
Abstract
A number of studies employing different in vitro assays have demonstrated the estrogen-like activity of natural substances. All assays have their advantages and limitations as a screening tool. No single in vitro assay is considered ideal for predicting estrogenic action in a complex in vivo system. To assess agonistic activities of several medicinal herbs on the estrogen receptor (ER) and their metabolic alteration, the Organization for Economic Cooperation and Development (OECD) Performance-Based Test Guideline No. 455 in vitro assay was performed in this study using recombinant VM7Luc4E2 cells in combination with rat liver S9 fractions. Ethanol extracts of medicinal herbs showed binding affinities for ER-α and ER-β at different levels. However, luciferase reporter assay using VM7Luc4E2 cells revealed that only two test extracts [Pueraria lobata root extract (PLE); Glycyrrhiza glabra root extract (GGE)] exhibited ER transcriptional activity when their activities were compared with the response by 17β-estradiol. Importantly, incubation of PLE or GGE with rat liver S9 fractions increased their ER transcriptional activities, in particular when phase I metabolic enzymes were activated. Puerarin and glabridin were the most abundant isoflavones found in PLE and GGE, respectively. The present results demonstrate that PLE and GGE possess potential as ER agonists with their metabolic activation. This study also suggests that the application of OECD in vitro assay with rat liver S9 fraction is an efficient screening tool to evaluate estrogenic activities of natural substances.
Introduction
Estrogen deficiency in postmenopausal women affects overall life quality, including impaired reproductive functions, emotional changes, and an increased risk of metabolic syndromes. Although estrogen replacement represents the most effective treatment for relief of these symptoms, many researchers have expressed safety concerns, primarily associated with hormone-dependent cancer. 1 The use of selective estrogen receptor modulators (SERMs) known to display mixed estrogen receptor (ER) agonistic or antagonistic activity is one of the effective therapeutic strategies to offset the potential risk of estrogen treatment. 2 However, there are still concerns over adverse side effects of SERM observed in several clinical trials. 3 Against this backdrop, a number of medicinal herbs have been identified as potential estrogenic agents, suggesting that they might be utilized as natural alternatives to conventional estrogen therapy in the management of menopausal symptoms. 4,5
Phytoestrogens, mainly including isoflavones, coumestans, and lignans, are nonsteroid plant derived substances that exhibit either ER agonistic or antagonistic activity. It has been demonstrated that phytoestrogens can exert estrogenic or antiestrogenic activity not only through ER-dependent mechanisms with varied binding affinities for both ER-α and ER-β, but also through ER-independent pathways. 5,6 In humans, phytoestrogens undergo complex enzymatic metabolism in the gastrointestinal tract and liver, resulting in their conversion into inactive or active metabolites with different ER binding profiles. 7 Therefore, for evaluating (anti)estrogenic activities of natural substances containing phytoestrogens, a systematic approach considering phytoestrogen metabolism with cellular mechanisms of estrogen action is critical. However, in many studies using plant extracts, estrogenic properties have simply been screened with their proliferative effects on ER-positive cancer cell lines.
As a novel and effective screening model for estrogenic action of plant extracts, an in vitro ER transcriptional activation assay was performed in the present study using human breast cancer cell line VM7Luc4E2. This assay has been adopted as the Organization for Economic Cooperation and Development (OECD) Performance-Based Test guideline No. 455. 8 The VM7Luc4E2 cells, variant MCF-7 human breast cancer cells, endogenously express both human ER forms and have been stably transfected with an estrogen response element (ERE)-luciferase reporter gene. The test method measures transcriptional activity via ER-mediated pathways in recombinant VM7Luc4E2 cells, and their relevance and reliability for identifying human ER agonists and antagonists of various chemicals have been demonstrated. 9 –11 In the present study, estrogen-like activities of several medicinal herbs reported to be natural sources of phytoestrogens 5,12 were examined using OECD-proved transactivation assay. Furthermore, to confirm metabolic modification of estrogenic activity by medicinal herb extracts, we investigated the shifts in transactivation ability with rat liver S9 fraction known to be the source of cytochrome P450-dependent monooxygenase system. 13,14 This study will provide useful information for comparing and understanding cellular estrogenic actions of medicinal herbs containing phytoestrogens.
