
Abstract
Obesity may cause behavioral alterations, while maternal obesity can contribute to metabolic disorders in subsequent generations. The effect of β-glucan-rich Pleurotus pulmonarius (βgPp) was investigated on mouse neurobehavior and hippocampus and its offspring's hippocampus development. Female ICR mice were fed with normal diet (ND), ND with βgPp, high-fat diet (HFD), or HFD with βgPp for 3 months followed by behavioral test and mating. Immunohistochemistry for the expression of neuronal nuclear protein (NeuN) and ionized calcium binding adaptor molecule-1 (Iba-1) in the hippocampus was carried out. βgPp significantly enhanced short-term object recognition memory in HFD-fed mice. βgPp also ameliorated the histological alterations and neuronal loss and increased Iba-1-positive microglia in the hippocampus regions of HFD-fed mice and their male offspring. These findings demonstrated that βgPp supplementation attenuated the effects of HFD on object recognition memory and the alterations on the hippocampal regions of maternal mice and their male offspring.
Introduction
Obesity is one of the major threats to human health in the 21st century. 1 Aside from a sedentary lifestyle and imbalance in energy consumptions, obesity is also related to the intake of food rich in fats, mainly saturated fats. 2 Studies in both humans and rodents substantiated the profound effect of high-fat diet (HFD) on biobehavior such as mood and cognition, particularly those that are hippocampus dependent. 3,4 HFD-induced anxiety and impairment in spatial and recognition memory performance have been reported in rodent studies. 3,4 The decline in cognitive performance is associated with HFD-induced chronic inflammation in the brain region, particularly the hippocampus. 5
Maternal obesity has a long-lasting effect on the offspring. 6 As evidenced in human studies, maternal obesity is associated with neurodevelopmental disorders in the offspring such as attention deficit, cognitive impairment, anxiety and depression. 7,8 These effects are also observed in the offspring of HFD-fed dams in rodent studies, which provided further insight into the mechanism involved in neurodevelopmental impairment in the offspring such as alterations in brain structure and gene expression and inflammation. 7 Impairment in neuronal markers, suppressed neurogenesis, increased inflammation, and oxidative stress in the hippocampus of offspring born to HFD-fed dams were reported in various other studies. 9,10
Mushrooms have become the focus of international medicinal research as they exert many medicinal properties such as antiobesity, 11 antioxidant, 12 immunomodulatory, 13 and neuroprotective abilities. 12 Pleurotus pulmonarius (Fr.) Quél. (grey oyster mushroom) is one of the most commonly cultivated mushrooms. Previous studies have documented the beneficial effects of β-glucan from P. pulmonarius against the adverse effects of obesity. It has been reported to prevent weight gain and hyperlipidemia, as well as possess antidiabetic, antioxidant, and anti-inflammatory activities. 14,15
However, to the best of our knowledge, the ability of β-glucan from P. pulmonarius (βgPp) to improve learning and memory performance and the effects of βgPp supplementation on obese maternal mice toward its offspring's hippocampus morphology have not been reported. Therefore, this study was conducted to further assess the effects of βgPp on cognitive performance and against the effect of gestational obesity on offspring's hippocampus.
Materials and Methods
Mushroom collection and isolation of βgPp
Pleurotus pulmonarius (strain PL27) was purchased from Nas Agro Farm, Jenderam Hulu, Selangor, Malaysia (University of Malaya repository: UMP002). The isolation of βgPp was carried out by adapting the protocol from a previous study. 15 Freeze-dried powder of P. pulmonarius was boiled in distilled water (8 h), cooled overnight, and centrifuged (1500 g, 45 min, 4°C). The supernatant was precipitated in ethanol (1:5 ratio) and the resultant polysaccharides were freeze-dried, dissolved in distilled water, and dialyzed against distilled water for 4 h. The aqueous solution was collected and freeze-dried to yield 37.96% (g/100 g dm) of βgPp (81.56% of all glucan content).
Animals and experimental design
The protocols were approved by Institutional Animal Care and Use Committee, Faculty of Medicine, University of Malaya (Ethics no: 20180210605/ANAT/R/RPS). Eighteen female ICR mice 7 weeks old (weighing: 24–25 g) were purchased and housed at the Animal Experimental Unit, University of Malaya. Mice were allowed free access to food and water and were maintained at a standard condition (18–23°C, 12-h light/12-h dark cycle, 50–70% relative humidity) in polypropylene cages over corn cob bedding. After acclimatization, mice were randomly assigned into four groups (n = 6) as follows: Normal diet (ND)+distilled water control group (ND: 11% fat, 65% carbohydrate, and 24% protein, Altromin #1320) ND+240 mg/kg β-glucan of P. pulmonarius (ND+βgPp) HFD+distilled water control group (HFD: 60% fats from lard, 24% carbohydrate, and 16% protein, Altromin #C1090-60) HFD+240 mg/kg β-glucan of P. pulmonarius (HFD+βgPp)
Oral administration of βgPp to the treatment group was performed five times a week and the dose was selected based on a previous study. 15 After 3 months on the treatment, female mice were housed with adult male mice overnight to facilitate mating. Upon successful mating, pregnant females were housed individually. Treatments were continued throughout gestation and lactation until weaning. Weight and food intake were measured daily. The offspring were culled to 8–10 per dam with equal sex ratio where possible, and upon weaning, offspring were switched on to ND. Maternal mice were euthanized on postnatal day (PD) 21. As studies suggested males encountered more negative effect of maternal obesity than females, 9 one male offspring per dam was euthanized for hippocampus histological analysis on PD14, 21, and 28.
