
Abstract
Muscular atrophy is a muscle disease in which muscle mass and strength decrease due to aging, injury, metabolic disorders, or chronic conditions. Proteins in muscle tissue are degraded by the ubiquitin-proteasome pathway, and atrophy accelerates this pathway. Akkermansia muciniphila and Faecalibacterium prausnitzii strains are effective agents against metabolic and inflammatory diseases in next-generation probiotic research. In this study, we evaluated the efficacy of A. muciniphila strain EB-AMDK19 and F. prausnitzii strain EB-FPDK11 in a mouse model of muscular atrophy, since atrophy inhibits energy metabolism and immune activation. After oral administration of each strain for 4 weeks, the hind legs of the mice were fixed with a plaster cast to immobilize them for a week. As a result, the administration of EB-AMDK19 and EB-FPDK11 strains improved grip strength but did not increase muscle mass. At the molecular level, A. muciniphila and F. prausnitzii treatments decreased the expression levels of ubiquitin-proteasome genes, atrogin-1, MuRF, and cathepsin L. They increased the expression level of the mitochondrial biogenesis regulatory gene, PGC-1α. The effect of the strains was confirmed by a decrease in myostatin. Furthermore, A. muciniphila and F. prausnitzii modulated the immune function by enhancing ZO-1 and inhibiting IL-6. In particular, EB-AMDK19 promoted the expression of IL-10, an anti-inflammatory cytokine. These results suggest that A. muciniphila and F. prausnitzii may have beneficial effects on muscular atrophy, verified by newly isolated EB-AMDK19 and EB-FPDK11 as potential next-generation probiotics.
INTRODUCTION
Skeletal muscle is the most abundant tissue in the human body, and it plays a fundamental role in health and disease. 1 Muscular atrophy, which is characterized by decreased muscle mass, is caused by aging, muscle diseases, or chronic diseases. 2 –6 Decreased muscle mass and muscle strength are associated with mortality and have been reported recently to be associated with gut microbiota dysbiosis and systemic inflammation. 7,8
In the elderly with sarcopenia, the increase in TNF-α and IL-6 was also accompanied by a decrease in muscle mass and muscle strength. 9 Malnutrition accompanying inflammation reduces muscle cross-section area in early-stage inflammatory bowel disease (IBD). 10 Low lean mass and sarcopenia, common in IBD, are predictive indicators of osteopenia/osteoporosis. 11 In addition, cancer patients experience a type of sarcopenia called cachexia, and sarcopenic obesity is a sign of muscular atrophy accompanied by an increase in the amount of adipose tissue. 12,13 In the case of diabetes, poor glycemic control (HbA1c > 8.0%) was associated with lower skeletal muscle strength and quality. 14 Therefore, muscular atrophy is a symptom attributed to various diseases and can be a target for treatment or an indicator of the patient's condition.
As muscle atrophy develops, the bulk of myofibrillar proteins is degraded using the ubiquitin-proteasome pathway. 15 Atrogin-1 and muscle RING-finger protein1 (MuRF), known as ubiquitin protein ligases, are markedly upregulated during atrophy resulting from disuse-induced muscle wasting or cachexia. 16,17 In addition, cathepsin L, known as a proteolytic enzyme, is a direct target of forkhead box protein O1 (FOXO1), a key molecule in energy metabolism. 18 Myostatin, a transforming growth factor-β (TGF-β) family, is a regulator that inhibits muscle growth and activates the FOXO1-ubiquitin ligase pathway. 19 While the protein degradation pathway in muscle is accelerated by Atrogin-1, MuRF, CathepsinL, and myostatin, a loss of muscle mass occurs due to a low activity of the phosphoinositide 3-kinase-Akt- Mammalian target of rapamycin pathway, which is involved in protein synthesis. 20
In addition, the pathway of immune activation is related to muscular atrophy. Genetic inhibition of muscle nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) activity was shown to suppress intratracheal lipopolysaccharide (LPS)-induced MuRF and resulted in a significant sparing of muscle tissue. 21 In sarcopenia patients, the ratio of IL-6 (an inflammatory cytokine)/IL-10 (an anti-inflammatory cytokine) was increased. 22 As such, several mechanisms are simultaneously activated in muscular atrophy, and related symptoms appear as severe disease progresses, thus making it difficult to treat.