Materials and Methods
Preparation of medicinal herb extracts
Five dried medicinal herbs (Pueraria lobata root, Glycyrrhiza glabra root, Rehmannia glutinosa root, Angelica gigas root, and Astragalus membranaceus root) and a dried deer antler were extracted with 10 volumes of 50% EtOH at 85°C for 8 h. The extracts were filtered, concentrated under reduced pressure, and then spray-dried. Prepared samples were stored at 4°C until future use.
Human ER competitive binding assay
Human ER-α and ER-β binding activities were examined using a PolarScreen™ Competitor Assay Kit, Green (Invitrogen, Carlsbad, CA, USA) according to the manufacturer's instructions. Fluorescence polarization values were measured at excitation wavelength of 480 nm and emission wavelength of 535 nm with a microplate reader (Envision; PerkinElmer, Wellesley, MA, USA). Relative binding affinity of a tested sample was calculated: [IC50 of 17β-estradiol (E2)/IC50 of tested sample] × 100.
Cell culture
Recombinant VM7Luc4E2 cells were provided by Dr. Michael Denison (University of California, Davis, CA, USA) and maintained in RPMI 1640 (Cellgro, Manassas, VA, USA) supplemented with 8% fetal bovine serum (FBS; Cellgro), penicillin (100 U/mL), and streptomycin (100 μg/mL). For analysis, cells were switched to phenol red-free Dulbecco's modified Eagle's medium (Gibco, Grand Island, NY, USA) supplemented with 4.5% charcoal-stripped FBS,
ERE-luciferase reporter assay
Luciferase reporter assay was performed using VM7Luc4E2 cells to evaluate effects of extracts on ER transcriptional activation with a method as described previously. 11 In brief, after incubating cells in 96-well culture plates for 24 h, estrogen-free medium (EFM) was replaced with new EFM containing E2 and test samples at serially diluted concentrations. After incubation for 24 h, the medium was removed from each well and replaced with luciferase assay solution (50 μL/well) (Steady-Glo® Luciferase Assay System; Promega, Madison, WI, USA). After shaking for 10 min at ambient temperature, luciferase activity was measured using a MicroBeta2 2450 plate counter (PerkinElmer). To control the quality of assays, responses of the cell line were continuously monitored using control wells in each assay plate according to test guidelines. 8 ER transcriptional activities of extracts were expressed as luciferase activity relative to E2.
Metabolic activation with rat liver subcellular fractions
To examine alterations of estrogenic activities of test extracts after liver metabolism, cellular assay with rat liver S9 fraction was carried out. Briefly, cells were incubated with S9 fraction (Rat Pooled Liver S9; Corning, NY, USA) alone (S9 inactive), S9+cofactor I (26 mM NADP+, 66 mM glucose-6-phosphate, 66 mM MgCl2) (S9-phase I active), or S9+cofactor I+cofactor II (40 U/mL glucose-6-phosphate dehydrogenase in 5 mM sodium citrate) (S9-phase I + II active) for 24 h. Cells were treated with extract alone or with extract and each S9 condition. After incubation, cells were lysed and centrifuged to measure luciferase activity.
Isoflavones analysis
Isoflavone contents in extracts were determined according to a method as described previously 15 with some modifications. Briefly, 100 mg of the extract powder was dissolved in 10 mL of 100% methanol. The solution was sonicated for 30 min, filtered, and injected to a high-performance liquid chromatography system equipped with a photodiode array detector (SPD-M30A; Osaka Soda, Osaka, Japan) for isoflavone analysis. The analyte was separated using a Capcell Pak C18 MG S5 column (250 × 4.6 mm; Osaka Soda). The solvent system consisted of two eluents: eluent A (0.1% acetic acid in distilled water) and eluent B (100% acetonitrile). A gradient elution was employed at different ratios of A:B (v/v); 8.5:1.5 (0–10 min), 7:3 (10–25 min), 0:10 (25–36 min), and 8.5:1.5 (36–50 min).