Novel object recognition test
Assessment of object recognition memory was carried out through the novel object recognition test (NORT) by adapting the protocol from a previous study. 16 During the acquisition phase, each mouse was allowed to explore two identical objects (A) in the open field arena for 5 min. An hour later (1-h NORT), each mouse was exposed to the familiar object (A) and a novel object (B) for 5 min for short-term memory assessment. After twenty-four hours (24-h NORT), each mouse was introduced to the familiar object (A) and a new novel object (C) for 5 min for long-term memory assessment. All tests were recorded by an overhead camera. The preference index (novelty preference) was calculated as follows:
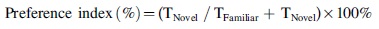
Histopathological analysis
At the end of the experiment, mice were perfused with 10% neutral buffered formalin under anesthesia. The brain tissues were processed in an automated tissue processor, Thermo Scientific Citadel 2000 (Fisher Scientific UK Ltd.), embedded in paraffin, and sectioned at 5-μm thickness using a rotary microtome, Leica 2045 Multicut (Leica Biosystems, Wetzlar, Germany). The tissue sections underwent routine histological processing and were stained with hematoxylin and eosin (H&E) for general microscopic assessment using Nikon Eclipse 80i, equipped with NIS element software (Nikon, Japan).
Immunohistochemical analysis
After deparaffinization, 5-μm thick hippocampus coronal sections underwent antigen retrieval with citrate buffer (pH 6.0) for 20 min at 95°C followed by incubation with hydrogen peroxide and protein blocking for 10 min at room temperature (cat. no. ab64264). Then the sections were incubated overnight at 4°C with primary antibodies diluted in tris-buffered saline with 1% bovine serum albumin: rabbit monoclonal anti-neuronal nuclear protein (NeuN) antibody, neuron marker (diluted 1:1500, cat. no. ab177487), and rabbit monoclonal anti-Iba1 antibody, microglia marker (diluted 1:2000, cat. no. ab178847).
After washing with TBS +0.025% triton X-100, sections were labeled with biotinylated goat anti-polyvalent antibody and then incubated with streptavidin peroxidase (cat. no. ab64264) for 10 min. Sections were developed with 3,3′-diaminobenzidine, DAB (cat. no. ab64264). All the antibodies and DAB detection kits were from Abcam (Cambridge, UK).
The percentage of NeuN-positive cells (number of immunoreactive cells/total number of hematoxylin-stained cells × 100%) 17 and the number of ionized calcium binding adaptor molecule-1 (Iba-1)-positive microglia cells were measured manually 18 with QuPath-0.2.3 software.
Statistical analysis
Data were presented as mean ± standard error mean of n = 6. Analysis and presentation of data were evaluated using GraphPad Prism version 9.0 (San Diego, CA, USA). The statistical differences between treatment groups were analyzed with one-way analysis of variance (ANOVA) followed by Tukey's post hoc test. Differences between novel and familiar object exploration time for NORT were analyzed by two-way ANOVA and Sidak's post hoc test. P Values <.05 were considered statistically significant.
Results
Effects of βgPp on body weight and food intake of maternal dams
There were no significant differences in the initial weight of mice among all treatment groups. However, after 90 days, HFD-fed mice had a higher body weight (BW), but not statistically significant compared to ND-fed mice (Table 1). On day 90, HFD-fed mice recorded a 45.11% ± 8.91% increase in BW from the initial BW, which was significantly higher than ND group (21.25% ± 3.83%). βgPp supplementation to HFD-fed mice reduced the BW gain to 41.35% ± 6.11%, but it was not statistically significant compared to HFD group. The average food consumptions were significantly (P < .001) reduced by HFD feeding and βgPp supplementation compared to ND-fed mice. The differences between ND group and ND+βgPp group were nonsignificant.
Effects of Pleurotus pulmonarius β-glucan on Maternal Body Weight, Weight Gain, and Food Intake
Data were obtained at the beginning of the experiments and on day 90, before mating. Values expressed as mean ± SEM (n = 6). Statistical analysis was performed using one-way ANOVA followed by Tukey's post hoc test.
P < .001 versus ND group.
βgPp, Pleurotus pulmonarius β-glucan; ANOVA, analysis of variance; BW, body weight; HFD, high-fat diet; ND, normal diet; SEM, standard error mean.
Effects of βgPp on object recognition memory of maternal dams
HFD group showed a slight deficit in short-term object recognition memory (1-h NORT) retention observed by a decrease in the novel object preference index compared to ND group from 60.66% ± 5.51% to 52.75% ± 4.36% (Fig. 1A). In contrast, βgPp supplementation to HFD-fed mice significantly improved short-term object recognition memory with a preference index of 69.46% ± 3.60%. Nonsignificant differences of novel object preference index were observed between both ND-fed mice groups. As observed in Figure 1B, only HFD+βgPp group explored the novel object significantly (P < .01) longer than the familiar object.