The essential role of gut microbiota in human health has been highly recognized. Thanks to advancements in human microbiome analysis, many bacterial species have been found to have a close relationship with human health and have been demonstrated to exert beneficial effects in animal models. For instance, it has been observed that a reduction in the prevalence of Akkermansia muciniphila and Faecalibacterium prausnitzii, which are highly abundant (accounting for 3–5% of the total microbiome) in healthy people, was related to the development of metabolic diseases (e.g., obesity, type 2 diabetes) and inflammatory diseases (e.g., IBD), respectively. 23 –27
Although there are no reports that A. muciniphila or F. prausnitzii is directly related to muscle disease, reduced abundance of A. muciniphila was observed in the hindlimb–unloading rodent model. 28 In addition, there was a significant depletion of F. prausnitzii and Roseburia inulinivorans, which produce short-chain fatty acids (SCFAs), in fecal microbiota of patients with sarcopenia. 29 Based on previous studies, for the first time, we investigated the effects of these two species (A. muciniphila and F. prausnitzii) on muscular atrophy by examining muscle strength, proteolysis, and immune-related signals in an animal model.
MATERIALS AND METHODS
Bacterial strains and growth condition
To isolate the A. muciniphila and F. prausnitzii, fresh human fecal samples were collected from each volunteer, and written informed consent was obtained for participants or their parents according to the legal guidelines. This research was approved by the Korea National Institute for Bioethics Policy (No. P01-201705-31-002) and the Institutional Review Board of Dongguk University, Ilsan Hospital (2018-06-001-012).
A. muciniphila EB-AMDK19 (KCTC13761BP) was isolated from a healthy Korean (female, age 35, body mass index [BMI] 23.3) according to the method previously described. 23,30 F. prausnitzii EB-FPDK11 was isolated from a healthy Korean (female, age 7, BMI 16.5) according to the previous methods with some modification. 31 Briefly, the extremely oxygen-sensitive colonies from each sample were obtained on a brain-heart infusion culture medium (herein referred to as YBHI) supplemented with 5 g/L yeast extract, 1 g/L cellobiose, 1 g/L maltose, and 0.5 g/L L-cysteine. A species-specific polymerase chain reaction (PCR) for A. muciniphila and F. prausnitzii was performed to identify species. Then, 16S rRNA gene sequencing was performed after PCR amplification.
The reference strains of A. muciniphila and F. prausnitzii used as a control in this study were obtained from the Deutsche Sammlung von Mikroorganismen und Zellkulturen German Collection of Microorganism and Cell Cultures (DSM22959, strain designation ATCC BAA-835, DSM17677, strain designation A2-165, respectively). They were cultured in soy-peptone based medium containing (l−1); 20 g soy-peptone; 10 g yeast extract; 2.5 g Dipotassium phosphate; 0.1 mg cyanocobalamin; 0.5 g L-cysteine hydrochloride, and some supplements under an anaerobic chamber filled with 90% nitrogen, 5% carbon dioxide, and 5% hydrogen at 37°C. For culturing A. muciniphila, the supplement was GlcNAc, lactose, fructose, and L-aspartic acid. For culturing F. prausnitzii, the supplement was GlcNAc, Glucose, and Sodium acetate.
For the animal experiment, bacterial cells were harvested by centrifugation at 12,000 g, 4°C for 5 min. Then, they were resuspended and diluted in anaerobic phosphate-buffered saline (PBS) with 20% glycerol to an end concentration of 1 × 108 CFUs/150 μL per mouse and stored at −80°C until oral administration.
Animals
Male C57BL/6 mice, aged 6 weeks, were purchased from Daehan Biolink Co., Ltd. (Cheongju, Republic of Korea). All mice were maintained in cages at 20 ± 3°C and humidity of 55 ± 5% with access to autoclaved distilled water and a normal chow diet under a 12-h light/12-h dark cycle. The study was approved by the “Institutional Animal Care and Use Committee” of Dongguk University (IACUC-2019-040-1) and was carried out in strict accordance with the recommendations of the “Guide for the Care and Use of Laboratory Animals” (Institute for Laboratory Animal Research, Committee for the Update of the Guide for the Care and Use of Laboratory Animals, National Research Council of The National Academies, USA; National Academies Press: Washington, DC, USA, 2011).
