
Abstract
We investigated the effects of bonito fish (Katsuwonus pelamis) elastin HC (KE) on skin dryness, wrinkles, and pigmentation in vitro and in vivo. In vitro, we evaluated the expression of mRNA genes and proteins related to skin dryness, wrinkles, and pigmentation. HaCaT and HS27 cells were exposed to ultraviolet B radiation (UVB) (50 mJ/cm2), and B16F10 cells were stimulated with 3-isobutyl-1-methylxanthine (IBMX, 250 μg/mL) for 72 h to induce melanin synthesis. All cells were treated with KE (50–400 μg/mL) for 24 h. We found that KE increased the expression of long-chain base 1, dihydroceramide desaturase 1, elastin, hyaluronan synthase 2, and ceramide synthase 4 mRNA or protein as well as hyaluronic acid and sphingomyelin levels in UVB-irradiated HaCaT cells. Moreover, KE regulated factors related to collagen production, wrinkles, and melanin production in UVB-irradiated HS27 cells and IBMX-stimulated B16F10 cells.
In vivo
, we evaluated skin hydration and the expression of mRNA genes and proteins in the skin, and conducted morphological observations in SKH-I hairless mice (5-week-old male). The mice were exposed stepwise to UVB and given KE (10, 20, and 30 mg/kg b.w.) for 8 weeks. We found that skin hydration and protein or mRNA expression related to skin moisturization were increased in the KE group. Moreover, KE intake increased factors related to collagen production, wrinkles, and melanin production in UVB-irradiated SKH-I hairless mice. These results suggest that KE may have efficacy for the development of treatments for improving skin health.
INTRODUCTION
Skin photoaging is a representative phenomenon of extrinsic skin aging, and because of this, skin health is emerging as one of the main concerns of modern people pursuing external beauty. 1 To prevent dryness, wrinkles, and pigmentation of the skin, research is being actively conducted not only on cosmetics that are applied directly to the skin but also on food that can be consumed continuously. 2 Photoaging is characterized by dryness due to water loss in the skin, the occurrence of wrinkles, and red and black pigmentation. 3
Among the tissues constituting the skin, the epidermis contains keratinocytes and melanocytes, and 90% of the dermis contains fibrous components such as collagen and elastin. 4 As the moisture of the skin decreases due to the various characteristics of photoaging, the skin barrier becomes increasingly damaged, it becomes dry, and the generation of wrinkles is accelerated. Therefore, a constant supply of moisture is very important, and the activities of hyaluronic acid and hyaluronic acid synthase are known to play a crucial role in moisturization. 5 –7 In addition, reactive oxygen species (ROS) generated by exposure to ultraviolet radiation (UV) from sunlight promotes the activity of matrix metalloproteinase (MMP), known as collagenase, and interferes with collagen synthesis in the dermis, which leads to wrinkle formation and the loss of elasticity. 8,9
Melanin content is the skin is increased as a defense mechanism against UV exposure. Tyrosinase plays an important role in the production of melanin, and melanin functions in protecting the skin by removing ROS. However, excessive melanin production has been reported to cause spots, freckles, melasma, and even melanoma. 10 Therefore, research on and the development of natural materials that help maintain healthy skin are needed to facilitate strategies for alleviating skin dryness, wrinkle formation, and pigmentation caused by UV rays.
Food-derived peptides have been proposed to enhance elastin synthesis, the extracellular matrix, and antioxidant activity in the skin. 11 –13 Food-derived elastin peptides, such as Ala-Hyp, Ala-Hyp-Gly, Pro-Hyp, and Pro-Hyp-Gly, have been shown to have potential positive value for the skin. 11 Recently, as the biological efficacy, availability, and reasonable price of bonito fish (Katsuwonus pelamis), as well as the existence of elastin peptides have become known, it has been used in studies on the mechanism of photoaging. 12 However, research on its various mechanisms related to skin health is required. K. pelamis is one of the most abundant aquatic foods. It is composed of 70.3% water, 25.9% protein, 1.8% lipid, 0.3% carbohydrate, and 1.7% raw fish meal. It is one of the representative fish species that can be enjoyed by all age groups.
HaCaT cells, immortalized human keratinocytes, have been widely used to study the physiological mechanisms involved in UV irradiation of the skin. 14,15 HS27 is a normal human skin fibroblast cell line. This cell line is damaged by strong ultraviolet A radiation (UVA) treatment and has been widely used in research on antioxidants and skin protection. 16 SKH-I hairless mice are useful animal models for evaluating both UV ray irradiation damage and the genetic characteristics of tumor formation. 17 The light sensitivity of SKH-1 hairless mice also makes it a suitable UV-induced photoaging model. Therefore, in this study, we evaluated the efficacy of bonito elastin HC (KE) on skin dryness, wrinkles, and pigmentation using several cell models and SKH-I hairless mice and evaluated the value of its utilization for the development of functional foods for skin health.
MATERIALS AND METHODS
Preparation of materials
KE was supplied by Hagoromo Foods Corporation (Shizuoka, Japan). L-Ascorbic acid and arbutin (Sigma-Aldrich, St. Louis, MO) were utilized as the positive controls.
High-performance liquid chromatography–MS analysis
KE (20 mg) was weighed, and 0.4 mL of deionized water was added, as the solution (50 mg/mL). The solution was diluted to 1/10 to prepare a final 5 mg/mL solution. High-performance liquid chromatography (HPLC) analysis was performed with the Ultimate 3000 system (Dionex). A Poroshell 120 EC-C18 column (2.1 × 100 mm, 2.7 μm) was used for separation. The column temperature was 30°C, the flow rate was 200 μL/min, and the injection volume was 5 μL. The mobile phase consisted of 0.01 M phosphate buffer (pH 6.7, A) and acetonitrile (B). The gradient conditions were as follows: 0–5 min (5% B), 5–28 min (5–30% B), 28–40 min (30–95% B), and 40–46 min (95–5% B).
MS analysis was performed with the MicroQ-TOF III mass spectrometer (Bruker Daltonics, Germany). The mode was ESI+, the flow rate was 200 μL/min, and scan range was 50–2000 m/z.