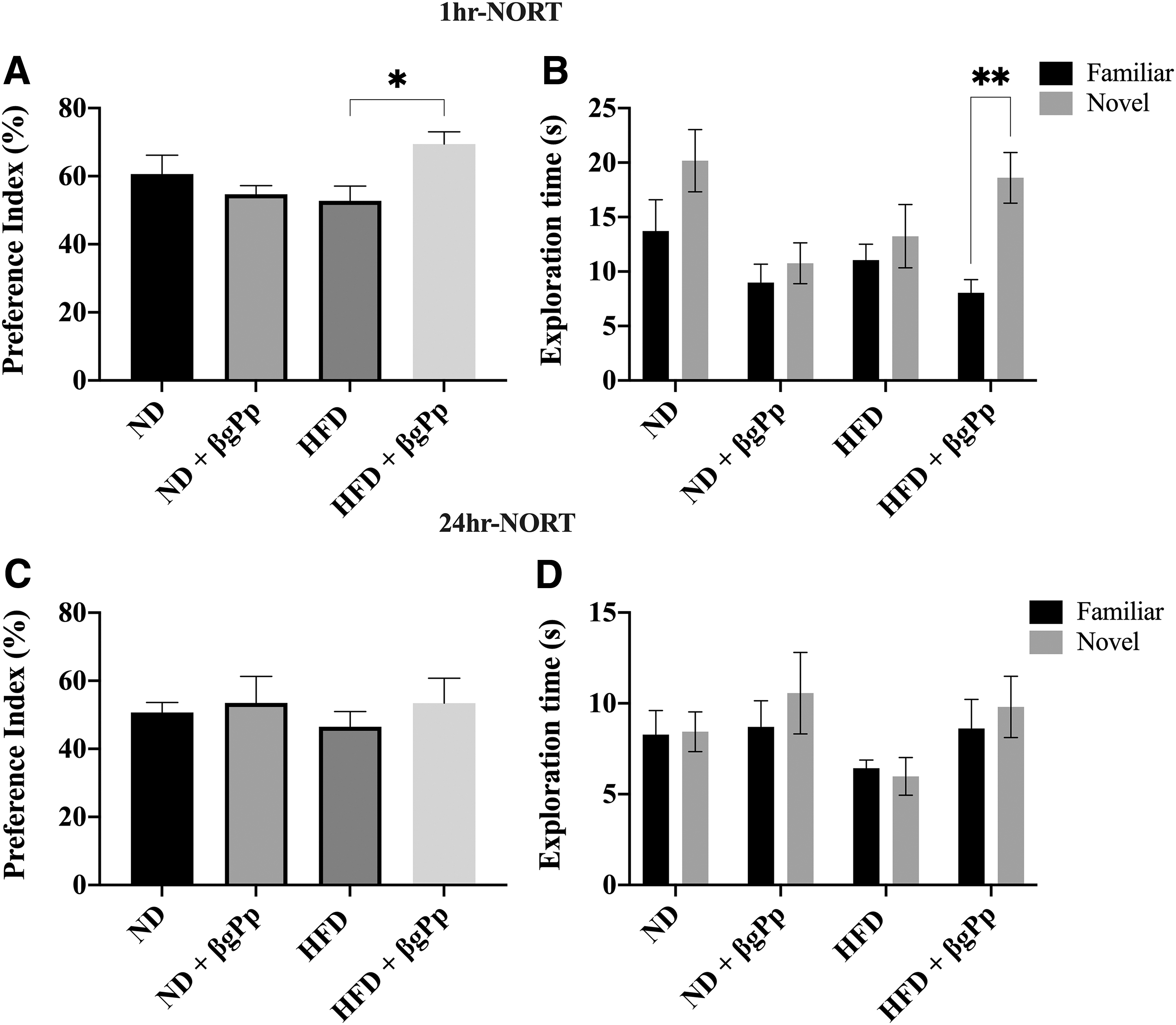
Effects of βgPp supplementation on object recognition memory. Object recognition memory was analyzed by NORT for short-term memory (1-h NORT) and long-term memory (24-h NORT).
However, for long-term object recognition memory (24-h NORT), although βgPp administration to HFD-fed group increased the novel object preference index (53.43% ± 7.32%), the difference was not significant (P > .05) compared to HFD group (46.52% ± 4.45%). Overall, no significant (P > .05) differences were observed across all groups for preference index (Fig. 1C) and object exploration time (Fig. 1D) for 24-h NORT.
Effects of βgPp supplementation on maternal hippocampus
The granule cell layers of the dentate gyrus (DG) and pyramidal cell layer neurons of cornu ammonis 1 (CA1) and cornu ammonis 3 (CA3) regions remained intact for ND group. As illustrated in Figure 2A, a higher intensity of degenerating neurons (shrunken neurons with pyknotic/hyperchromatic nuclei) was observed at the DG and CA3 regions of HFD group. Nonetheless, these neuronal losses were ameliorated by βgPp supplementation with fewer pyknotic nuclei observed throughout all hippocampus regions. Immunohistochemical analysis of NeuN demonstrated a significantly lower percentage of NeuN-positive cells at DG (P < .001) and CA1 (P < .05) regions of HFD group compared to ND group (Fig. 2B, D).
