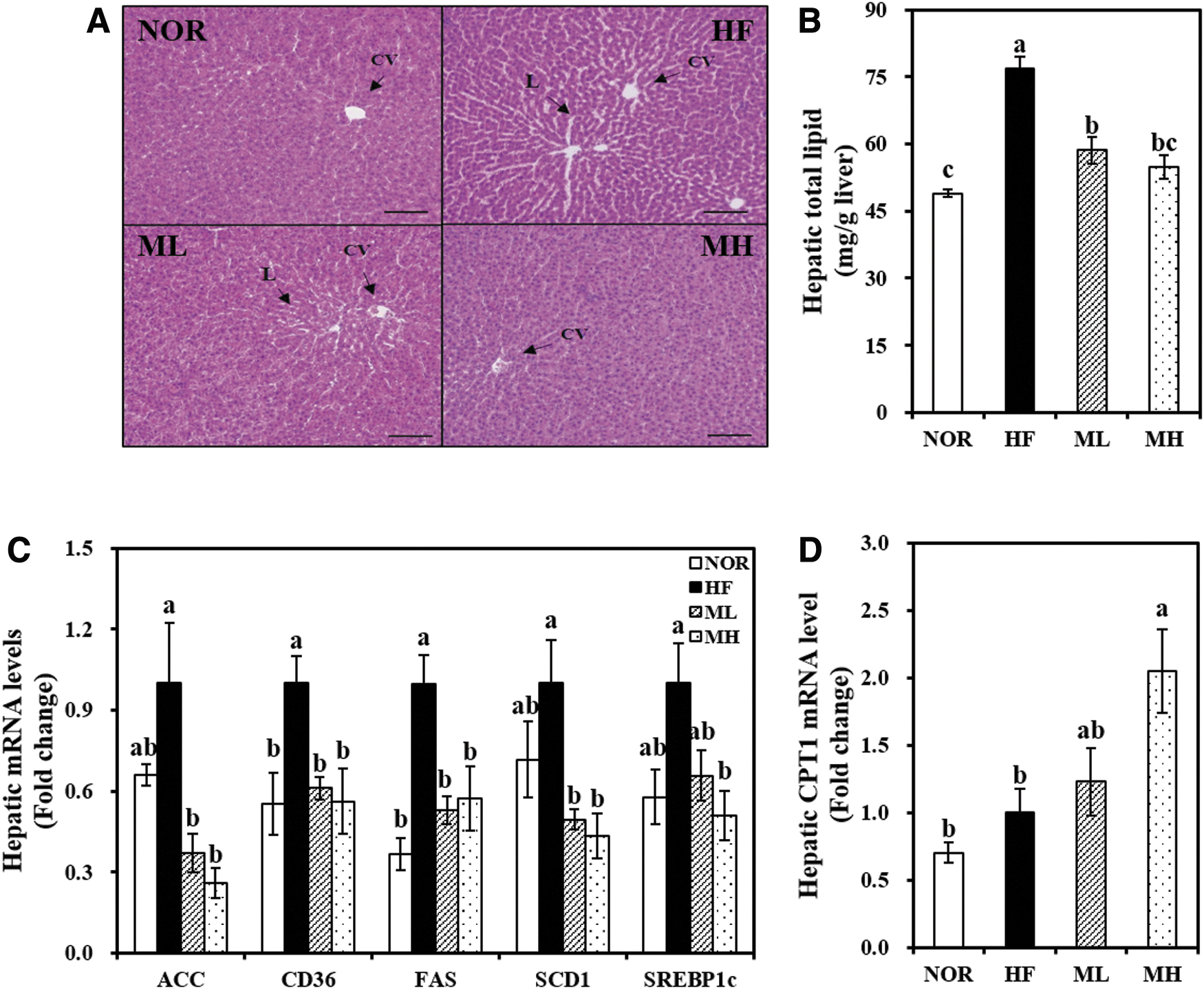
Effects of HHP mulberry leaf extract on hepatic expression related to fatty acid uptake and de novo lipogenesis.
The hepatic mRNA expression of cluster of differentiation 36 (CD36) was significantly inhibited by 44.0% in the MH group compared with the HF group (Fig. 3C). The hepatic acetyl-CoA carboxylase (ACC), fatty acid synthase (FAS), stearoyl-CoA desaturase (SCD1), and sterol regulatory element binding protein 1c (SREBP1c) gene expressions were significantly decreased by HML (Fig. 3C).