Treatments
The study scheme is depicted in Figure 1A. After an acclimatization period of 1 week, 63 mice were divided randomly into 7 groups. Nine mice were used in each group as: Normal, Immobilization,
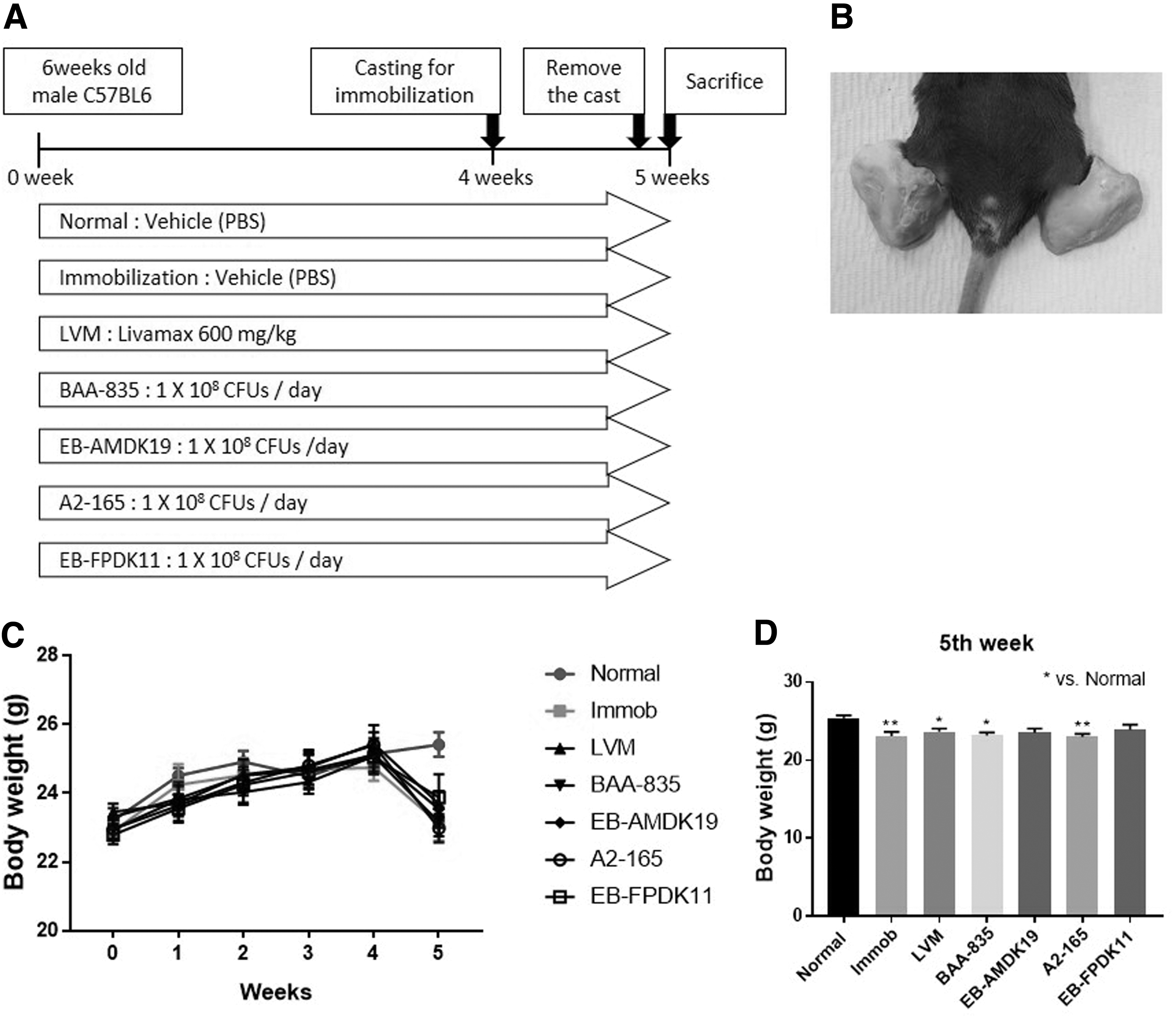
Immobilization-induced muscular atrophy in mouse.