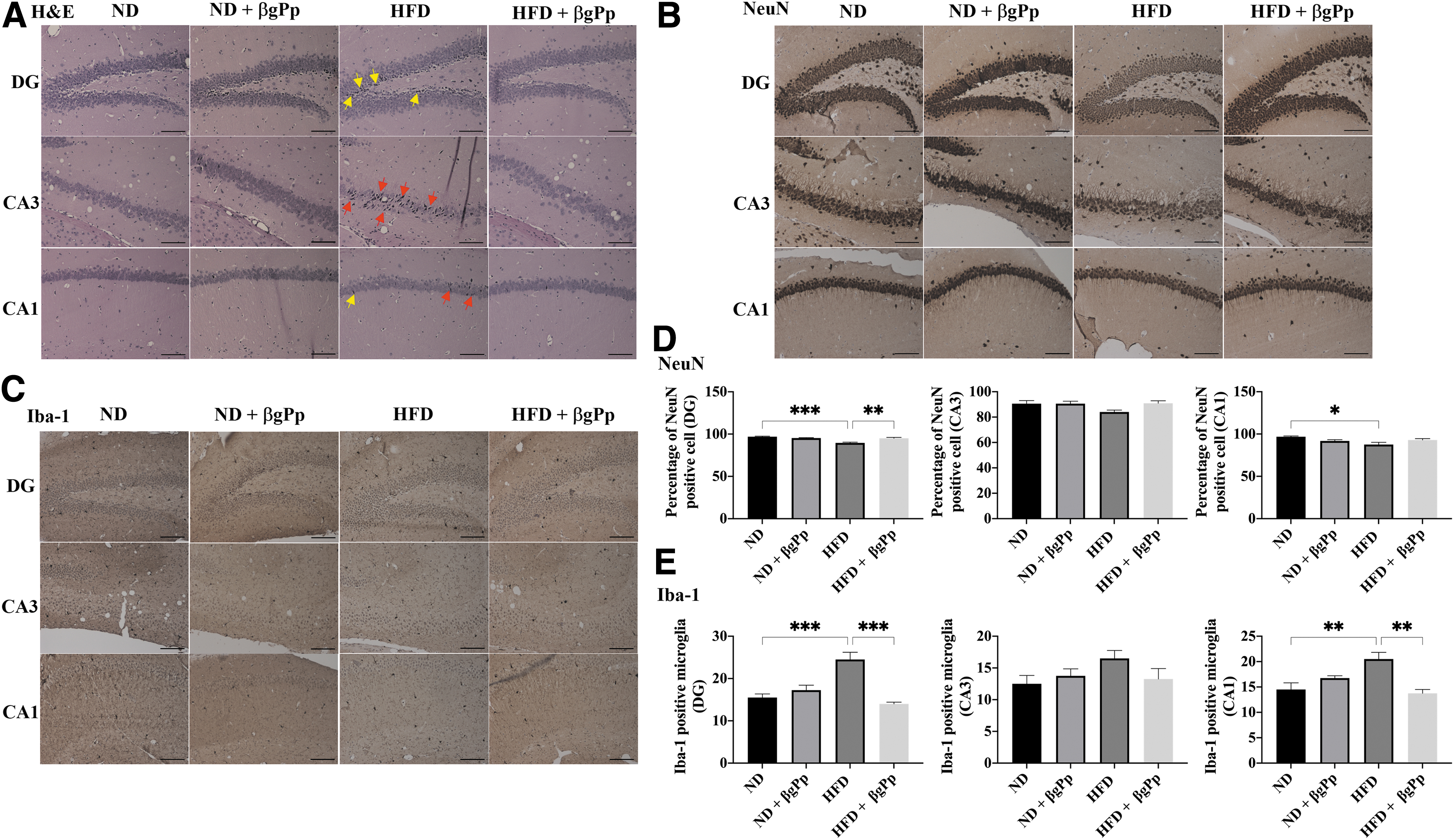
Effects of βgPp supplementation on maternal hippocampus.
Analysis of Iba-1 expression showed significantly higher counts of Iba-1-positive microglia at DG (P < .001) and CA1 (P < .01) regions of HFD group than ND group (Fig. 2C, E). βgPp supplementation was able to significantly ameliorate the loss of neurons (DG, P < .01) and significantly reduced the number of Iba-1-positive microglia in HFD-fed mice hippocampus (DG, P < .001; CA1, P < .01). The differences between ND group and ND+βgPp group were nonsignificant.
Effects of βgPp on histomorphology of male offspring hippocampus
Early morphological changes due to maternal HFD feeding were observed at the hippocampus of PD14 and PD21 male offspring. As illustrated in Figures 3A and 4A, a higher intensity of pyknotic/hyperchromatic nuclei was visualized at the DG and CA3 regions of offspring born to HFD dams compared to offspring born to ND dams and HFD+βgPp dams.