Cell culture and treatments
HaCaT (human keratinocytes) cells were provided by Professor Hwang of the College of Life Sciences, Kyung Hee University. Hs27 (human fibroblasts) and B16F10 (melanoma) cells were purchased from ATCC. Each cell line was grown based on our conventional culture manuals. 18 Each cell line seeded for each experiment was washed, and HaCaT and Hs27 cells were exposed to 50 mJ/cm2 of UV rays using five Sankyo Denki G5T5 lamps (Sankyo Denki Co., Yokohama, Japan). Differentiation of B16F10 cells was accomplished by treating them with IBMX (250 μg/mL) for 72 h to induce melanin synthesis. 18 Next, all cells stimulated with ultraviolet B radiation (UVB) or IBMX were treated with ascorbic acid (100 μg/mL) and KE (50, 100, 200, and 400 μg/mL).
Experimental animals and treatments
All animal experiments were approved by the Institutional Animal Care and Use Review Committee of Kyung Hee University (KHGASP-20-347). Male SKH-I hairless mice (5 weeks old, 20 ± 1 g) were purchased from Saeronbio, Inc., (Uiwang, Korea). The housing facility was maintained at 23°C ± 2°C, with 50% ± 4% relative humidity and a 12-h light/12-h dark cycle.
The mice (total of 56) were randomly divided into seven groups with eight animals per group as follows: normal control (NC), UVB-irradiated control (C), positive control (PC) 1 (L-ascorbic acid 200 mg/kg b.w.), PC 2 (arbutin 200 mg/kg b.w.), and KE 10, 20, and 30 mg/kg b.w. All experimental diets were provided via oral administration. We used UVB lamp equipment (Sankyo Denki Co) for step-by-step mouse UVB exposure. UVB irradiation conditions were the same as those in our conventional method to induce skin dryness. 19 The experiment was conducted for 8 weeks, and skin tissue samples of the mice were collected for analysis.
Real-time polymerase chain reaction
HaCaT and Hs27 cells were seeded at 5 × 105 cells/well in a 6-well plate separately and stabilized. Total RNA was extracted from UVB-/KE-treated cells and animal skin tissues using the RNeasy Extraction Kit (Qiagen, Gaithersburg, MD). Next, RNA quantification, cDNA synthesis, and real-time polymerase chain reaction (RT-PCR) were performed as with our previous conventional method. 20 The gene sequences were designed using the Primer express program (Applied Biosystem, Thermo Fisher Scientific, Waltham, MA) (Table 1). RT-PCR was performed using a Bio-Rad CFX (Bio-Rad Laboratories, Inc., Hercules, CA). All results were automatically calculated as delta–delta CT values using Bio-Rad CFX Manager 3.1 (Bio-Rad).
Primer Sequences Used in Real-Time Polymerase Chain Reaction Quantification of mRNA
DEGS1, dihydroceramide desaturase 1; GAPDH, glyceraldehyde-3-phosphate dehydrogenase; LCB1 (SPT), long-chain base 1 (serine palmitoyltransferase); TGFβR1, transforming growth factor beta-receptor 1.
Western blotting
HaCaT, Hs27, and B16F10 cells were seeded at 5 × 105 cells/well in separate6-well plates, stabilized, treated with KE sample/UVB, or differentiated. Cells and animal skin tissues were homogenized using the CelLytic™ MT Cell Lysis Reagent (Sigma-Aldrich) with Halt™ Protease & Phosphatase Inhibitor Cocktail (Thermo Fisher Scientific). Subsequently, protein quantification, sample loading, and protein transfer were performed as in our previous protocol. 21 Primary antibodies and horseradish peroxidase-conjugated secondary antibodies were purchased from Cell Signaling Technology and Abcam (Cambridge, MA). The visualization and analysis were also conducted with our conventional method. 21
Enzyme-linked immunosorbent assay
HaCaT cells seeded at 1 × 104 cells/well in 96-well plates were stabilized and treated with KE sample/UVB. Sphingomyelin levels in HaCaT cells were measured using the Sphingomyelin Assay Kit (Abcam, Cambridge, United Kingdom), and the hyaluronic acid in the supernatant was measured using the hyaluronic acid ELISA Kit (Biovision, Inc., CA). HS27 cells seeded at 1 × 104 cells/well in 96-well plates were stabilized and treated with KE sample/UVB. Interleukin (IL)-1β, IL-6, and tumor necrosis factor (TNF)-α levels in the HS27 cell supernatant were measured using the Duoset ELISA Kit (R&D system, Minneapolis, MN).
B16F10 cells were seeded at 2 × 105 cells/well in a6-well plate, stabilized, differentiated, and treated with the KE sample. The activity of tyrosinase (Assay Kit, Abcam), nitric oxide (NO Assay Kit, Abcam), glutathione (Assay Kit, Biovision, Inc., Mountain View), cyclic adenosine monophosphate (cAMP ELISA Kit, Enzo Life Sciences, PA), superoxide dismutase (SOD Assay Kit, Biovision), catalase (Assay Kit, Biovision), and glutathione peroxidase (GPx Assay Kit, Biovision) was measured in B16F10 cells or the cell supernatant. All experiments were performed based on the Enzyme-Linked Immunosorbent Assay (ELISA) protocol according to the manufacturer's method.
Inhibition of melanin production
B16F10 cells seeded at 5 × 104 cells/well in a 24-well plate were stabilized, differentiated, and treated with the KE sample. After 72 h, the cells are washed with DPBS, 125 μL of cell lysis reagent buffer (Sigma) was dispensed to separate the cells, and the supernatant obtained was quantified for protein with Bradford Dye Reagent (Bio-Rad). Next, 300 μL of a solution containing 10% dimethyl sulfoxide (DMSO) in 1 N NaOH was added to the cells and dissolved at 100°C for 10 min, and absorbance was measured at 450 nm with an ELISA reader (Bio-Rad).
Measurement of skin hydration
Skin hydration in mice was measured before euthanization using a Howskin device (Innoinsight, Inc., Yongin, Korea). The moisture value was measured in such a way that the electronic terminus of the instrument was read from the back of the mice and transmitted to the application program of a mobile phone.
Hematoxylin & Eosin staining
Hematoxylin & Eosin (H&E) staining was conducted to measure skin wrinkles and thickness. After collecting the skin tissue samples from the mice and fixing them in 10% (v/v) formalin, paraffin blocks were created, sliced to a 5 μm thickness, and attached to slides. Additionally, after deparaffinization with xylene, hydrolysis was conducted in 100% alcohol, and the skin samples were stained with H&E and observed using a microscope (Nikon eclipse TS100, Tokyo, Japan).
