
Abstract
Tamarixetin (TX) is an O-methylated flavonoid naturally derived from quercetin. TX has bioactive properties; however, whether it shows antilipogenic activity remains unknown. Therefore, in the present study, we aimed to determine the antilipogenic effects of TX using 3T3-L1 adipocytes. The 3T3-L1 adipocytes were cultured in a differentiation medium with or without TX. Lipid accumulation was diminished and the mRNA expression of lipogenesis-related genes was decreased following TX treatment. We found that TX exhibited antilipogenic effects by inhibiting the expression of p300/CBP-associated factor (pCAF), a histone acetyltransferase, as confirmed by pCAF knockdown. Furthermore, TX inhibited both pCAF expression and its activity, thereby reducing the total acetylation level of nonhistone and histone proteins. Finally, TX decreased the expression of CCAAT/enhancer-binding protein alpha and beta (CEBPα and CEBPβ), and peroxisome proliferator-activated receptor γ along with pCAF expression during adipogenesis of 3T3-L1 cells in a time-dependent manner. Collectively, our findings suggest that TX is a potent antilipogenic agent derived from natural products and may be used as a pCAF inhibitor.
Introduction
Obesity, which is primarily caused by excessive accumulation of triacylglycerol (TG) in adipose tissues, represents a major risk factor for the development of metabolic disorders, such as type 2 diabetes mellitus, nonalcoholic fatty liver disease, and cardiovascular disease. 1 –4 Several studies have investigated the association between obesity and cancer development, progression, and recurrence. 5,6 The consumption of bioactive food components derived from fruits and vegetables has been established as a possible approach for preventing obesity and various studies have shown that such components exert their effects by regulating diverse processes.
Epigenetic regulation is known to mediate the in vivo actions of bioactive food components. Epigallocatechin gallate, for instance, interacts with histone deacetylases (HDACs) and DNA methyltransferases (DNMTs), thereby affecting epigenetic modulation. 7 Apigenin decreases DNMT 8 and HDAC activities. 9 Curcumin is a potent modulator of DNMTs, HDACs, histone acetyltransferases (HATs), and microRNAs. 10 Several other bioactive food components, including genistein, 11,12 indole-3-carbinol, 13 lycopene, 14 quercetin, 15,16 resveratrol, 17 and sulforaphane, 18,19 have been reported to affect epigenetic regulation.
Adipogenesis involves a series of processes mediating the formation of adipocytes and it is closely linked to the development of obesity; various factors involved in these processes are also targets of epigenetic regulation. Peroxisome proliferator-activated receptor γ (PPARγ) and CCAAT/enhancer-binding protein α (C/EBPα), which have been established as key players in the early stage of adipogenesis, are regulated by epigenetic components. However, few genome-wide profiling studies have demonstrated that GCN5/p300/CBP-associated factor (PCAF)-mediated H3K9ac and CBP/p300-mediated H3K27ac are essential for the initiation of PPARγ transcription in adipogenesis. 20,21
These HATs, GCN5, PCAF, CBP, and p300, are responsible for the epigenetic regulation and acetylation of histone and nonhistone proteins. Although histone acetylation is a general marker of transcription, mediating a charge neutralization of DNA in the nucleosome, 22 recent studies indicate that recognition of acetylation in transcriptional proteins (nonhistone proteins) at lysine residues is more important than the change in charge itself. 23
In an ongoing screening study to identify phytochemicals showing anti-HAT activity, we identified tamarixetin (TX) as a novel HAT inhibitor. This natural monomethoxyflavone derived from a quercetin of Cyperus teneriffae has been reported to have antioxidant, 24 anti-inflammatory, 25 and anticancer 26 effects. However, there are no studies reporting the antiobesity effects of TX. Thus, in the present study, we investigated the effects of TX on adipocyte differentiation by analyzing the mRNA expression of several adipogenic genes, and evaluated the anti-HAT activity implicated in this process.
Materials and Methods
3T3-L1 differentiation and TX treatment
Mouse 3T3-L1 preadipocytes were obtained from the American Type Culture Collection (ATCC, Manassas, VA, USA). The cells (2 × 104/well) were cultured in a six-well plate in high-glucose Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% bovine calf serum and antibiotics (Welgene, Daegu, Korea) at 37°C in a humidified atmosphere with 5% CO2 until reaching 100% confluence (day 0). After 2 days, the cells were provided a differentiation medium containing the mixture of 3-isobutyl-1-methylxathine, dexamethasone, and insulin (MDI) (0.5 mM 3-isobutyl-1-methylxanthine, 1 μM dexamethasone, and 1 μg/mL insulin) with 10% fetal bovine serum (FBS) for differentiation of preadipocytes into adipocytes.