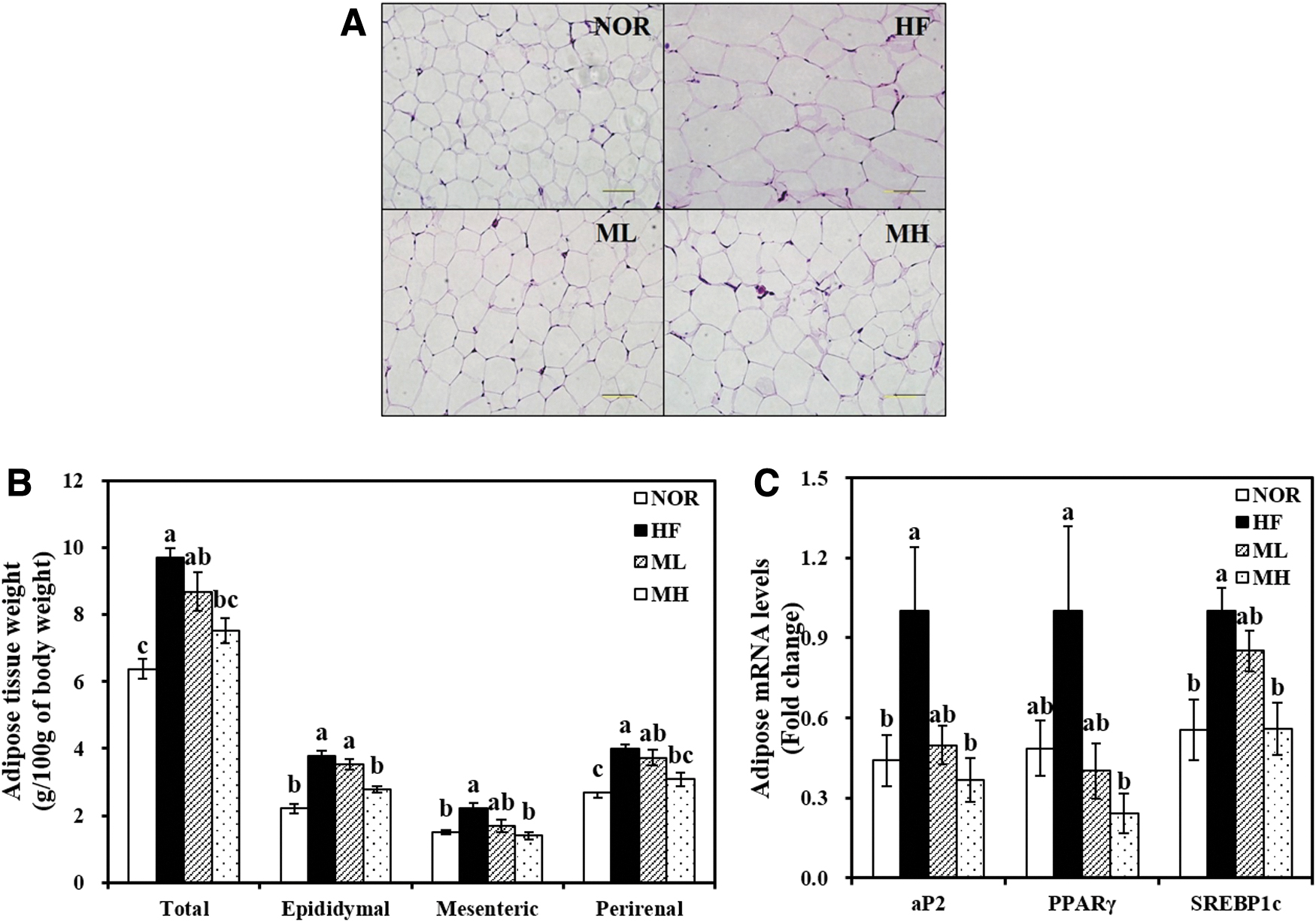
Effect of HHP extract of mulberry leaf on adipocyte hypertrophy and gene expression involved in adipogenesis.
HML significantly increased hepatic CPT1 gene expression in a dose-dependent manner by 1.23-fold by ML and 2.05-fold by MH (Fig. 3D). These data suggest that HML-decreased hepatic fat deposition might be associated with its decreased hepatic mRNA levels related to fat uptake, fatty acid oxidation, and synthesis.