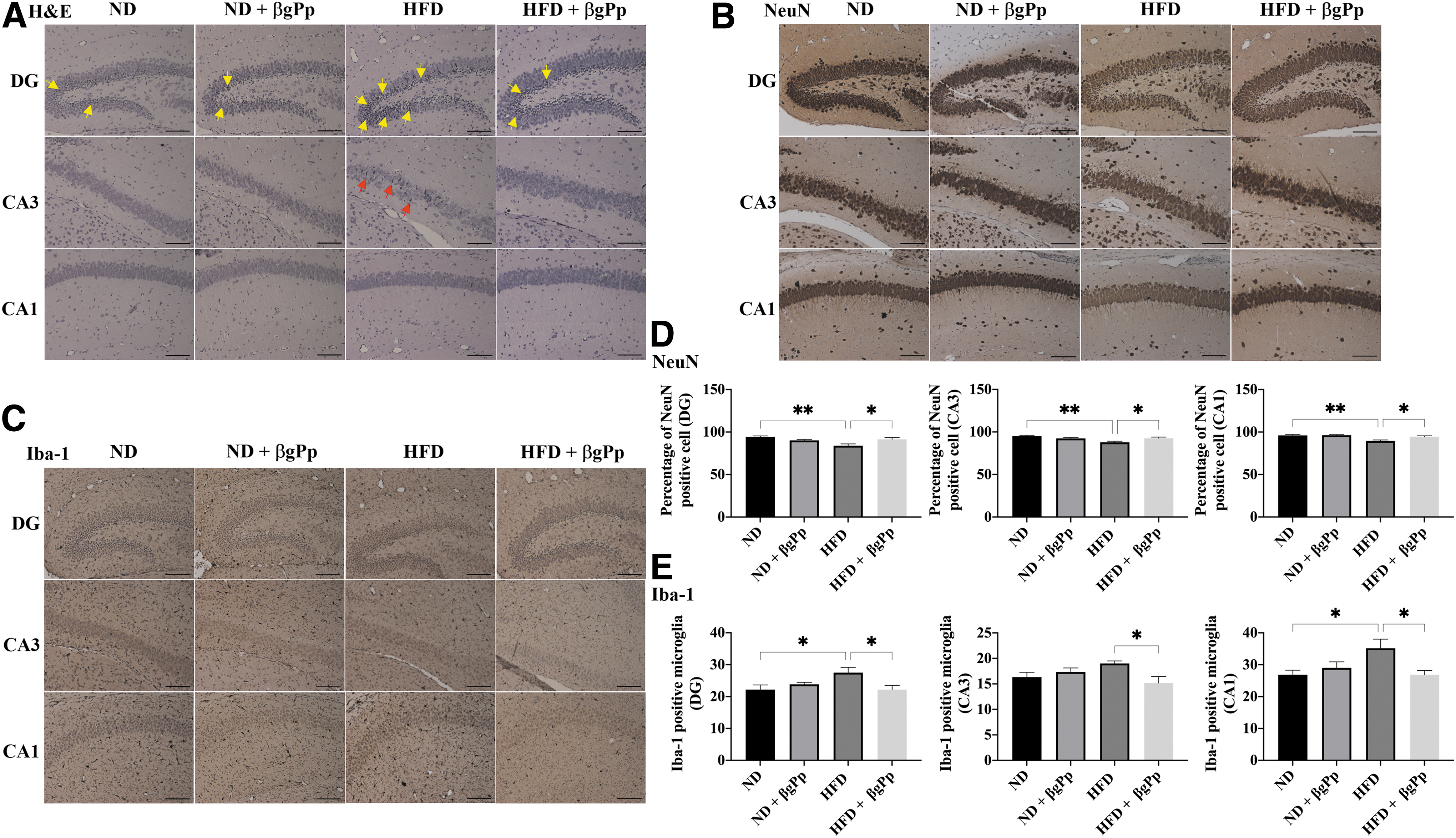
Effects of βgPp supplementation on hippocampus morphology of offspring on PD14.
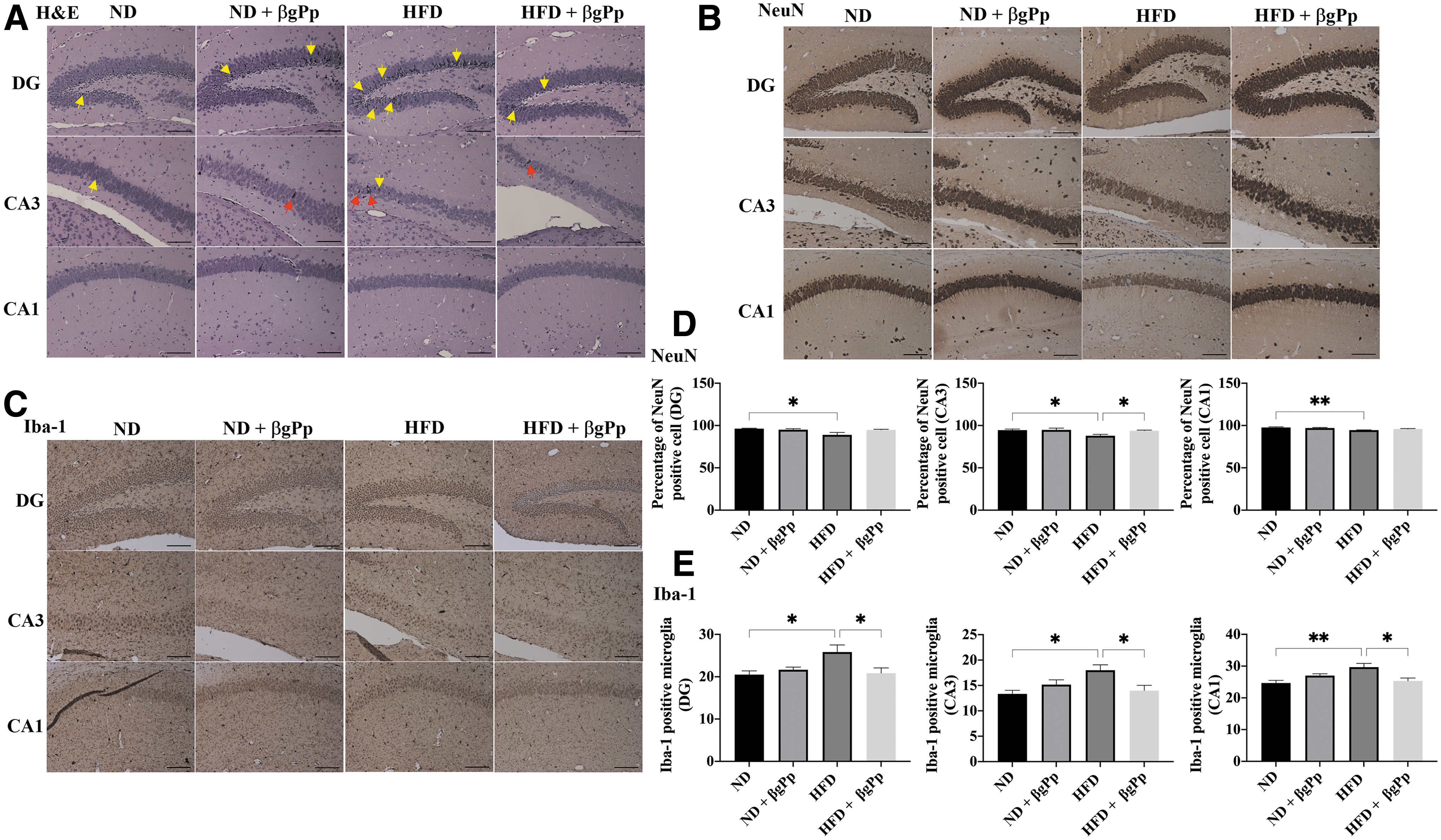
Effects of βgPp supplementation on hippocampus morphology of offspring on PD21.
The percentage of cells expressing NeuN marker was significantly (P < .01) reduced in the hippocampus regions of PD14 offspring belonging to HFD dams (Fig. 3B, D), which were significantly increased in the HFD+βgPp offspring hippocampus. A significant reduction of NeuN-positive cells in PD21 offspring of HFD dams (Fig. 4B, D) was noted at DG, CA3 (P < .05), and CA1 (P < .01) regions and was significantly elevated, particularly in the CA3 region of offspring belonging to HFD+βgPp dams.
A similar trend was observed for Iba-1-positive microglia. A significantly higher count of Iba-1-positive microglia was observed at the hippocampus (Fig. 3C, E) of PD14 offspring of HFD dams (DG and CA1, P < .05) and hippocampus (Fig. 4C, E) of PD21 offspring born to HFD dams (DG, CA3, P < .05; CA1, P < .01) compared to offspring of ND dams. Nonetheless, these were significantly (P < .05) reduced in the hippocampus of offspring of HFD+βgPp dams on PD14 (Fig. 3E) and PD21 (Fig. 4E). Nonsignificant differences were observed between offspring born to ND dams and ND+βgPp dams.
Upon weaning of all offspring to ND, no significant changes were observed in H&E-stained sections (Fig. 5A) and NeuN-positive cells counts (Fig. 5B, D) in the hippocampus of male offspring for all groups on PD28. However, Iba-1 analysis (Fig. 5C, E) on PD28 demonstrated an increase of Iba-1-positive microglia at the hippocampus regions of offspring born to HFD dams, and the count was slightly reduced in the hippocampus of offspring of HFD+βgPp dams.