Statistical analysis
All experimental results are presented as the mean ± standard deviations. We analyzed the significance of values with Duncan's multiple-range test after performing a one-way analysis of variance (ANOVA) using the SPSS statistical program (SPSS PASW Statistic 23.0, SPSS, Inc., Chicago, IL). Statistical significance was considered at P < .05.
RESULTS
HPLC-MS analysis of KE
HPLC analysis of the KE revealed two peaks matching those of the commercial standards desmosine and isodesmosine with retention times of ∼67.1 min and 69.3 min, respectively (Fig. 1). The KE contained 1–1.13 mg/g isodesmosine and 0.74–0.92 mg/g desmosine.
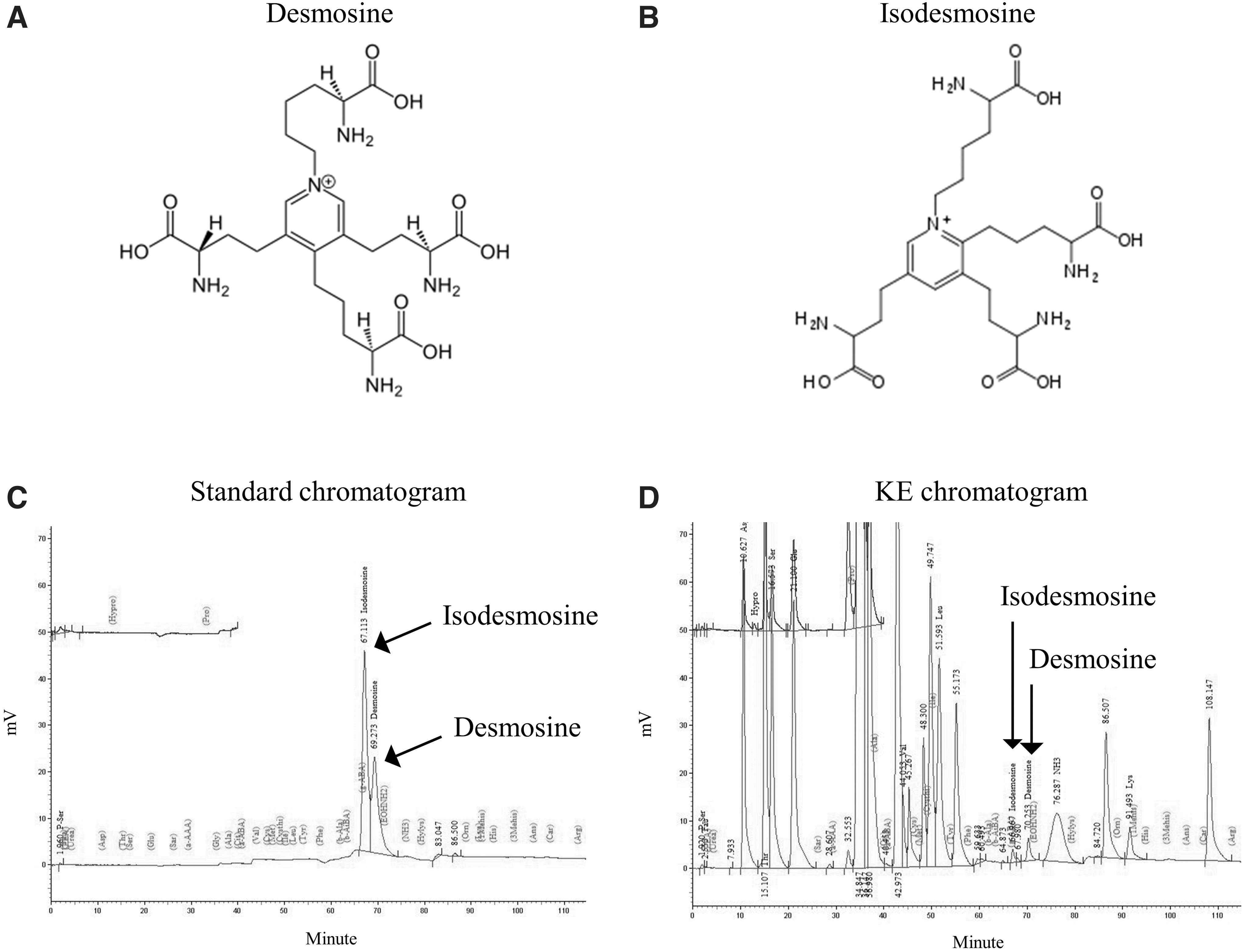
HPLC chromatograms of demosine and isodemosine in the KE.
Effects of KE on factors related to skin moisturizing in UVB-irradiated HaCaT cells
Testing mRNA expression related to skin moisturization revealed that long-chain base 1 (LCB1) expression was significantly increased in the KE groups (45.9% min to 73.4% max) compared with that in the C group (P < .05; Fig. 2A). KE increased the dihydroceramide desaturase 1 (DEGS1) expression (59.9% min to 115.1% max) and the elastin expression (32.6% min to 99.7% max) in a dose-dependent manner, compared with that in the C group (P < .05; Fig. 2B, C). Based on the protein expression, KE increased hyaluronan synthase 2 (HAS2) expression (45.9% min to 99.9% max) and ceramide synthase 4 (CerS4) expression (19.8% min to 42.6% max) compared with that in the C group (P < .05; Fig. 2D–F).