The differentiation medium applied to the cells was replaced with DMEM supplemented with 10% FBS and insulin (10 μg/mL) every 2 days until day 7. TX (ChemFaces, Wuhan, China) was treated in the cells at day 0 (TX-treated group), newly replaced through media changes, and then cells were harvested to the experiments at day 7.
Determination of the effect of the cytotoxic effect of TX on 3T3-L1 cells
At the end of differentiation (day 7), cellular cytotoxicity was measured using the 3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2H-tetrazolium bromide (MTT) assay (Sigma, St. Louis, MO, USA). Cells were exposed to 1 mg/mL MTT solution for 2 h at 37°C and then dissolved in dimethyl sulfoxide (DMSO). To determine cell viability, absorbance of this solution was measured at 570 nm (Molecular Devices, Sunnyvale, CA, USA).
Determination of the effect of TX on lipid accumulation
To evaluate the antiadipogenic effects of TX on 3T3-L1 differentiation, 3T3-L1 cells were treated with TX at 25 or 100 μM, at days 0, 2, 4, and 6 of the experiment with the media changed every 2 days. Differentiated adipocytes were washed with phosphate-buffered saline (PBS) and fixed in 4% formaldehyde. Cells were rinsed with distilled water and stained with 0.1% Oil Red O solution for 15 min at room temperature. Stained cells were washed with distilled water, and lipid droplets were observed. Cell images were captured under the microscope (Olympus IX51; Olympus corporation, Central Valley, PA, USA). Oil Red O dye was dissolved in isopropanol and the dissolved dye was used to measure absorbance at 510 nm (Molecular Devices).
Determination of the effect of TX on expression of adipogenesis-related genes
For quantitative reverse transcription polymerase chain reaction (qRT-PCR) assay, 3T3-L1 cells were seeded in six-well plates (2 × 104 cells/well). After adipocyte differentiation, as described above, cells were harvested and total RNA was isolated using TRIzol reagent (TaKaRa, Shiga, Japan). Complementary DNA was synthesized from 1 μg of RNA and qRT-PCR was performed using the iCycle iQ system (Bio-Rad, Hercules, CA, USA) with SYBR Green Master Mix (Roche, Basel, Switzerland). The qRT-PCR assays were performed in triplicate, and the primers used are listed in Supplementary Table S1. The relative mRNA expression levels were normalized to the expression of β-actin, which was used as the internal control, and analyzed by the comparative quantification method.
For western blotting, the 3T3-L1 cells were harvested and washed with cold PBS and extracted with lysis buffer (Cell signaling, Beverly, MA, USA) containing protease and phosphatase inhibitors (Roche). Total cell lysates were centrifuged at 20,000 g for 20 min at 4°C and adjusted to 15 μg for western blot analysis. Cell lysates were separated on sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) and then transferred onto nitrocellulose membranes.
After blocking in 5% nonfat skim milk, the membranes were incubated with the primary antibodies αp300 (Santa Cruz, CA, USA), αCBP (Santa Cruz), αpCAF (Santa Cruz), αC/EBPα (Cell signaling, Denvers, MA, USA), C/EBPβ (Cell signaling), αPPARγ (Cell signaling), αHSP90αβ (Cell signaling), and αAcLys (Cell signaling) (Supplementary Table S2). The membranes were visualized using a chemiluminescence reagent (Thermo Fisher Scientific, Waltham, MA, USA) by FUSION Solo S (Vilber Lourmat, ZAC de Lamirault, France).
Determination of the effect of TX on HAT activity in vitro
HAT activity was measured using a cell-free assay system (BioVision Biotechnology, Milpitas, CA, USA) and the HeLa cell nuclear extract (BioVision Biotechnology) as the enzyme source, according to the manufacturer's protocol. To evaluate the inhibitory effect of TX on specific HATs, pCAF (pCAF; Active motif, Carlsbad, CA, USA) recombinant enzyme was added instead of the nuclear extracts. IC50 values were calculated using Prism 8 software (GraphPad Software, Inc., CA, USA).
Determination of the effect of TX on histone acetylation
The 3T3-L1 cells were cultured and harvested as described above, and prepared for histone extraction according to the manufacturer's protocol (Abcam, Cambridge, MA, USA). Histone proteins were isolated from whole cells using the prelysis buffer and the balance buffer containing dithiothreitol (DTT). The histone extracts were subjected to western blotting to detect the levels of histone acetylation at H3K9, H3K27, and H3K36 using specific antibodies (Supplementary Table S2).