Data analysis
Values of IC25, IC50, PC20, and PC50 were calculated to detect effective concentrations of test samples relative to E2 control in accordance with the OECD test guideline. 8 Data were obtained as average values from three wells in each experiment. Data are expressed as mean ± standard error for three independent experiments on different days. Statistical differences were determined by one-way ANOVA with a Tukey-Kramer post hoc method using SAS software (SAS Institute, Inc., Cary, NC, USA). Statistical significance was considered when the P value was <.05.
Results
Binding affinities of medicinal herb extracts for ER-α and ER-β
Binding affinities of ethanol extracts of medicinal herbs were determined with a competitive ligand binding assay. Competitive binding curves of E2 and six extracts to ER are shown in Figure 1. IC25 and IC50 values of E2 and extracts are summarized in Table 1. Among the tested samples, Rehmannia glutinosa root extract (RGE) exhibited the highest binding affinity with relative binding affinity values of 0.043 for ER-α and 0.048 for ER-β. On the other hand, A. membranaceus root extract had the lowest affinity, showing the highest IC25 value and nondetectable IC50 value for both forms of ER. Pueraria lobata root extract (PLE) showed a higher binding affinity for ER-β than ER-α while other extracts had similar potencies for both ER forms.

Competitive binding curves of E2 and test extracts to ER-α
Estrogen Receptor Binding Affinities of Medicinal Herb Extracts
IC25 = the concentration of a tested sample equivalent to 25% of binding affinity of E2 at 1 mg/mL.
IC50 = the concentration of a tested sample equivalent to 50% of binding affinity of E2 at 1 mg/mL.
RBA (relative binding affinity) = (IC50 of E2/IC50 of tested sample) × 100.
AGE, Angelica gigas root extract; AME, Astragalus membranaceus root extract; DAE, Deer antler extract; E2, 17β-estradiol; ER, estrogen receptor; GGE, Glycyrrhiza glabra root extract; ND, not detected; PLE, Pueraria lobata root extract; RGE, Rehmannia glutinosa root extract.
Proficiency test for in vitro ER transactivation assay
To confirm the responsiveness of the test system for ER agonist, a proficiency test was performed according to the OECD test guidelines. 8 Fold induction of E2 passed the acceptable criteria for the ER agonist assay (Table 2). As positive standards, bisphenol A and butylbenzyl phthalate showed suitable ranges of potency expected for ER agonists. Spironlactone, a negative standard, showed no significant induction of luciferase activity. These results were in accordance with expected responses for each substance that was presented in OECD test guidelines.
Proficiency Test with In Vitro Estrogen Receptor Transactivation Assay
The concentration of a tested substance at which the response is 20% of the response induced by E2 (17β-estradiol).
The concentration of a tested substance at which the response is 50% of the response induced by E2 (17β-estradiol).
ER transcriptional activity of medicinal herb extracts
In the classical cellular mechanism of estrogen action, nuclear E2-ER complexes can directly bind to ERE in target gene promoters and regulate transcriptional processes. 16 In this study, because all tested samples showed varied binding activities for both ER forms, we next examined their transcriptional activities by performing ERE-luciferase reporter assay using recombinant VM7Luc4E2 cells. Among the tested samples, only PLE and Glycyrrhiza glabra root extract (GGE) showed dose-dependent increases in luciferase activity with a typical sigmoid shape (Fig. 2). When their activities were expressed based on response induced by E2, PLE exhibited a stronger ER agonistic potency than GGE, showing a lower PC50 and a higher relative transcriptional activity value than GGE (Table 3). RGE had the highest ER binding affinity but no ER transcriptional activity (Tables 1 and 3).