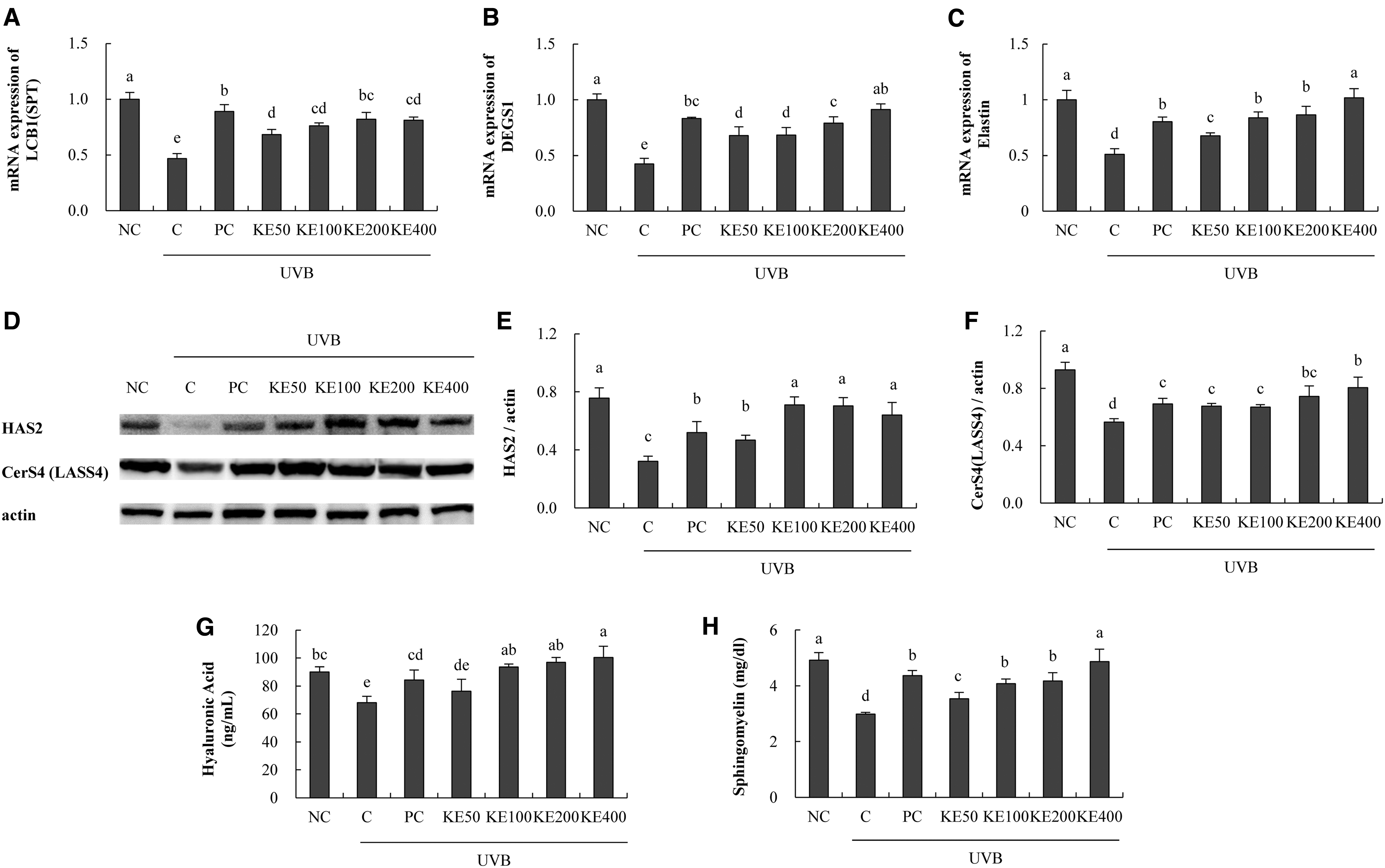
Effects of KE on factors related to skin moisturizing in UVB-irradiated HaCaT cells. RT-PCR, western blotting, and ELISA were performed.
Moreover, KE dose dependently increased the hyaluronic acid (11.9% min to 47.5% max) and sphingomyelin (18.8% min to 63.2% max) levels compared with those in the C group (P < .05; Fig. 2G, H). Increased expression of LCB1, DEGS1, and CerS4 can affect the synthesis of ceramide, and increased HAS2 expression affects the production of hyaluronic acid. Therefore, it is considered that KE treatment contributed to the increase of skin moisture in the HaCaT cell line.
Effects of KE on factors related to wrinkle suppression in UVB-irradiated HS27 cells
Evaluation of mRNA expression related to wrinkle suppression revealed that KE increased TGFβR1 expression (79.9% min to 172.1% max), procollagen type I expression (40.1% min to 100.7% max), and collagen type I expression (23.9% min to 71.2% max) compared with those in the C group (P < .05 Fig. 3A–C). For protein expression, KE decreased p-JNK/JNK expression (33.1% min to 56.1% max), p-c-Fos/Fos expression (29.0% min to 55.9% max), p-c-Jun/Jun expression (14.2% min to 46.7% max), MMP-1 expression (25.0% min to 32.3% max), MMP-3 expression (14.8% min to 23.9% max), MMP-9 expression (29.3% min to 50.1% max), and p-Smad3/Smad expression (26.8% max) compared with that in the C group (P < .05; Fig. 3D).
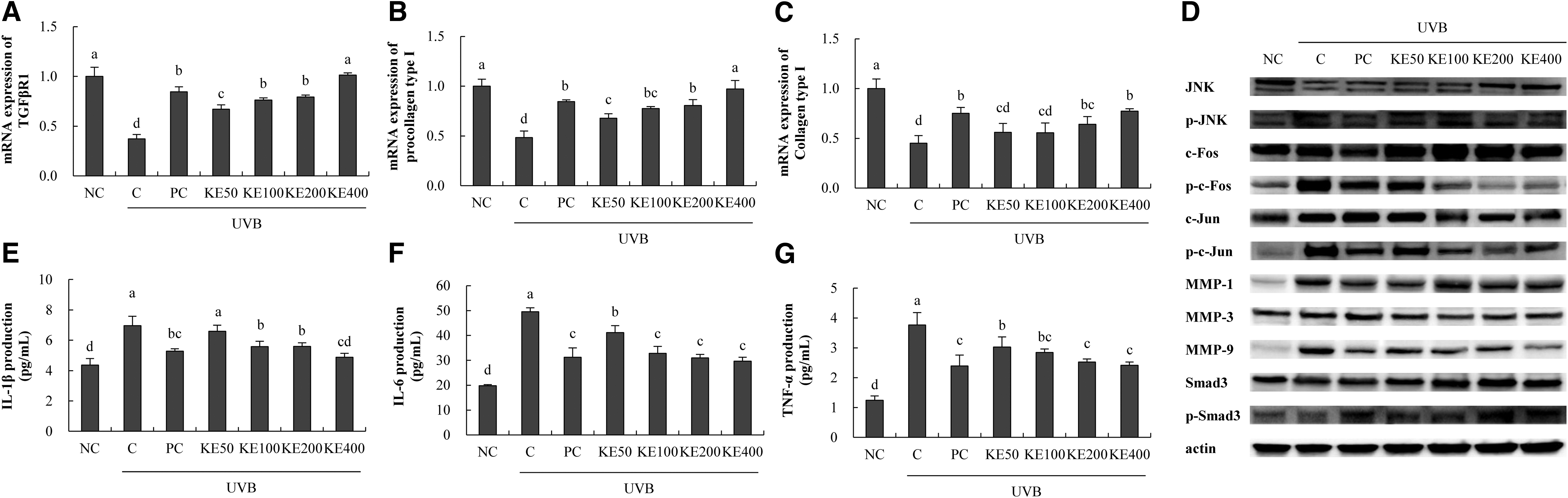
Effects of KE on factors related to wrinkle suppression and proinflammatory cytokines in UVB-irradiated HS27 cells. RT-PCR, western blotting, and ELISA were performed.