Statistical analyses
Data were analyzed using either Students' t-test or one-way analysis of variance with Tukey's multiple comparison test. The values were presented as mean ± SD. Statistical analyses were conducted using the SPSS (Version 20; SPSS, Inc., Chicago, IL, USA) or Prism 7 (GraphPad Software, Inc.) software. Statistical differences were considered significant at P < .05. Mean with different superscript letters are significantly different, P < .05.
Results
TX decreased lipid accumulation in 3T3-L1 adipocytes
We evaluated the effect of TX on lipid accumulation in 3T3-L1 cells, and TX (100 μM) treatment significantly reduced the formation of the MDI-triggered lipid droplets (Fig. 1A). Notably, lipid accumulation in the group treated with 100 μM TX was similar to that in the control group, to which MDI medium was not added. This result may be attributed to TX-induced cytotoxicity. Therefore, cell viability was measured at day 1 and at day 7 after the differentiation of 3T3-L1 cells under the same conditions (Fig. 1B). However, during the 3T3-L1 differentiation process, TX treatment did not induce cytotoxicity (Fig. 1B). Thus, TX effectively disrupted adipogenesis in 3T3-L1 cells without inducing cytotoxicity.
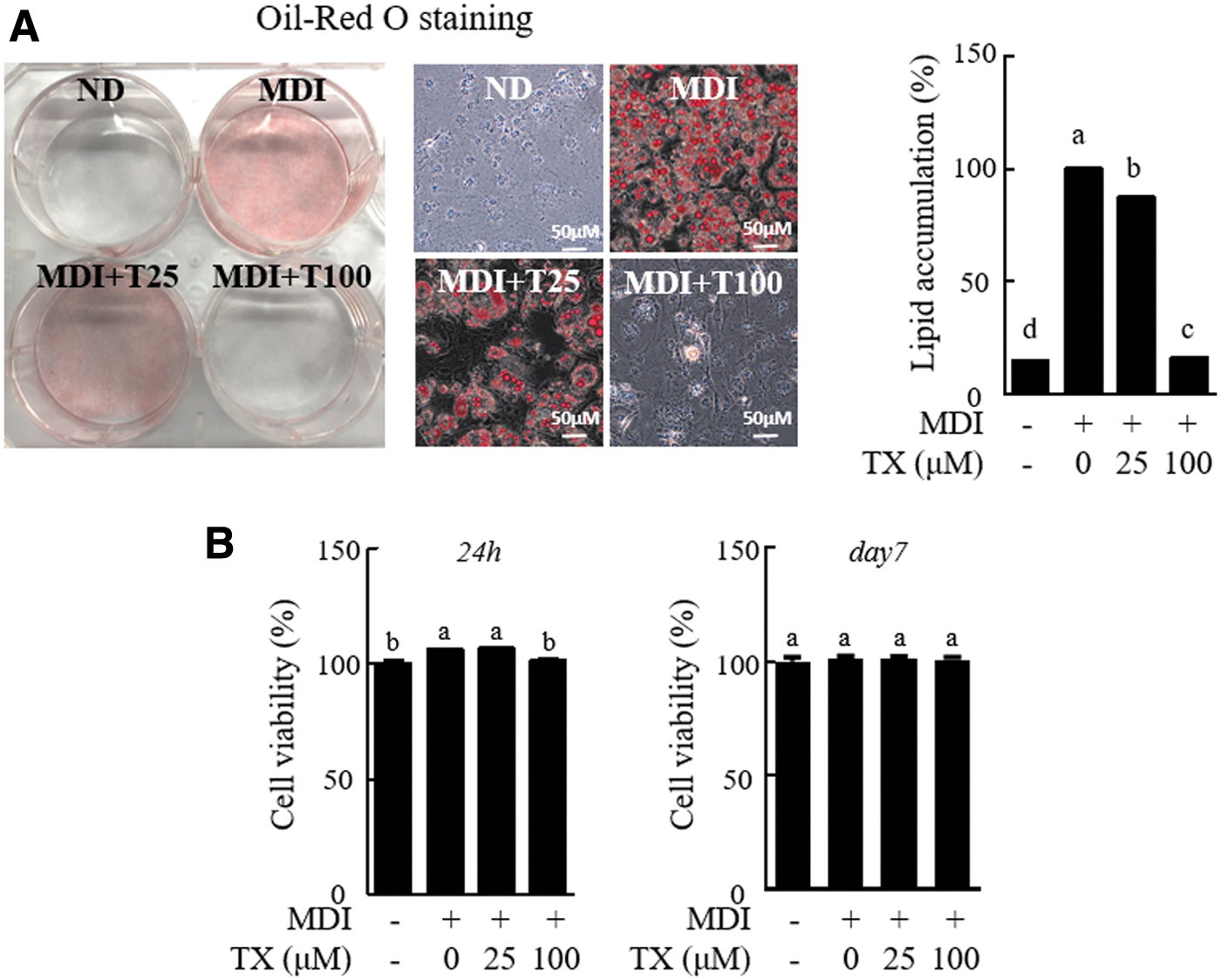
TX decreased lipid accumulation in differentiation of 3T3-L1 preadipocytes into adipocytes.