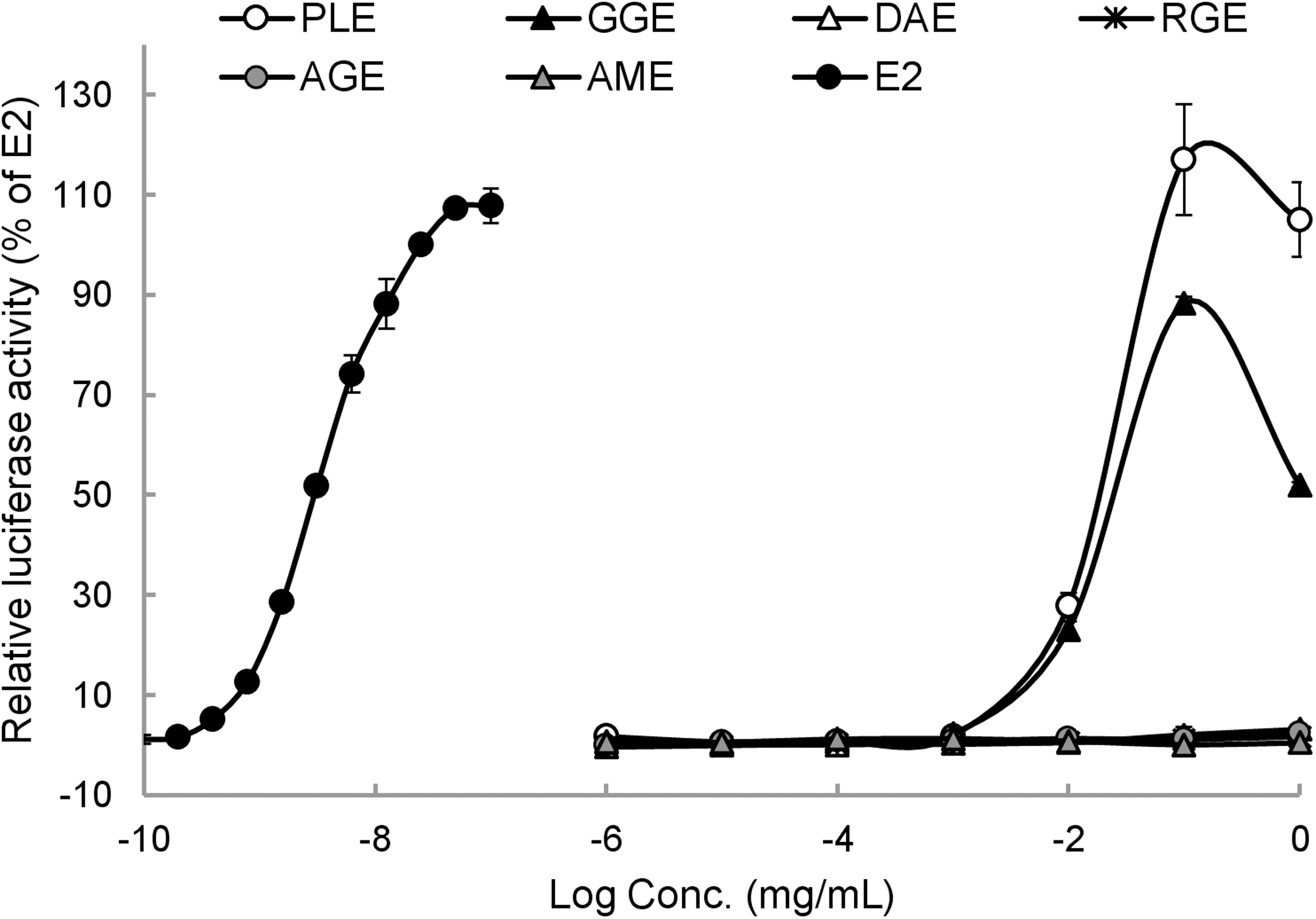
Luciferase activity of test extracts in ER transactivation assay using VM7Luc4E2 cells. The ER transcriptional activity of each extract was expressed as luciferase activity relative to E2. Data are shown as means ± SEM (n = 3).
Estrogen Receptor Agonistic Effects of Medicinal Herb Extracts in In Vitro Estrogen Receptor Transactivation Assay
The concentration of a tested sample at which the response is 20% of the response induced by E2.