These factors are involved in collagen synthesis, and in particular, KE treatment modulated the change in the expression of MMPs, thus suggesting the possibility of contributing to wrinkle suppression in HS27 cell line.
Effects of KE on proinflammatory cytokines in UVB-irradiated HS27 cells
KE decreased IL-1β production (5.4% min to 29.8% max), IL-6 production (17.0% min to 40.2% max), and TNF-α production (19.5% min to 35.9% max) compared with levels in the C group (P < .05; Fig. 3E–G).
Effects of KE on factors related to whitening in melanogenesis-induced B16F10 cells
In UVB-irradiated B16F10 cells, KE decreased the melanin contents (28.6% min to 43.2% max) compared with those in the C group (P < .05; Fig. 4A, B). KE decreased tyrosinase activity (15.3% min to 34.8% max), NO (28.9% min to 40.8% max), and cAMP levels (25.1% min to 43.2% max) compared with those in the C group (P < .05; Fig. 4C, D, F). Moreover, KE increased the glutathione level (21.7% min to 97.6% max) compared with that in the C group (P < .05; Fig. 4E).
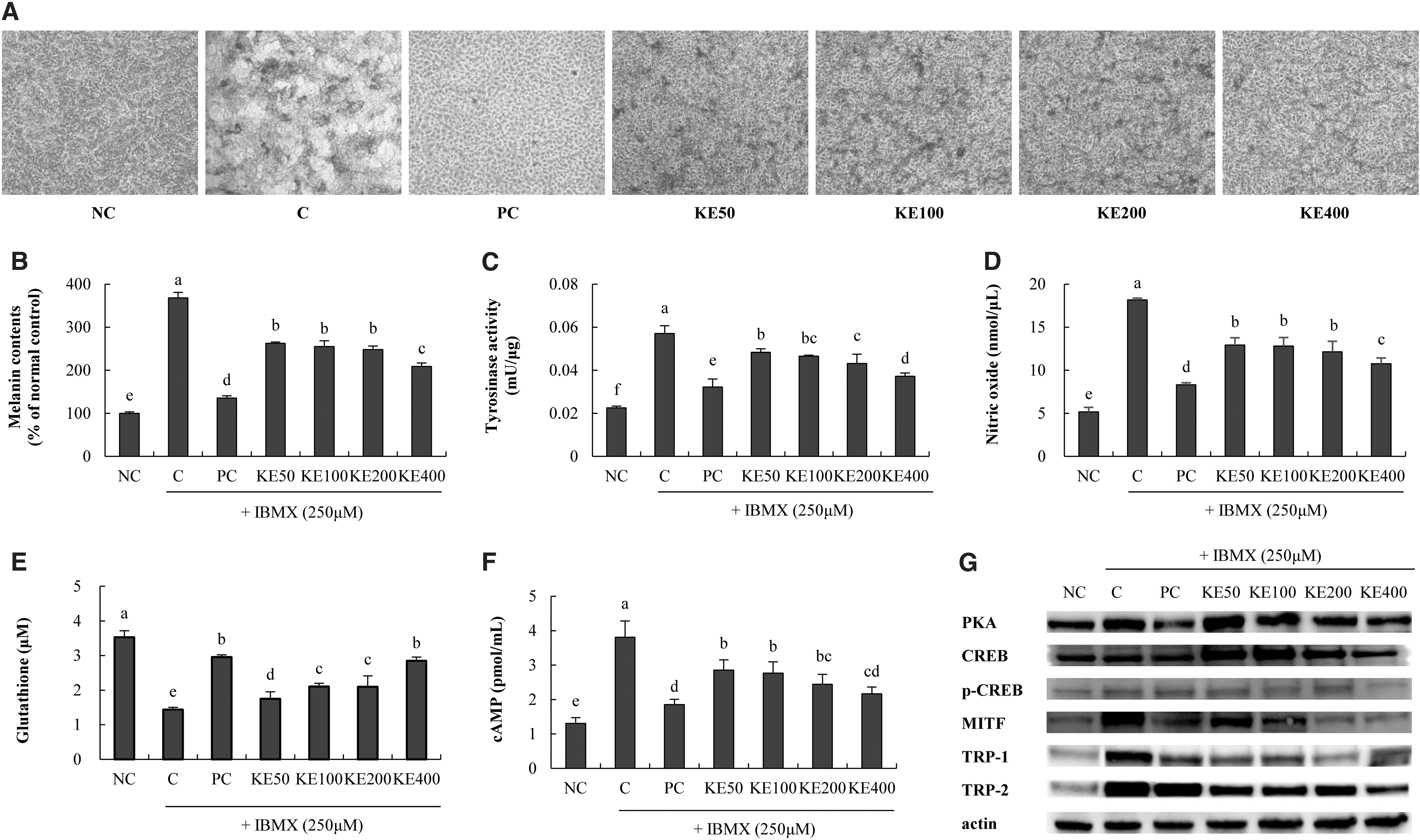
Effects of KE on factors related to whitening in IBMX-stimulated B16F10 cells. ELISA and western blotting were performed.
For protein expression, KE decreased PKA expression (7.2% min to 35.9% max), p-CREB/CREB expression (19.9% min to 56.0% max), MITF expression (16.3% min to 76.0% max), TRP-1 expression (45.7% min to 46.4% max), and TRP-2 expression (24.6% min to 49.4% max) compared with that in the C group (P < .05; Fig. 4G). In the melanogenesis mechanism, the cAMP-PKA pathway also affects the expression of lower levels of CREB and MITF, and finally changes the level of tyrosinase. Treatment with KE resulted in a decrease in melanin pigment, suggesting the possibility that it may exert whitening activity in the B16F10 cell line.
Effects of KE on body weight, food consumption, food efficiency rate, and organ weights of UVB-irradiated SKH-I hairless mice
Table 2 shows the effects of KE on weight gain, food intake, food efficiency rate (FER), and organ weights. There were no significant differences among all groups in the aforementioned parameters.