TX decreased the mRNA expression of adipogenesis-related genes
To determine the mechanism by which TX disrupted adipogenesis, we performed qRT-PCR for four functional groups of genes involved in lipid metabolism: transcription factor, fatty acid synthesis, TG synthesis, and lipid transport.
The mRNA expression of all genes, including transcription factors (C/EBPα, C/EBPβ, Pparγ1, Pparγ2, and liver X receptor α [LXRα]), fatty acid synthesis-related genes (fatty acid synthase [Fasn], ATP-citrate synthase [Acly], and acetyl-CoA carboxylase [Acc]), TG synthesis-related genes (glycerol-3-phosphate acyltransferase [Gpat], 1-acylglycerol-3-phosphate O-acyltransferase 2 [Agpat2], diacylglycerol acyltransferase 1 [Dgat1], and monoacylglycerol acyltransferase [Mogat1]), and lipid transport-related genes (fatty acid-binding protein [Fabp4], Cidec encoding fat-specific protein 27 [Fsp27], and [Cd36]) were reduced after TX treatment (Fig. 2). Thus, TX could downregulate key adipogenesis-related genes and adipogenesis regulators in 3T3-L1 cells.
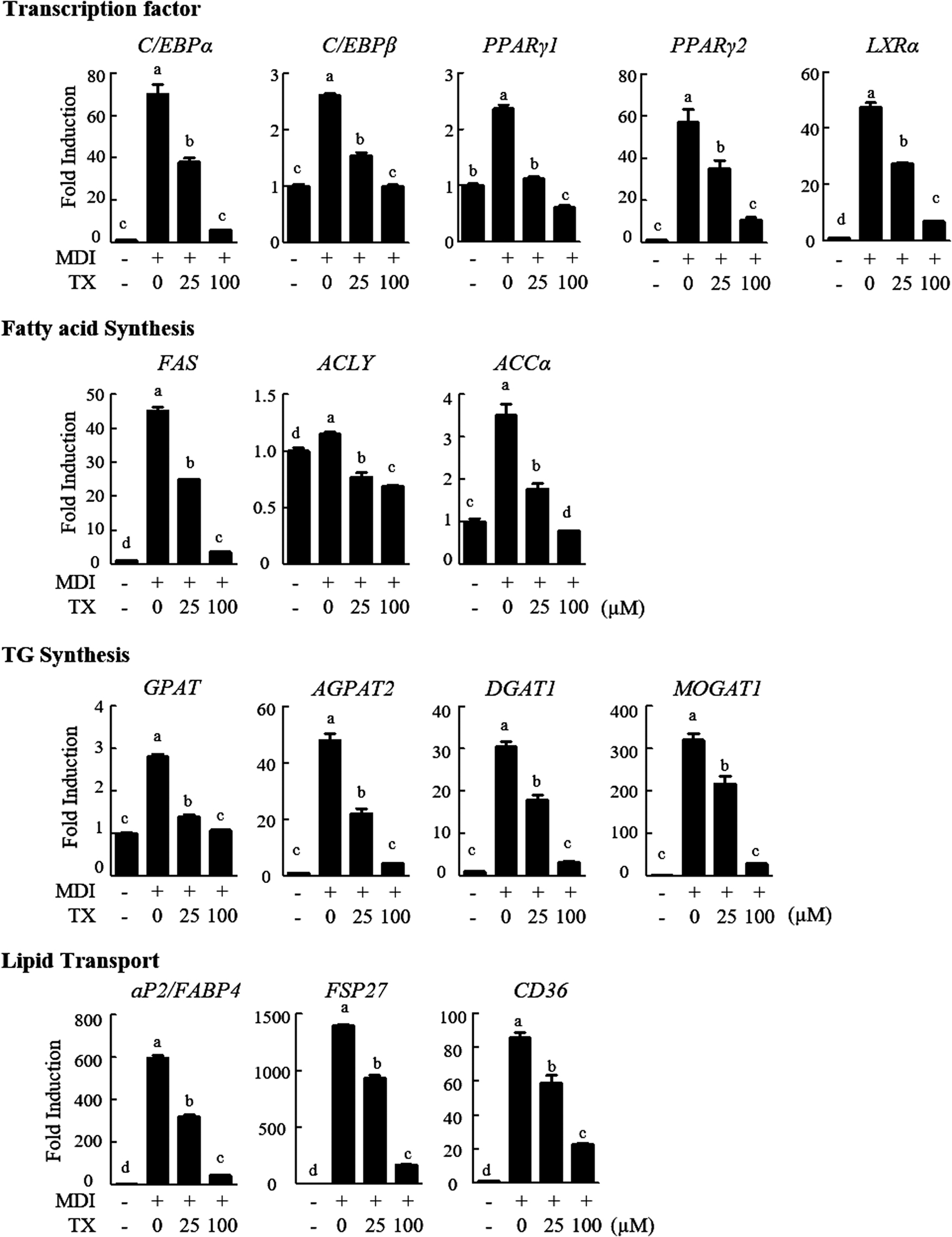
TX regulated mRNA expressions related to adipogenic genes, including transcription factors and related to fatty acid and TG synthesis and lipid transport. The 3T3-L1 cells were treated with TX during differentiation and its mRNA levels were measured by qRT-PCR. The values presented are the means ± SD of three independent experiments. Means with different superscript letters are significantly different, P < .05. qRT-PCR, quantitative reverse transcription polymerase chain reaction; TG, triacylglycerol.
TX suppressed adipogenesis through inhibition of pCAF in 3T3-L1 cells