The concentration of a tested sample at which the response is 50% of the response induced by E2.
RTA (relative transcriptional activity) = (PC50 of E2/PC50 of tested sample) × 100.
Besides ER agonistic effect, ER antagonistic effects of extracts were also evaluated using VM7Luc4E2 cells. When cells were treated with a combination of extract and E2, all extracts failed to inhibit E2-induced ER transactivation (Supplementary Fig. S1), indicating no ER antagonistic effect of tested plant extracts.
Metabolic modification of ER transcriptional activity
To address whether in vitro estrogenic activities observed for PLE and GGE would be maintained after hepatic metabolism, we further investigated shifts in ER transcriptional activity with rat liver S9 fraction. Figure 3 shows dose–response curves of PLE and GGE on luciferase activity under inactivation or activation of phase I and phase II liver metabolizing enzymes. Importantly, ER transcriptional activities of both PLE and GGE were significantly enhanced by liver metabolizing enzymes compared to S9 inactive control when their activities were compared based on PC20 and PC50 values (Table 4). In both PLE and GGE, the estrogenic activity was the highest with phase I enzymes and somewhat decreased with the activation of phase I + II (Fig. 3 and Table 4). These results suggest that ER agonistic activities of PLE and GGE could be enhanced with the formation of active metabolites through hepatic first-pass effects, in particular by P450 enzymes.
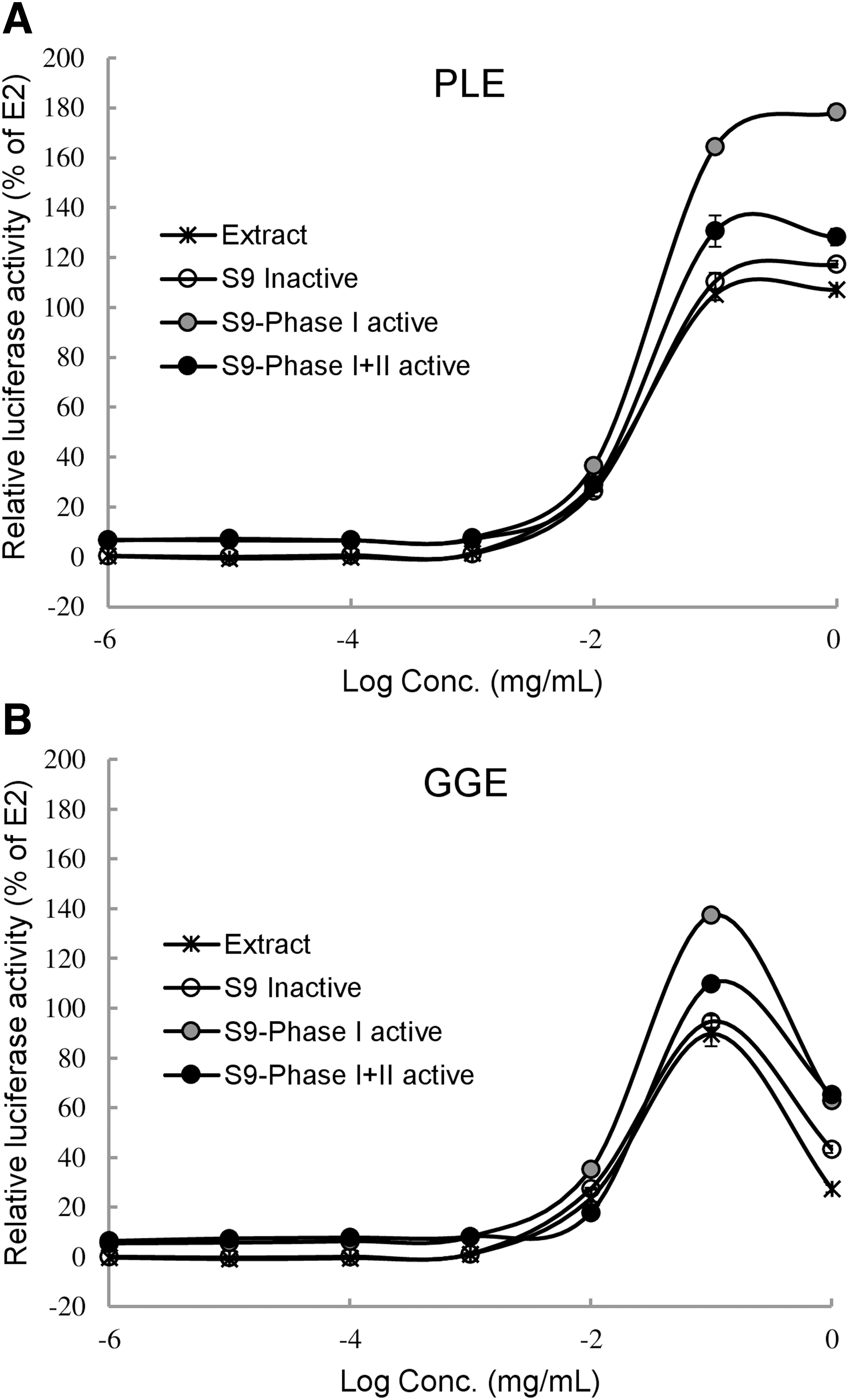
Metabolic activation of PLE
Estrogen Receptor Agonistic Effects of Selected Extracts with Rat Liver S9 Fraction
The concentration of a tested sample at which the response is 20% of the response induced by E2 (17β-estradiol).
The concentration of a tested sample at which the response is 50% of the response induced by E2 (17β-estradiol).
P < .05, ** P < .01 compared to S9 inactive control.
Isoflavone contents in PLE and GGE