Body Weight Gain, Food Consumption, Food Efficiency Rate, and Organ Weight
Normal AIN93G, Control UVB irradiation+AIN93G, PC1: UVB irradiation+AIN93G+L-ascorbic acid 200 mg/kg∙b.w., PC2: UVB irradiation+AIN93G+Arbutin 200 mg/kg∙b.w., KE 10: UVB irradiation+AIN93G+Katsuwonus pelamis elastin 10 mg/kg∙b.w., KE 20: UVB irradiation+AIN93G+K. pelamis elastin 20 mg/kg∙b.w., KE 30: UVB irradiation+AIN93G+K. pelamis elastin 30 mg/kg∙b.w. The results are expressed in mean ± SD values. Significances are indicated using Duncan's test at P < .05.
Weight gain (g) = final body weight (g) – initial body weight (g).
FER (food efficiency rate) = {weight (g)/food intake (g)}*100.
NS, no significance; SD, standard deviation; UVB, ultraviolet B radiation.
Effects of KE on factors related to skin moisturization in UVB-irradiated SKH-I hairless mice
Evaluation of the mRNA expression related to skin moisturization revealed that KE intake increased the expression of LCB1 (8.2% min to 70.7% max), DEGS1 (9.4% min to 66.8% max), and fibrilin-1 (27.9% min to 78.4% max) compared with that in the C group (P < .05; Fig. 5A–C). Moreover, KE intake increased back skin hydration (25.4% min to 30.0% max) in the mice compared with that in the C group (P < .05; Fig. 5D). KE increased the protein expressions of HAS2 (3.2% min to 42.4% max) and CerS4 (15.2% min to 24.1% max) compared with that in the C group (P < .05; Fig. 5E–G). LCB1 and DEGS1 are enzymes involved in the production of ceramide, and the results were similar to those of in vitro experiments, and the moisturizing activity of KE by changing fibrilin-1 and HAS2 was confirmed.
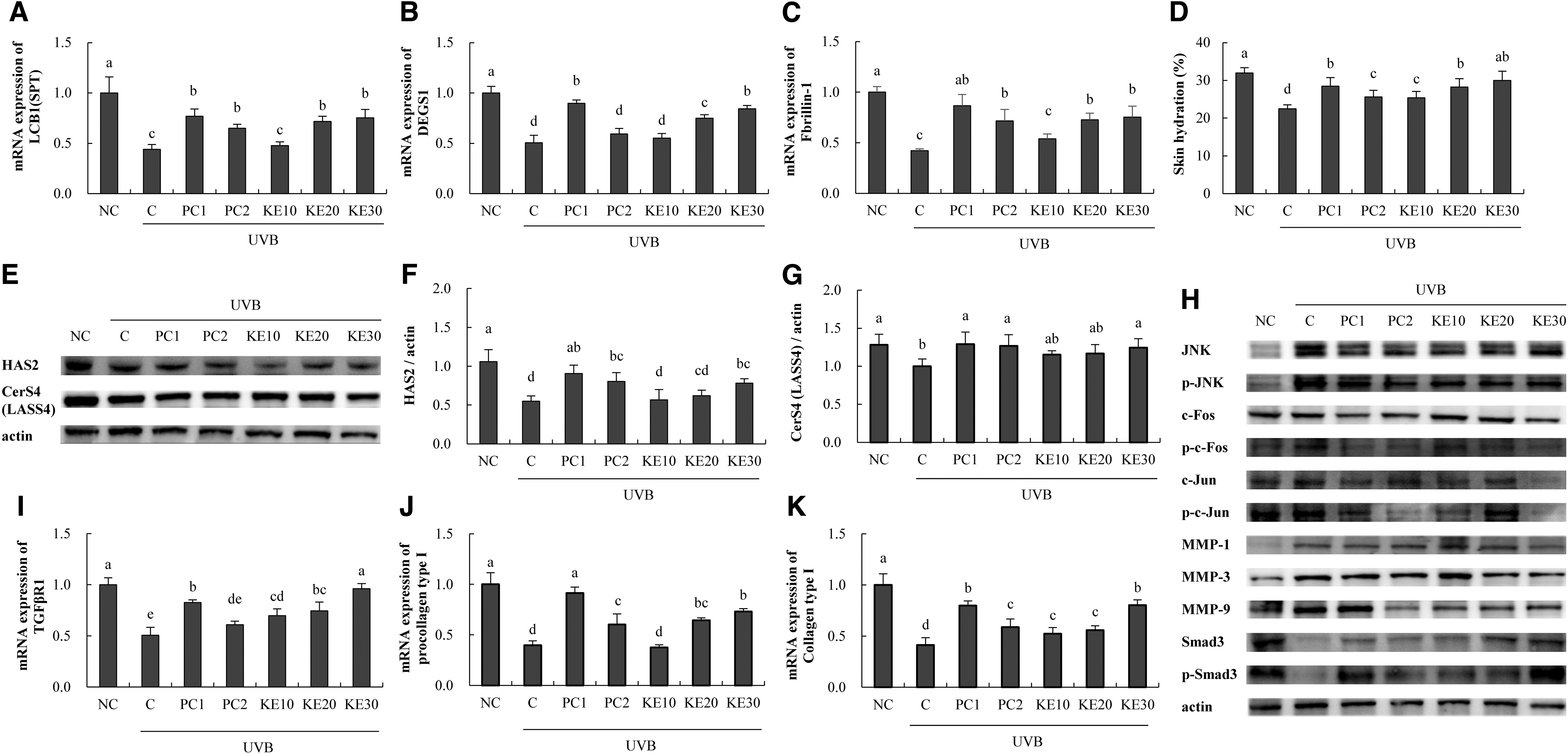
Effects of KE on factors related to skin moisturizing and wrinkle suppression in UVB-irradiated SKH-I hairless mice. RT-PCR, western blotting, and ELISA were performed.