Effect of HML on adipocyte hypertrophy and adipogenic gene expression
HML significantly suppressed the weight of WATs, including eWAT, mesenteric, and perirenal, and total WAT weight (P < .05) (Fig. 3B). A 25.8% significant reduction of eWAT weight was observed in MH group, compared with the HF group (P < .05). Consistent with WAT mass, H&E-stained adipose tissue exhibited a smaller adipocyte size in the HML groups (Fig. 3A).
As shown in Figure 3C, aP2 gene expression was 50% and 63% lower in the ML and MH groups, respectively. In a dose-dependent manner, HML significantly suppressed PPARγ and SREBP1c gene expressions (P < .05).
Effect of HML on adipose tissue inflammation
HF-induced CLSs in which macrophages surround a single adipocyte were observed, but CLSs did not appear in the HML-supplemented groups (Fig. 4A). In addition, mRNA levels of M1 macrophage markers CD11c and inducible nitric oxide synthase (iNOS) were significantly suppressed by 44% and 37%, respectively, in the eWAT from MH-fed rats, compared with eWAT from HF-fed obese rats. In contrast, the high level of HML in the HF diet had about two fold higher gene expression of the M2 macrophage maker, arginase1 (Arg1) compared with the HF diet (Fig. 4B). Moreover, 0.8% HML significantly reduced the serum NO levels by 46% compared with the HF diet (Fig. 4C).
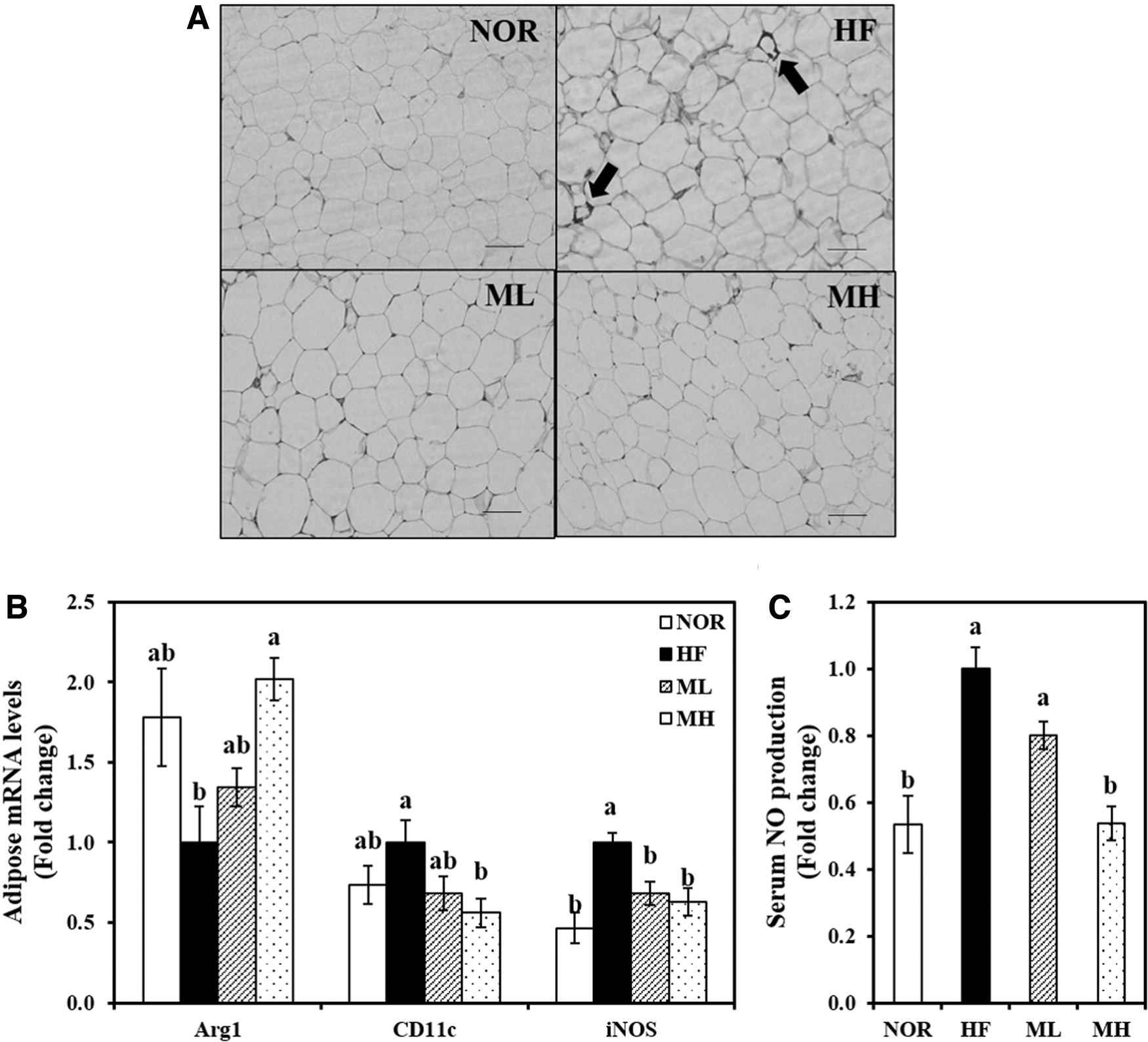
Influence of HHP extract of mulberry leaf on adipose tissue macrophage infiltration and polarization and serum NO concentration.