Isoflavones are generally considered to be the major estrogen-like compounds from natural sources and may activate both genomic and nongenomic estrogen signaling pathways. 17 Based on previous reports on major isoflavones in P. lobata and G. glabra, 18,19 we identified seven isoflavones (puerarin, daidzin, genistin, daidzein, genistein, glycitein, and glabridin) from both PLE and GGE (Table 5). Total isoflavone content analyzed in this study was higher in PLE than in GGE. Puerarin was the most abundant isoflavone found in PLE, followed by daidzin. Among isoflavones, glabridin was the only isoflavone that had higher content in GGE than in PLE.
Isoflavone Contents in Extracts of Pueraria lobata Root and Glycyrrhiza glabra Root
Values represent means ± standard error (n = 3).
Discussion
The ER transactivation assay using VM7Luc4E2 cells has been widely used to identify endocrine disrupting chemicals since the test method was adopted as an OECD test guideline in 2012. In addition, the use of in vitro metabolizing system coupled with the ER transactivation assay has been suggested to predict the metabolic fate of xenobiotics by hepatic phase I and phase II reactions. 13 Indeed, metabolic activation or inactivation of various estrogen-like substances has been evaluated using different assay systems with rat liver S9 fractions. 14,20 The liver S9 fraction contains both microsomal and cytosolic components that can provide metabolic information including phase I and phase II reactions. It is relatively inexpensive and easy to use, making it an efficient in vitro screening system. 21 In this study, we investigated ER agonistic activities of several medicinal herb extracts by using the VM7Luc4E2 assay with S9 enzyme mixtures and found that estrogenic capacities of PLE and GGE could be enhanced after hepatic metabolism. All tested plant extracts showed relatively weak binding activities for both ER-α and ER-β compared to E2. However, the estrogen-mimetic effect on ER transactivation was only observed for PLE and GGE (Tables 1 and 3).