Effects of KE on wrinkle formation, skin thickness, and antioxidant activity in UVB-irradiated SKH-I hairless mice
In UVB-irradiated SKH-I hairless mice, the groups receiving the PC1, PC2, and KE diets showed reduced wrinkle formation and skin thicknesses in a dose-dependent manner compared with those in the C group (Fig. 6A). Measurement of the antioxidant activity in UVB-irradiated SKH-I hairless mice revealed that SOD, catalase, and GPx activities in the C group were decreased (38.4%, 61.5%, and 47.3%, respectively) compared with that in the NC group. Furthermore, PC1 and PC2 intake increased SOD, catalase, and GPx activities compared with those in the C group. Moreover, KE intake increased SOD activity (6.6% min to 21.6% max), catalase activity (36.3% min to 72.9% max), and GPx activity (14.7% min to 37.3% max) compared with those in the C group (P < .05; 6C, D, and E).
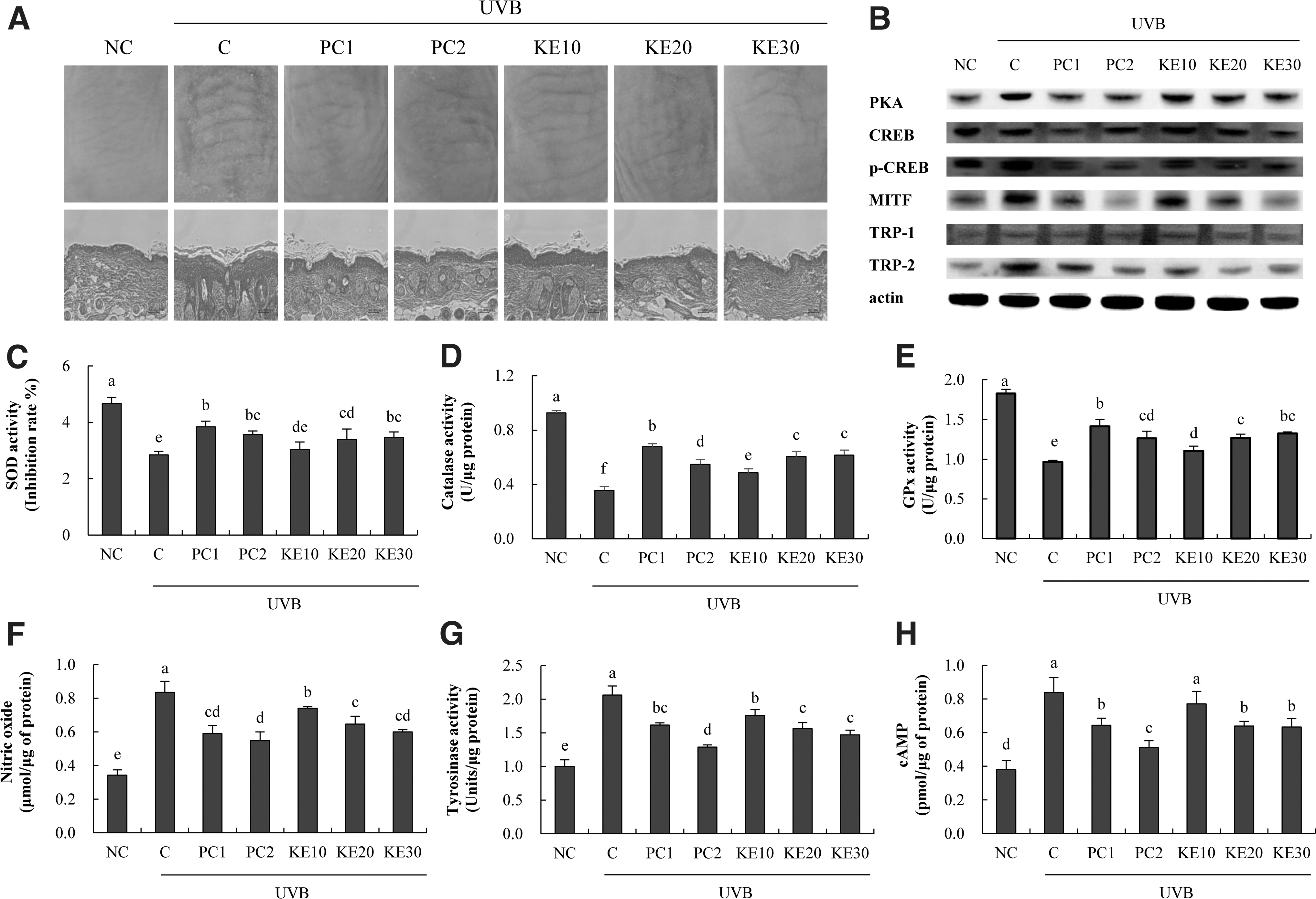
Effects of KE on wrinkle formation, skin thickness, antioxidant activity, and whitening activity in UVB-irradiated SKH-I hairless mice. H&E staining, western blotting, and ELISA were performed.
Effects of KE on factors related to wrinkle suppression in UVB-irradiated SKH-I hairless mice
Determination of the mRNA expression related to wrinkle suppression revealed that KE intake increased TGFβR1 expression (37.6% min to 189.9% max), procollagen type I expression (83.7% max), and collagen type I expression (26.8% min to 94.9% max) compared with levels in the C group (P < .05; Fig. 5I–K). KE intake decreased p-JNK/JNK protein expression (30.5% min to 36.0% max), p-c-Fos/Fos expression (37.3% min to 49.0% max), p-c-Jun/Jun expression (30.3% min to 37.8% max), MMP-1 expression (11.8% min to 29.5% max), MMP-3 expression (30.6% max), MMP-9 expression (36.0% max), and p-Smad3/Smad expression (20.8% min to 78.2% max) compared with those in the C group (P < .05; Fig. 5H). These markers acting on wrinkle-related mechanisms were similar to the in vitro experimental results.
Effects of KE on factors related to whitening activity in UVB-irradiated SKH-I hairless mice
In terms of protein expression, KE intake decreased the expression of PKA (18.6% max), p-CREB/CREB (19.2% min to 29.5% max), MITF (23.4% min to 40.5% max), TRP-1 (15.3% min to 33.1% max), and TRP-2 (66.8% max) compared with their levels in the C group (P < .05; Fig. 6B). Moreover, KE decreased NO levels (11.3% min to 28.1% max), tyrosinase activity (14.6% min to 28.6% max), and cAMP levels (8.2% min to 24.4% max) relative to those in the C group (P < .05; Fig. 6F–H). The whitening activity of KE was confirmed by the activity of the cAMP-PKA pathway, the activity of its submarkers CREB and MITF, and the change in the level of tyrosinase.