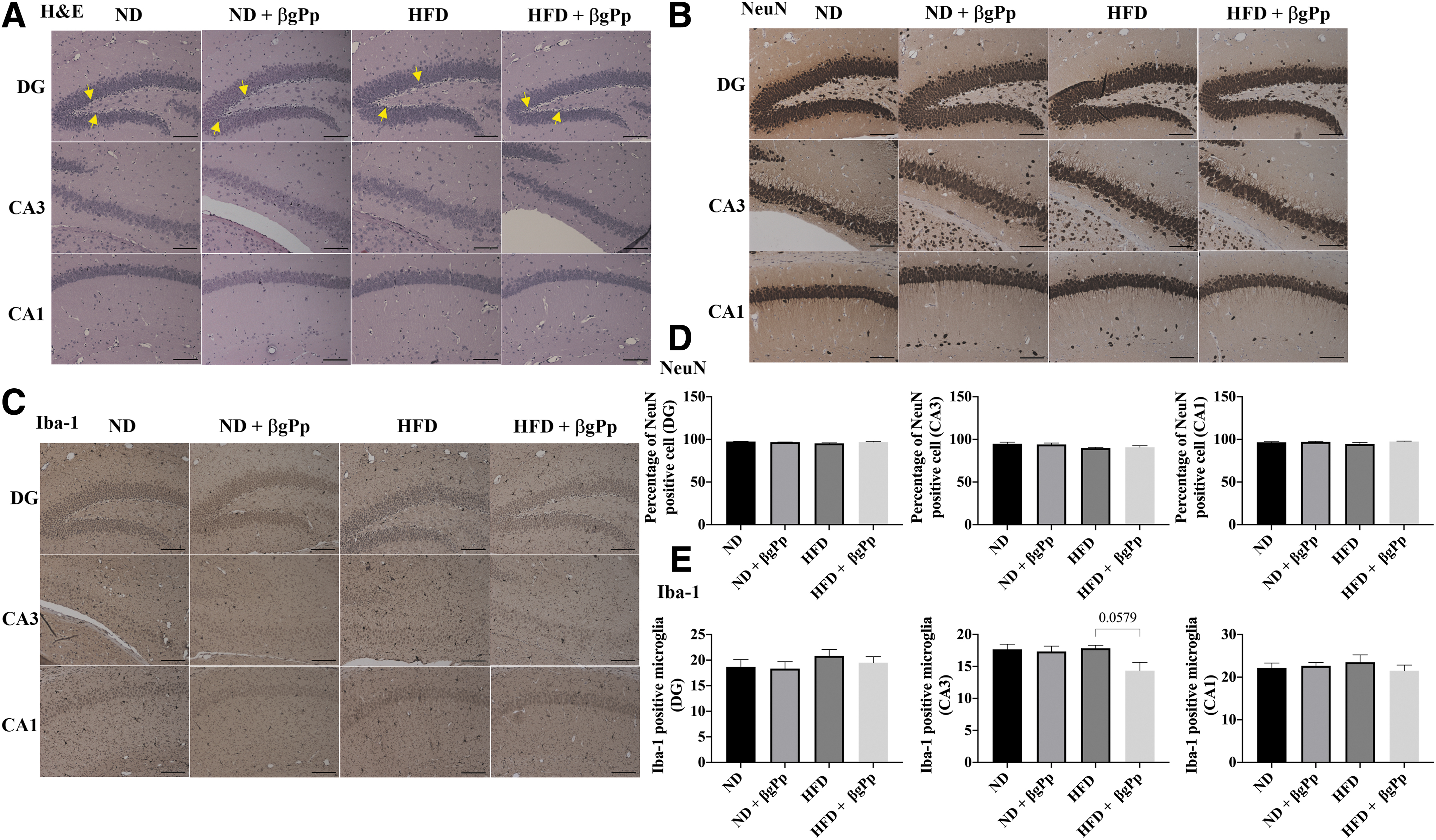
Effects of βgPp supplementation on hippocampus morphology of offspring on PD28.
Discussion
Obesity prevalence worldwide is alarming and its association with other severe health conditions is detrimental to humans. 6 Concerns are on the rise regarding the effects of maternal obesity on their offspring's health and development. Accumulating evidence has linked maternal obesity with the risk of obesity, metabolic diseases, and neuropathological changes in the offspring. 19
The BW gain of HFD-fed mice was approximately doubled compared to those of ND-fed mice and this may indicate that HFD-fed mice accumulated body fats easily since HFD is known to cause adipocyte hypertrophy and hyperplasia. 20 βgPp supplementation to HFD-fed mice was able to reduce the BW gain consistent with our previous study. 15 βgPp also reduced food intake of HFD-fed mice in this study, which could be due to the solubility and viscosity properties of β-glucan. Soluble fibers with viscosity properties found in β-glucan can induce satiety and prevent an increase in appetite, which aid in slowing down nutrient absorption and delaying gastric emptying. 21