We found that TX could regulate various downstream signaling pathways by inhibiting the expression of adipogenesis-related transcription factors. Various studies have shown that transcription factors and HATs are associated with adipogenesis. 20,27 Therefore, we investigated the HATs implicated in adipocyte differentiation.
Small interference RNA (siRNA) systems against representative HATs, p300, CBP, and pCAF, were developed (Fig. 3A). siRNA was first transiently transfected in 3T3-L1 preadipocytes, and then lipid accumulation and mRNA expression of the transcription factors were evaluated through Oil Red O staining and qRT-PCR, respectively. pCAF knockdown significantly inhibited adipogenesis and reduced the mRNA expression of transcription factors, PPARγ2, C/EBPα, and LXRα (Fig. 3B, C); however, it did not affect C/EBPβ expression. These results suggested that pCAF expression was specifically regulated during adipocyte differentiation.

pCAF knockdown inhibits lipid accumulation and adipogenic transcriptional factors.
TX inhibited acetylation of histone and nonhistone proteins through suppression of pCAF
To investigate whether TX affected pCAF activity, we measured the changes in HAT and pCAF activities following TX exposure using an in vitro cell-free system. TX treatment remarkably decreased the global HAT activity in a dose-dependent manner (Fig. 4A, left panel) and showed an inhibitory effect with an IC50 value of 21.63 μM (Fig. 4A, right panel). By changing the enzyme source from nuclear extract to pCAF recombinant protein bearing an HAT domain, the inhibitory effect of TX was confirmed. As expected, pCAF activity was significantly inhibited after TX treatment in a dose-dependent manner (Fig. 4B, left panel) with an IC50 value of 7.30 μM (Fig. 4, right panel).