DISCUSSION
Following abnormal climate changes, destruction of the ozone layer increases the amount of UV radiation that reaches the surface of the earth, and skin lesions caused by UV rays have become one of the biggest concerns for people. 22 UV rays reduce moisture content, cause a loss of skin fibers, diminish elasticity, and cause wrinkles and pigmentation. 23 Excessive UV ray exposure can also cause skin cancers such as advanced melanoma of the skin, leading to the use of drugs with side effects. 24 Therefore, an interest in developing functional foods and cosmetics that do not have side effects that are beneficial for skin health is steadily increasing. 1
Elastin, an important protein of elastic fibers, provides elasticity to tissues, such as skin, blood vessels, and lungs, and is insoluble due to its crosslinked structure, high hydrophobicity, and high resistance to protein degradation. 25,26 Elastin affects the structure of the extracellular matrix and has a variety of physiological functions. 21 Recently, as the existence of elastin in bonito has become commonly known, it has been used to study various mechanisms related to photoaging by matching biological effects in relation to skin health. 27 It has been reported that small peptides of elastin hydrolyzed from bonito, also known as skipjack tuna, have free radical-scavenging ability. Because cell damage caused by UV rays is primarily due to the negative effect of ROS, materials with free radical-scavenging ability are promising as strong photoprotective agents. 27
Elastin from skipjack tuna has been used in various studies related to skin health, but more research is needed to address the mechanisms related to moisturization, wrinkle suppression, and whitening activity at the genetic and protein level as a whole. Moreover, desmosine and isodesmosine were detected in KE elastin. These are two of the most frequently found crosslinking amino acids in elastin, and they play an important role as crosslinking agents that bind the polymer chain of amino acids and could be a criterion to evaluate the purity and concentration of elastin. 28,29
In this study, KE increased the factors related to skin moisturization, such as LCB1, DEGS1, elastin, HAS2, CerS4, hyaluronic acid, and sphingomyelin, in UVB-irradiated HaCaT cells. LCB1, also called serine palmitoyltransferase, is an enzyme sequentially involved in the synthesis of ceramide along with DEGS1 and CerS4. Moreover, HAS2 is an enzyme primarily involved in the synthesis of hyaluronic acid. 30,31 In addition, in our previous study, the HS27 cell line was used to study photoaging.
In this study, we confirmed that KE increased the expression of the collagen synthesis factors, TGFβR1, procollagen type I, and collagen type I, in the UVB-treated HS27 cell model compared with levels in the UVB-treated controls. In addition, KE downregulated the protein expression of p-JNK, pc-fos, pc-jun, MMP-1, MM-3, MMP-9, and p-Smad3 in this cell model compared with their levels in the UVB control group, and proinflammatory levels of the cytokines IL-1β, IL-6, and TNF-α were also reduced. These are associated with a variety of diseases and are known as proinflammatory cytokines. 32 –34
IL-1β is a major mediator of inflammation, inducing fever and activating immune cells to exert their proinflammatory and antimicrobial effects. 35 IL-6 is a proinflammatory cytokine that stimulates specific immune reactions, hematopoiesis, and acute-phase responses. It has been reported that the level of IL-6 changes in various inflammatory diseases and rheumatoid arthritis. 36,37 TNF-α, a multifunctional proinflammatory cytokine, is produced by macrophages or monocytes during acute inflammation and plays a key role in regulating the cytokine cascade in several inflammatory diseases. 38
It is known that skin damage caused by UV ray irradiation increases the activity of MMPs that decompose collagen and other fiber components of the dermis. 8,39 MMPs are enzymes involved in the breakdown of cell substrates and exist in various forms. Among them, MMP-1 initiates the decomposition of collagen types I and III, and the MMP-3 form decomposes collagen type IV and promotes the activity of pro-MMP-1. In addition, the MMP-9 conformation further promotes the degradation of collagen fragments. 3,40
In the present study, in the B16F10 cell model induced to undergo IBMX-mediated melanogenesis, KE decreased the melanin content, tyrosinase activity, and NO and cAMP levels compared with those in the melanogenesis-induced control. In addition, KE increased the level of glutathione and regulated the protein expression of PKA, p-CREB, MITF, and tyrosinase-related proteins (TRPs). Activation of cAMP activates PKA and CREB, and NO activates cGMP through guanylate cyclase. These two pathways lead to increased MITF expression and increased tyrosinase activity. They play an important role in melanin synthesis.
Melanin is synthesized into brownish pheomelanin or red eumelanin as the activation of tyrosinase in melanocytes leads to an increase in L-3, 4-dihydroxyphenylalanine (L-DOPA) as well as in TRP 1 and 2. 41 –44 Apart from the L-DOPA pathway, there is also a pathway that affects the production of pheomelanine via cysteinyldopa under the influence of glutathione increase and glutathione-s-transferase and gamma-glutamyl transferase. 45 Tyrosinase is an important enzyme in the production of melanin and has been reported to be a rate regulator catalyzing the initial stage. 46
In vivo, KE intake increased skin hydration by increasing the gene and protein expressions of enzymes involved in the production of ceramide and hyaluronic acid in the skin of SKH-I hairless mice. From the skin morphological observation results, it was confirmed that the intake of KE significantly reduced skin wrinkles and thickness, increased the activity of antioxidant enzymes, and regulated gene and protein expression of factors involved in collagen synthesis. In addition, in terms of whitening activity, KE intake regulated related factors involved in melanin synthesis.
In this study, we investigated whether KE could affect skin dryness, wrinkles, and pigmentation in several cell models, and in SKH-I hairless mice. We found that KE exerted a positive effect on skin health by regulating factors involved in skin moisturization, antiwrinkle processes, and whitening activity in keratinocytes, dermal cells, melanocytes, and in animal models stimulated with UVB light and IBMX (only melanocytes). Compared with previous studies, these results are significant in that they are not only related to moisturization and wrinkle suppression, but the indicators involved in whitening activity were also confirmed. Moreover, we conducted a deep mechanistic study.
The results of this study, show that KE aids in moisturization improvement, wrinkle suppression, and increased whitening activity in the skin in vitro and in vivo. In particular, the amounts of desmosine and isodesmosine detected in KE are expected to be competitive compared with concentrations in other similar elastins in the future. Therefore, KE could be used in the cosmetics or functional food products aimed at improving general skin health.
Footnotes
AUTHOR DISCLOSURE STATEMENT
No competing financial interests exist.
FUNDING INFORMATION
No funding was received for this article.