Effect of HML on adipocyte and hepatic mRNA expression involved in inflammation and systematic pro-inflammatory cytokine levels
The MH diet significantly suppressed the HF-increased hepatic mRNA expression of IL1β, IL6, and TNFα by 51%, 53%, and 52%, respectively, compared with the HF diet (Fig. 5A). In eWAT, HF induced at least a 1.5-fold increase in the gene expressions of IL6, MCP1, and TNFα (P < .05) (Fig. 5B). HML significantly downregulated the HF-induced IL6, MCP1, and TNFα gene expression in a dose-dependent manner, with reductions of 54%, 39%, and 54%, respectively (Fig. 5B).
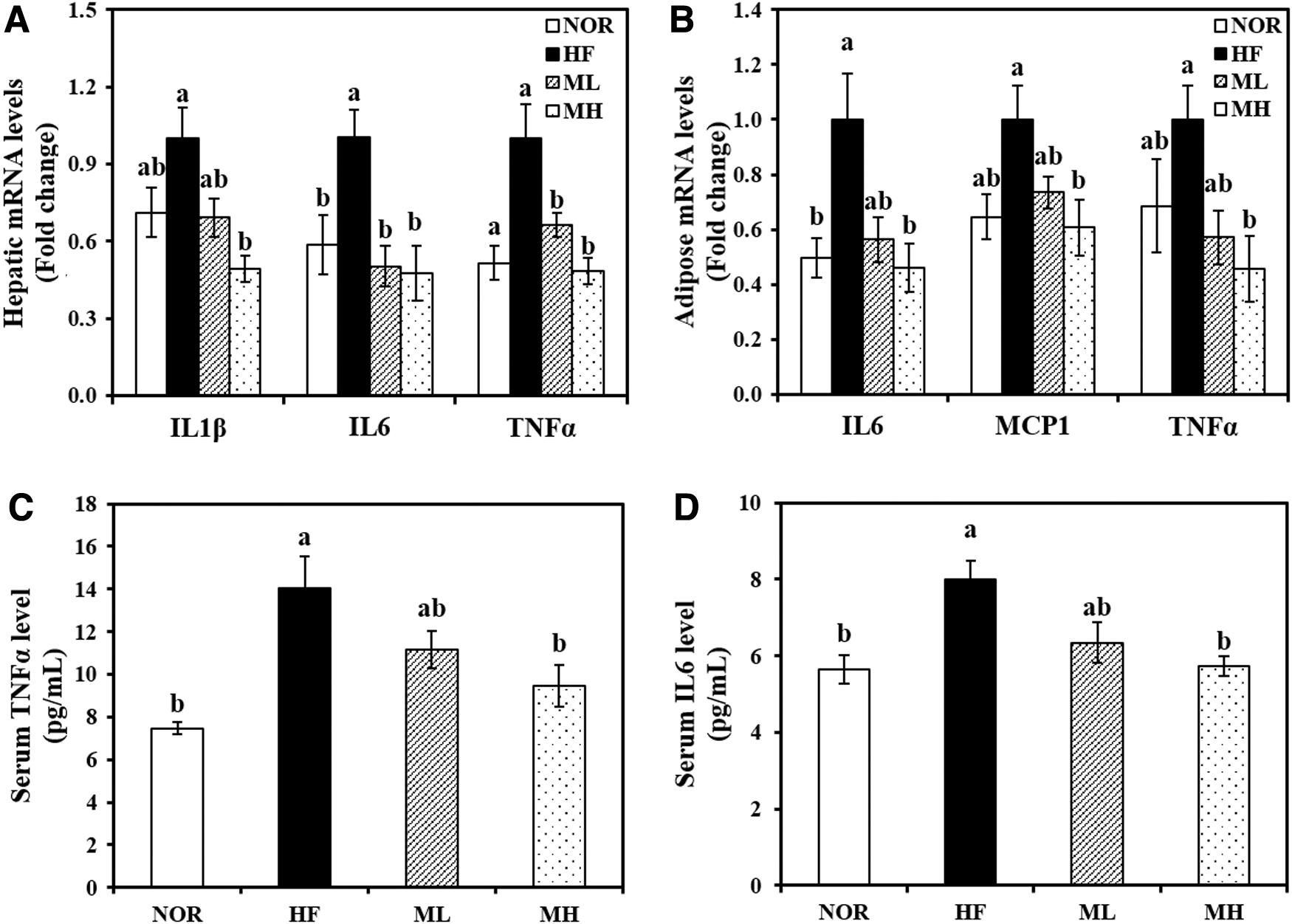
Effect of HHP extract of mulberry leaf on pro-inflammatory cytokines. Hepatic mRNA levels of IL1β, IL6, and TNFα
Similar to adipocyte and hepatic mRNA expression involved in inflammation, serum IL6 levels in the HF group were significantly increased by 1.41-fold, compared with the NOR group. A dose responsive serum IL6 reduction was observed with HML supplementation. Statistical significance was reached at the 0.8% dose (ML) (Fig. 5C). In addition, 0.8% HML significantly inhibited HF-increased serum TNFα concentration by 33% (Fig. 5D).
Discussions
Increased fat accumulation and inflammation in adipose tissue during the progression of obesity has been proposed as a pathophysiological link between obesity and its associated metabolic complications. 2,3,5 The use of herbal extracts including mulberry leaves might be a promising approach to attenuate obesity, atherosclerosis, diabetes, and inflammation. 6 –10
Similar to previous results from studying antioxidant, antidiabetic, or anti-inflammatory effects on mulberry leaves, 11 –14 in our previous study, HHP-processed mulberry leaves contained 2.99 ± 0.33 mg quercetin 3-O-rutinoside (rutin), 0.72 ± 0.07 mg quercetin 3-O-glucoside (isoquercitrin), 5.00 ± 0.03 mg quercetin 3-O-(6″-O-malonyl)glucoside, 1.17 ± 0.20 mg kaempferol 3-O-rutinoside (nicotiflorin), quercetin 3-O-(2″-O-malonyl)glucoside (morkotin), kaempferol 3-O-glucoside (astragalin), and 2.58 ± 0.46 mg kaempferol 3-O-(6″-O-malonyl)glucoside per 100 g of dried sample, respectively. There were 14.25 mg of total flavonols contained in 100 mg dried mulberry leaves extracted. 