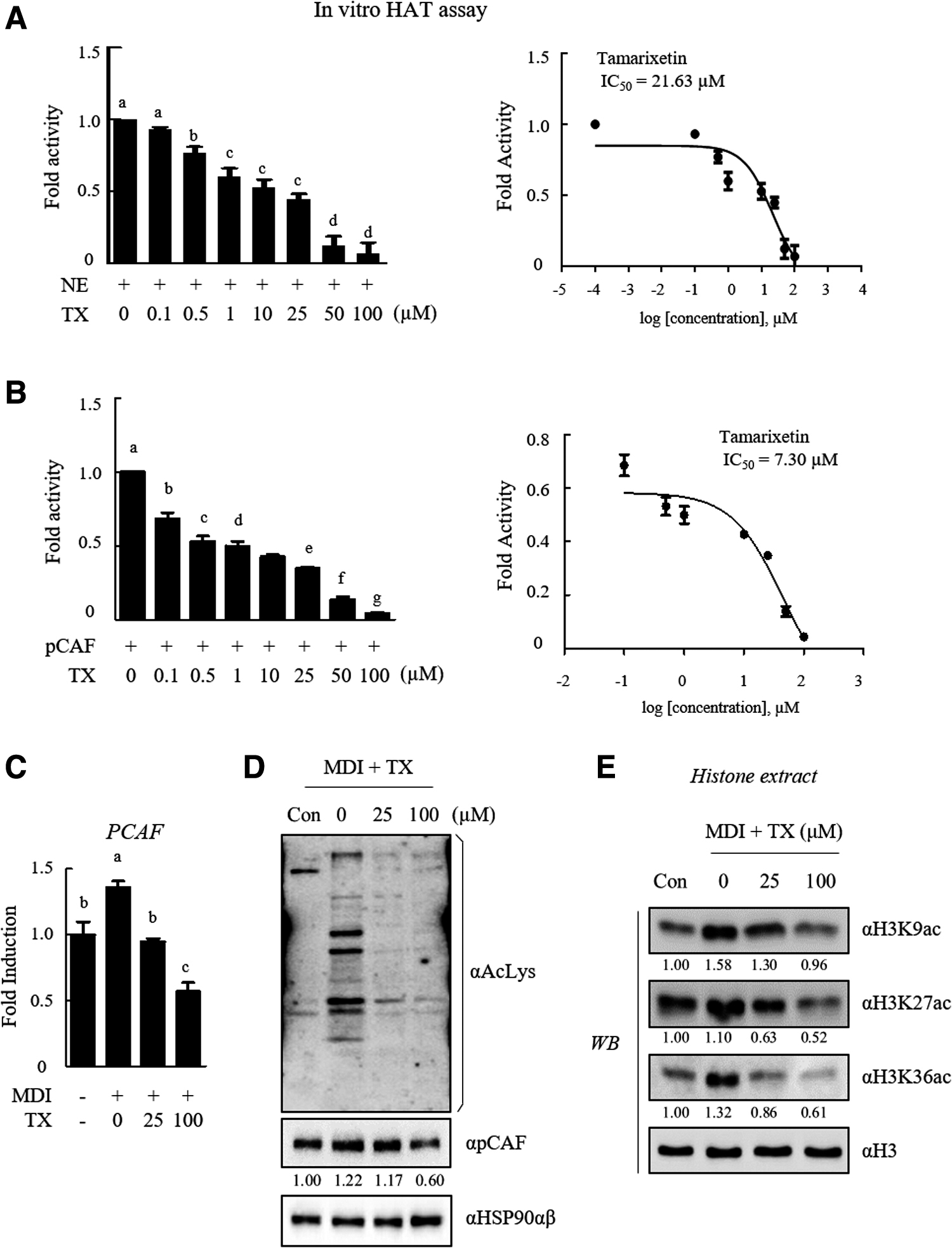
TX is a novel HAT inhibitor.
To determine whether the reduction of pCAF activity following TX treatment was associated with the inhibition of pCAF transcription by TX, qRT-PCR was performed after TX treatment during adipocyte differentiation. We found that TX treatment during adipogenesis effectively reduced the pCAF mRNA expression (Fig. 4C).
We further confirmed the effect of TX-mediated pCAF inhibition by determining the level of acetylation at the lysine residues of histone and nonhistone proteins in 3T3-L1 adipocytes. pCAF protein expression decreased following TX treatment; accordingly, the amount of acetylated protein decreased in the cell, and suppression of acetylation in H3K9, H3K27, and H3K36 residues was found (Fig. 4E). Collectively, these data suggested that TX represents a novel and potent pCAF inhibitor of histone and nonhistone proteins' acetylation.
TX decreased protein expression during 3T3-L1 cell differentiation
To verify whether TX effectively controls adipogenesis through pCAF inhibition, we monitored the changes in PPARγ, C/EBPα, and C/EBPβ expressions, as well as changes in pCAF expression, during adipogenesis with and without TX treatment. C/EBPβ expression increased during the beginning of adipocyte differentiation but gradually decreased from day 2 (Fig. 5A). In contrast, PPARγ and C/EBPα were expressed on days 1 and 2, respectively, and the expression decreased on day 6. Notably, the expression of pCAF gradually increased with adipocyte differentiation. TX treatment decreased the expression of pCAF, C/EBPα, C/EBPβ, and PPARγ (Fig. 5B). Altogether, these results demonstrated that TX downregulated the key transcription factors associated with adipogenesis by downregulating pCAF.