Mounting evidence has demonstrated the deleterious effects of HFD on object recognition memory. 4,22 However, in this study, HFD only caused a slight reduction on object recognition memory. This was independent of the HFD effect on BW as HFD-fed mice did not exhibit locomotor activity deficits. Nonetheless, it is novel to note that βgPp supplementation resulted in a significant improvement of object recognition memory in HFD-induced obese mice, suggesting the ability of βgPp to reverse the adverse effects of HFD.
It has been proven in rodent studies that HFD induces inflammation and apoptosis of hippocampal neurons, which disrupt cognitive function. 5,23 In this study, we observed neuronal degeneration in the hippocampus of HFD-fed mice. Interestingly, the supplementation of βgPp was able to prevent morphological changes caused by HFD. This finding is also evidenced with analysis of NeuN immunohistochemistry. We observed a significant reduction of NeuN immunoreactivity in the HFD group hippocampus, which is consistent with previous studies. 24,25 Nonetheless, this effect was reversed with βgPp supplementation, suggesting its neuroprotective effect.
Our analysis for Iba-1-positive microglia on the hippocampus of maternal mice suggested neuroinflammation as a possible mechanism for the NeuN-positive cell reductions. HFD has been shown to increase microgliosis in the hippocampus, which is indicative of inflammation. 26,27 Microglia is activated in response to pathogenic insults; however, excessive activation may worsen the inflammation. 28 We detected a significant microgliosis in the HFD-fed mice hippocampus. This was significantly reduced by βgPp supplementation, demonstrating its anti-neuroinflammatory activity, which may contribute to the protection of hippocampal neurons.