18 In this study, we investigated whether HML, obtained by nonthermal food processing technology (HHP) had beneficial effects on obesity and its associated inflammation in HF-fed rats.
The dosages of HML in this study were determined based on a previous study showing that mulberry leaves have positive effects on lipogenesis, fatty acid oxidation, and oxidative stress in rats. 22 Taking into account the average daily food intake and final body weight, we used concentrations of 0.4% and 0.8% HML in the HF diet, which correspond to ∼148 and 303 mg/day per kg body weight. These HML concentrations were well tolerated by rats, as indicated by no statistical difference in serum concentrations of ALT and AST.
Consistent with the close association between chronic excessive energy surplus and adipose tissue expansion, 1 –4 the HF diet induced significant increases in body weight, adipocyte size, and WAT weights, which were significantly alleviated by HML supplementation to the HF diet.
The fatty acid carrier protein, aP2, plays a critical role in adipocyte differentiation and fat accumulation. 23 PPARγ and SREBP1c are key transcription factors for adipogenesis or de novo fatty acid synthesis. 24,25 In this study, HML significantly downregulated the HF-increased gene expression of aP2, PPARγ, and SREBP1c. In previous studies, rutin and quercetin, which are present in high concentrations HML, inhibited aP2 and PPARγ adipogenic gene expression in HF-induced obese rats and adipocyte differentiation and lipogenesis in 3T3-L1 adipocyte cells, 26,27 suggesting that flavonoid-rich HML might attenuate obesity-induced adipocyte hypertrophy upon excessive energy intake.