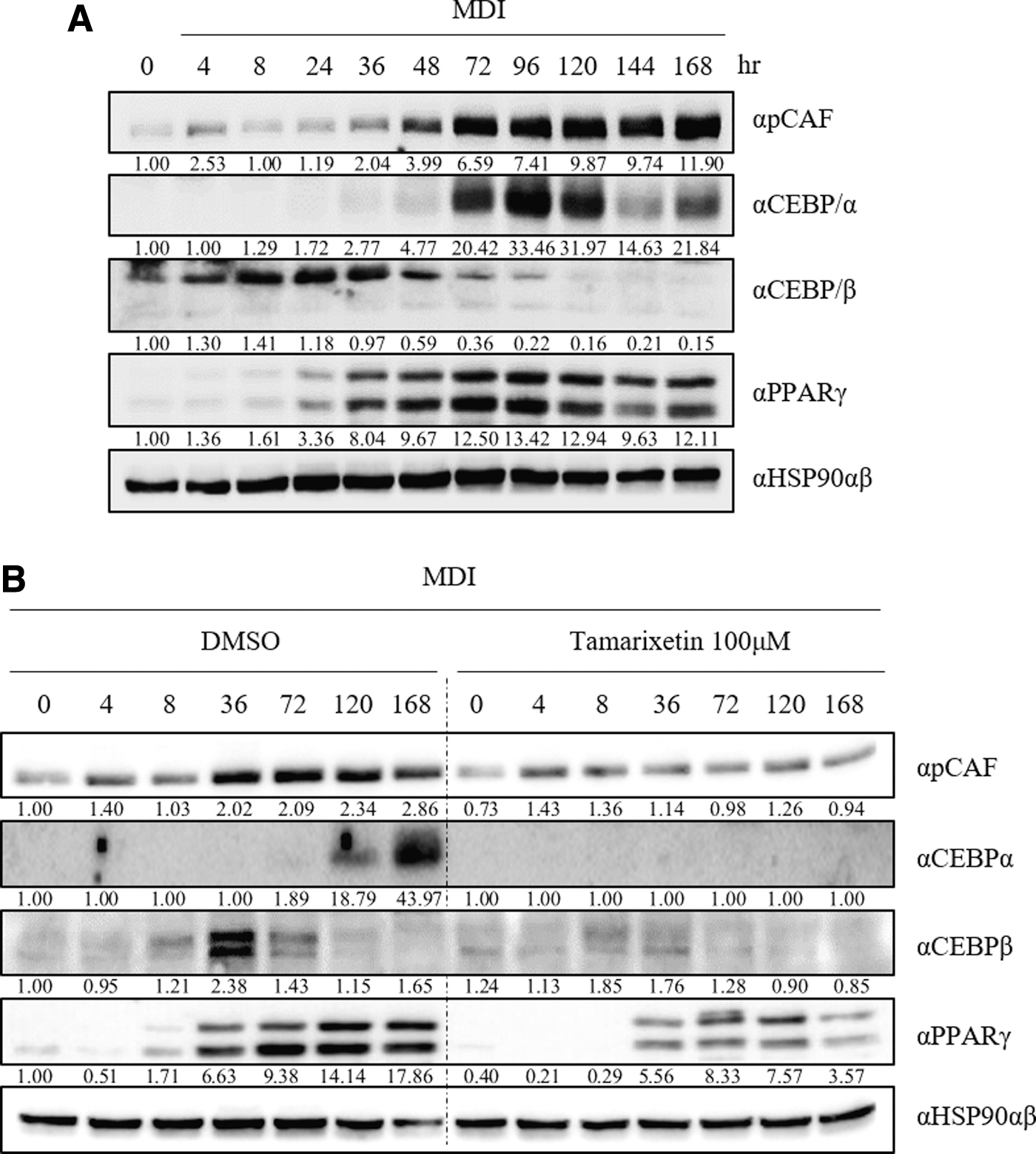
TX blocked 3T3-L1 adipocyte differentiation.
Discussion
Obesity is associated with the development and progression of various comorbidities, ranging from nonalcoholic fatty liver disease to hepatic cancer, cardiovascular disorders, and diabetes mellitus. 28 The etiology of obesity is attributed to the complex association between a wide variety of factors, including genetics, environment, hormones, and nutrition. In the present study, we investigated the antiobesity effects of TX, a food-derived metabolite, with a particular emphasis on the possibility of TX regulating certain epigenetic factors.