P. lobata and G. glabra in extract form has been shown to display estrogen-like activities in both in vitro and in vivo models. 22 –25 These studies have reported that their extracts could improve menopausal symptoms in ovariectomized mice and stimulate cell proliferation and estrogen-responsive gene expressions in breast cancer cells. However, their regulatory actions in cellular ER signaling are still unclear. Our current results, for the first time, demonstrated that PLE and GGE could promote ER-mediated transcriptional activation in VM7Luc4E2 cells. More importantly, ER transactivation by PLE or GGE was more prominent with S9 fraction treatment, suggesting their metabolic activation in the liver. It has been demonstrated that isoflavones are bioactive compounds responsible for estrogenic activities of numerous medicinal plants including P. lobata and G. glabra. 26
Consistent with previous studies reporting major isoflavone compounds in PLE and GGE, 18,19 our data showed that PLE and GGE had different isoflavone compositions. In the digestive system, dietary isoflavones undergo extensive intestinal and hepatic first-pass metabolisms that contribute to their bioavailability and physiological effects. 27 During enterohepatic circulation, isoflavones undergo phase I (cytochrome P450-mediated) and/or phase II biotransformation in the liver. Several studies have reported the significance of phase I reactions for metabolic activation of isoflavones, 28,29 whereas phase II reactions (conjugation pathway) have been shown to be associated with loss of biological activity of isoflavones. 30,31 These reports are consistent with our current findings showing the highest ER agonistic activities of both PLE and GGE with S9-phase I treatment (Fig. 3 and Table 4). Their ER agonistic activities with phase I + II fractions were lower than their activities with phase I, but still higher than the S9 inactive control. These results suggest that hepatic phase I and phase II biotransformation may generate more active metabolites than parent estrogenic compounds in PLE and GGE. However, further studies are needed to analyze in vivo isoflavone metabolites generated from PLE and GGE and investigate their individual estrogen-like actions.
Total isoflavone content was higher in PLE than in GGE, which accounts for the more potent ER agonistic effect of PLE observed in this study. Puerarin is a well-known major isoflavone in P. lobata, as confirmed by our data (Table 5). It has been demonstrated that puerarin exerts estrogen-mimic effects in ovariectomized rodent models. 32,33 Besides its estrogenic effects, several studies have shown that puerarin has inhibitory effects on estrogen-dependent endometriosis in both in vitro and in vivo models, 34,35 indicating its antiestrogenic effect in the uterus tissue. These findings suggest the potential action of puerarin as an SERM that can exert mixed estrogen agonist and antagonist activities depending on target tissues. In fact, the potential of many phytoestrogen compounds including isoflavones as natural SERMs has been demonstrated, particularly regarding their antagonistic activities against ER-α. 6,36 In this study using breast cancer cells, we did not observe ER antagonist activity for any tested plant extracts (Supplementary Fig. S1). However, their potential SERM functions in a tissue-specific manner remain to be explored.
Classical estrogen signaling is regulated depending on a balance between ER-α and ER-β activities in target organs. ER-α is highly expressed in the uterus, prostate, ovary, testis, epididymis, breast, and liver, while ER-β is expressed in prostate, testis, ovary, bone marrow, and brain. 37 VM7Luc4E2 cell is a genetic variant of the MCF-7 human breast cancer cell and predominantly express ER-α. Thus, our ER transactivation data indicate the effects of PLE and GGE are mainly mediated via ER-α. Although most phytoestrogen compounds have been shown to bind more strongly to ER-β than to ER-α, 38,39 their interactions with each ER subtype would provide different physiological functions.
In summary, our data demonstrated that PLE and GGE exhibited weak estrogen-mimetic effects on ER binding and transcriptional activation. In the presence of phase I or phase I + II hepatic enzymes, both PLE and GGE were metabolically activated to have higher ER transactivation abilities. These effects might be associated with the presence of phytoestrogen isoflavones in PLE and GGE. The present study suggests that ER transactivation assay using recombinant VM7Luc4E2 cells in combination with the rat liver S9 fraction could provide meaningful information for the evaluation of potential estrogenicity of phytoestrogen-rich medicinal plants.
Footnotes
Acknowledgments
The authors thank Dr. Michael Denison, UC Davis, for providing the VM7Luc4E2 cells.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the Kwang Dong pharmaceutical, South Korea and by the Chung-Ang University Graduate Research Scholarship in 2020.
Supplementary Material
Supplementary Figure S1