Recent studies have demonstrated the neuroprotective effects of β-glucan on the brain tissue are mediated by the gut-brain axis by improving gut microbiota composition. Fermentation of β-glucan in the colon elevated Bacteroidetes phylum levels, which then triggered the production of short-chain fatty acids. This may affect neuroinflammation, besides its involvement in the regulation of neurotransmitters and neurotrophic factors, which also influence neurological function. 29,30
β-glucan also confers neuroprotection by its anti-inflammatory activities as studies have shown its inhibitory action on microgliosis and decreased expression of proinflammatory cytokines such as interleukin (IL)-6, tumor necrosis factor-alpha (TNF-α), and IL-1β. 31 In a diabetic rat study, β-glucan showcased its antioxidant activity by reducing MDA concentrations and increased catalase levels in the brain tissue, contributing to its neuroprotective capability. 32
According to our findings, βgPp supplementation to ND-fed mice was nontoxic to the hippocampus as observed in H&E-stained, NeuN, and Iba-1 hippocampus immunohistochemistry. Object recognition memory was also not affected by βgPp supplementation to ND-fed mice. However, βgPp showcased its neuroprotective capabilities against mice fed an HFD. Previous studies demonstrated that HFD may induce gut dysbiosis, which resulted in cognitive impairment, and β-glucan supplementation ameliorated those effects. 22,31 In another study, oat β-glucan was shown to prevent cognitive impairment, inhibited microglia activation, downregulated IL-6, TNF-α, and IL-1β mRNA expression, and improved Tau signaling in HFD-fed mice hippocampus through gut-brain axis. 22
Another recent study utilizing β-glucan from Shiitake mushroom, Lentinula edodes, in HFD-fed mice also demonstrated amelioration of cognitive impairment by inhibiting gut dysbiosis, microgliosis, upregulation of proinflammatory cytokines, and recovered BDNF and synaptic markers level. 31 Further studies are required to investigate whether βgPp utilizes the same mechanism, which resulted in the improvement of cognitive performance.
Maternal influences on the neural development of offspring may be mediated by several factors such as nutrition, infection, and stress. 33 Viral infections during pregnancy may induce inflammation, hence releasing proinflammatory cytokines, which eventually lead to offspring neurodevelopmental disorders. Maternal stress may also result in the same outcome due to fetal glucocorticoid overexposure, catecholamines, and kynurenic acid accumulation in the placenta and fetal brain, as well as altered oxytocin signaling and maternal microbiome. 33
However, within the scope of our study, nutritional factor was our main focus. Growing evidence has indicated nutritional insults such as maternal obesity influences the offspring's neural development. 19,34 Studies involving maternal obesity have demonstrated that inflammation and alteration in iron regulation and myelination in the offspring's brain may eventually lead to neurobehavioral impairment. 35
Maternal obesity has also been associated with the reduction of gene expression involved in neurodevelopment, learning, and memory such as glucose transporter, insulin, and leptin receptor in the offspring's hippocampus. 7 Impaired development of serotonergic and dopaminergic systems as well as hippocampal BDNF production have been observed in the offspring of obese mothers. 7 Other proposed factors that can affect offspring's neural development include placental dysfunction, inflammation, and alterations in obese maternal gut microbiome, followed by microbial transmission during prenatal and postnatal periods. 36,37
Hence, we explored the effects of maternal HFD feeding during pregnancy and lactation on the offspring's hippocampus. The hippocampus is vulnerable to early nutritional insult, and its’ crucial involvement in task performance, learning, and memory piqued our interest in making it our prime focus. 38 It is also worth noting that most hippocampal development in rodents occurred during the prenatal and early postnatal periods, 38 which coincided with the duration of HFD exposure toward maternal mice in our study.
Maternal HFD intake has been shown to impair neurogenesis and increase apoptosis in the male offspring's hippocampus. 39,40 Similar to the maternal counterparts, we observed a significant neuronal loss at the hippocampal regions of offspring from HFD dams. To the best of our knowledge, this is the first study that demonstrates the transmission of beneficial effect of βgPp supplementation from maternal mice to offspring's hippocampal development. The pathological changes in the hippocampal regions due to maternal HFD feeding were attenuated, as the offspring of HFD+βgPp dams have similar hippocampus morphology as the offspring of ND-fed dams.
The effect of HFD-induced inflammation displayed by a significant increase of Iba-1-positive microglia was observed in the hippocampus of male offspring from HFD-fed dams. Other studies have attributed the increase of Iba-1-positive microglia on the offspring hippocampus to maternal HFD feeding. 41,42 The antineuroinflammatory activity of supplemental βgPp in the HFD-fed maternal mice was also apparent in the hippocampi of offspring as a marked reduction in Iba-1-positive microglia.
The sample size used in this study was appropriate as it was based on the principles of 4Rs (replacement, reduction, refinement, and rehabilitation) in animal research. However, by increasing it, subsequent studies may present more significant effects of HFD and βgPp on maternal mice and their offspring. Nonetheless, this study demonstrated the potential beneficial effects of βgPp on obese mice and their offspring. Aside from reducing weight gain, βgPp supplementation also improved object recognition memory. Dietary supplementation of βgPp also ameliorated histopathological changes at the hippocampal region of obese maternal mice and their male offspring.
Data Availability Statement
The data that support the findings of this study are available from the corresponding author, P.D., upon reasonable request.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by Universiti Malaya under the Research University Grant, RU Grant [Project No: RF010C-2018], and the Ministry of Higher Education, Malaysia, for the Fundamental Research Grant Scheme, FRGS [Project No: FP008-2017A; MOSTI code: FRGS/1/2017/SKK08/UM/01/12; Reference ID: 11334].