It has been reported that TX has anti-inflammatory, 25 gastroprotective, 29 cardioprotective, 30 and anticancer activity. 26 However, there is no direct evidence for the antiadipogenic effects of TX. The present study showed that lipid accumulation during adipogenesis was dramatically reduced without cytotoxicity following TX treatment (Fig. 1B). No previous study has reported the TX-mediated antiadipogenic effect; however, being a quercetin metabolite, TX was expected to have an antiadipogenic effect as several studies have reported the antiobesity effects of quercitin. 31
Notably, TX remarkably affected the expression of adipogenic genes (Fig. 2). However, qRT-PCR analysis demonstrated that TX did not selectively affect each gene. The observed effects were more likely to be caused by a ripple effect caused by upstream signaling pathways than from the direct regulation of each individual gene by TX; therefore, the TX-mediated downregulation of transcription factors may induce an overall decrease in the expression of downstream genes. Adipocyte differentiation is triggered from the initiation of the crosstalk of transcription factors.
C/EBPβ, which is expressed during the early stage of adipogenesis, is associated with PPARγ induction. 32 PPARγ is considered a master transcriptional factor 33 in adipogenesis facilitated by C/EBPα at later stages. 34,35 PPARγ and C/EBP factors function together to regulate the downstream genes associated with various signaling pathways for adipocyte differentiation. Consensus C/EBP-binding motifs have been identified in >90% of PPARγ occupancy regions of the promoter of its target genes. 36 Furthermore, this was confirmed from direct chromatin immunoprecipitation-chip analysis demonstrating the colocalization of C/EBPα at the majority of the PPARγ-binding region. 36 Based on these studies, TX might inhibit adipocyte differentiation through regulation at a level higher than that of transcriptional factors.
Acetylation of histone lysine residues, an epigenetic modification, is generally associated with gene activation. Gcn5 and PCAF have been shown to function upstream of PPARs during adipogenesis. 37 Furthermore, PCAF/Gcn5 acetylates C/EBPβ and sequentially activates C/EBPα expression through C/EBPβ. 38 Results of the present study showed that PCAF knockdown decreased adipocyte differentiation (Fig. 3). The formation of lipid droplets in the adipocytes and the mRNA expression of the key transcriptional factors, PPARγ and C/EBPα, were only affected by PCAF selectively, and not by p300 and CREB-binding protein (CBP). However, it has been reported that PPARγ gene expression is inhibited through ribozyme-mediated targeting by either p300 or CBP. 21
Furthermore, Crebbp knockout mice showed a remarkably reduced mass of white adipose tissue compared with that of wild-type mice. 39 Unlike PCAF, p300 and CBP regulate the expression of their target genes rather than that of transcriptional factors; therefore, we could not verify if lipid accumulation during adipogenesis was affected by both of the HAT enzymes. However, it is noteworthy that the expression of both p300 and CBP was low compared with that of PCAF in adipocytes (Fig. 3A). Based on these results, PCAF might be more abundant, at least during adipocyte differentiation.
TX dramatically decreased HAT activity in a cell-free system (Fig. 4), suggesting that TX may function as a global HAT inhibitor. Therefore, the application of TX may be desirable in treating diseases where the regulation of activity by either p300 or CBP is predominant. In practice, it has been reported that food-derived phytochemicals showing anti-HAT activity exert a global rather than a specific effect. 40,41
Analysis of adipogenesis showed that TX is a novel PCAF inhibitor. TX successfully suppressed the mRNA expression of PCAF resulting in a reduced level of protein acetylation in adipocytes. A previous study demonstrated the dynamic changes of acetylation status in human tissues, 42 indicating that protein acetylation is associated with a wide range of cellular functions. Furthermore, mass spectral analysis identified 189 acetylated proteins in cardiac tissues of obese mice. 43 Therefore, HAT enzymes represent important targets to prevent the development and progression of human diseases. Our study indicated that TX exhibited antiadipogenic effects through PCAF-mediated downregulation of key transcriptional factors (Fig. 5).
In conclusion, TX may disrupt mRNA expression of PCAF, thereby decreasing PCAF activity, which leads to decreased protein acetylation and mRNA expression of major transcription factors, such as PPARγ and C/EBPα. As a result of these events, the expression of various genes involved in adipogenesis is inhibited following TX treatment. Although TX was found to function as an HAT inhibitor and antiadipogenic agent in the present study, direct evidence regarding both the mode of action and application of TX in vivo remains to be elucidated. Given the limitations of this study, an in-depth study should be conducted.
Footnotes
Author Disclosure Statement
The authors declare no competing financial interests.
Funding Information
This work was supported by the Main Research Program (E0210400 and E0210601) of the Korea Food Research Institute (KFRI), funded by the Ministry of Science, ICT, and Future Planning.
Supplementary Material
Supplementary Table S1
Supplementary Table S2