Adipose tissue expansion leads to a local hypoxic state, which causes adipocyte death and macrophage localization and polarization. 4,5,28,29 In this study, ATM recruitment, characterized by CLSs, was not observed in the eWAT of HML-supplemented obese rats. Increased M1/M2 polarization and secretion of adipocyte-derived pro-inflammatory cytokines results in adipose tissue inflammation. 4,5 CD11c+ is a marker of pro-inflammatory immune cells related to macrophage infiltration in obese adipose tissue. 30 iNOS, an inflammatory marker, produces NO. 31
In contrast, Arg1 secreted from anti-inflammatory M2 macrophages inhibits iNOS activation. 32 In this study, HML supplementation significantly upregulated the HF-decreased Arg1 mRNA expression and decreased the HF-increased CD11c and iNOS mRNA levels in eWAT. The reduced serum NO production level in obese rats fed with HML was consistent with the M1 macrophage-related gene expression, suggesting that HML might improve obesity-associated ATM polarization to the M1 phenotype. In relation to obesity-associated pro-inflammatory adipokines, MCP1 is a chemotactic signal secreted from adipocytes and triggers macrophage recruitment. 33 IL6 and TNFα are pro-inflammatory cytokines produced and secreted from ATMs. 34
In this study, HML inhibited gene expressions of IL6, MCP1, and TNFα in adipose tissue of obese rats and reduced the serum IL6 and TNFα levels. Likewise, mulberry leaf extract decreases cytokines production of iNOS, IL6, and TNFα in RAW264.7 macrophage cells treated with lipopolysaccharide. 12 Therefore, HML was thought to attenuate obesity-induced inflammation through downregulation of gene expression related to inflammation in eWAT.
Given the close association between increased serum lipid profiles and obesity, excess circulating lipid concentrations trigger the inflammatory response and cause obesity-related NAFLD and dyslipidemia. 35 Consistent with previous studies that showed a favorable effect of mulberry leaves leaf extract on dyslipidemia, 8,36 in this study, HML significantly suppressed HF-induced serum FFA, LDL-C, TC, and TG concentrations, and decreased fat deposition was observed in H&E-stained liver tissues. Hepatic fat deposition results from the imbalance between hepatic lipogenesis and fat oxidation. CD36 is a membrane transporter for hepatic FFA uptake. 37 SREBP1c is a major transcription factor for lipogenesis by regulating ACC and FAS. 38
ACC catalyzes malonyl-CoA formation in fatty acid synthesis and inhibits CPT1 expression. 39 FAS catalyzes the production of free palmitate 40 and SCD1 is related to hepatic TG synthesis. 41 In addition, CPT1 transports fatty acids into the mitochondrial inner matrix for β-oxidation. 42 Previously, treatment with the hot water and ethanolic extracts of mulberry leaf suppressed the hepatic lipogenic protein levels of CD36, SREBP1c, FAS, and ACC. 7,36 Consistent with these results, in our study, 0.8% HML significantly decreased the hepatic gene expressions of CD36, ACC, FAS, SCD1, and SREBP1c and increased CPT1 expression. In light of this, HML might ameliorate hepatic fat accumulation by suppressing fatty acid uptake and lipogenesis and enhancing fat catabolism through regulating hepatic gene expression during the development of obesity.
In the pathogenesis of obesity-related metabolic diseases, increased fat deposition is responsible for dysregulated adipokine production. 43 TNFα, IL6, and IL1β are potent pro-inflammatory cytokines, and their levels are highly correlated with the degree of steatosis. 44 A previous study reported that mulberry leaves extracted with hot water reduced serum TNFα level. 9 In our study, HML remarkedly reduced the hepatic mRNA levels of IL1β, IL6, and TNFα, but also the systematic circulating IL6 and TNFα concentrations. These anti-inflammatory effects of HML might be partially associated with reducing hepatic fat deposition, local inflammatory cytokine levels in the liver tissue, and systemic inflammation levels.
This study demonstrates that HML attenuates obesity-associated adipose tissue mass and ATM recruitment associated with hepatic fat accumulation and inflammation. Therefore, this is the first study to suggest that mulberry leaves prepared by HHP might be useful as a nutraceutical to improve hyperlipidemia, hepatic steatosis and inflammation, and obesity.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This research was funded by the National Research Foundation of Korea (NRF) grant funded by the Korea government (MSIT) (2019R1A2C1002861) and Brain Korea 21 Four of the National Research Foundation funded by the Ministry of Education of Korea.
Supplementary Material
Supplementary Table S1