Livamax, BAA-835, EB-AMDK19, A2-165, and EB-FPDK11. Livamax (LVM; Telcon RF Pharmaceutical, Republic of Korea) suspension was administered orally at 600 mg/kg as a branched-chain amino acid (BCAA) preparation. A. muciniphila ATCC BAA-835 (BAA-835 group), A. muciniphila EB-AMDK19 (EB-AMDK19 group), F. prausnitzii A2-165 (A2-165 group), or F. prausnitzii EB-FPDK11 (EB-FPDK11 group) was orally administered at a dose of 1 × 108 CFUs/150 μL of PBS/day. The normal and immobilization (Immob) groups were administered with sterile PBS as a vehicle. Oral administration was performed for 5 weeks. The animals were given free access to the diet and sterile water throughout the treatment period. All groups except the normal group had plaster casts on their hindlimbs for immobilization (Fig. 1B). The mice were placed in the plaster casts at the fourth week after oral administration and were maintained for 1 week.
Grip strength test
After removing the plaster cast at the end of 1 week, the grip strength of the hindlimbs was measured by referring to the methods of the manufacturer's protocol and other studies. 32 The grip strength of the hindlimbs was measured with Grip Strength Meter (JD-A-22; Jeungdo Bio & Plant Co., Ltd., Seoul, Republic of Korea). The front paws were raised and the tail gently pulled so that only the hind paws of the mouse could grasp the platform and measure the grip strength of the hindlimb. This measurement was repeated thrice.
Blood and tissue samples
At the end of experiment, blood was collected using cardiac puncture under anesthesia, and then the mice were sacrificed. The tibialis anterior and gastrocnemius muscle tissue and large intestine were harvested and weighed; then stored at −80°C for later analysis. Blood samples were centrifuged for 10 min at 1500 g and 4°C to obtain serum and then stored at −70°C.
Analyses of the serum
The myostatin concentration in serum was determined using an enzyme-linked immunosorbent assay (ELISA Kit; GDF-8/Myostatin Quantikine ELISA Kit, DGDF80; R&D Systems, Inc., Minneapolis, MN, USA) according to the manufacturer's instructions.
Tissue RNA extraction and quantitative PCR
Total RNA was isolated from muscle tissue and large intestine using a TRIzol® reagent (15596026; Invitrogen, Waltham, MA, USA) according to the manufacturer's protocol. For real-time PCR, cDNA was prepared using an M-MLV cDNA Synthesis Kit (EZ006M; Enzynomics Co., Ltd., Daejeon, Republic of Korea). Real-time quantitative PCR (qPCR) was carried out on a QuantStudio™ 3 (A28567; Thermo Fisher Scientific, Waltham, MA, USA) using a qPCR 2 × SYBR Green Premix (RT501UM; Enzynomics Co., Ltd.). The obtained data were normalized using glyceraldehyde-3-phosphatase dehydrogenase as a housekeeping gene. Primer sequences for the targeted mouse genes are listed in Table 1.
Primer Sequences for Quantitative Polymerase Chain Reaction
Analysis of EB-AMDK19 and EB-FPDK11 distribution in fecal samples from the mice
Before oral administration of EB-AMDK19 or EB-FPDK11, feces of mice were collected. EB-AMDK19 or EB-FPDK11 was orally administered at 1 × 108 CFU/150 μL each. After 4 h, feces were collected once more. For analysis of EB-AMDK19 and EB-FPDK11 abundance in feces, fecal bacterial DNA was extracted by QIAamp PowerFecal Pro DNA Kit. 33,34 After adjusting the DNA concentration, levels of EB-AMDK19 and EB-FPDK11 were assessed by qPCR with specific primers (Supplementary Table S1). PCR was run with SYBR® Green real-time PCR Master Mix on a QuantStudio 3 real-time PCR system (Applied Biosystems). The abundance of EB-AMDK19 and EB-FPDK11 was calculated as copies per gram of feces. Statistical analyses and plots were generated with GraphPad Prism version 7.04.
Statistical analyses
GraphPad Prism 5 (GraphPad, San Diego, CA, USA) was used for statistical analyses. For all experiments, one-way analysis of variance followed by Tukey's post hoc test was performed. All data are represented as the mean ± standard error of the mean. Significant differences are indicated as *P < .05, **P < .01, ***P < .001.
RESULTS
Improvement of grip strength by A. muciniphila or F. prausnitzii
Animal experiments were performed as shown in Figure 1A. Each group received orally administered treatments for 4 weeks before immobilization. Plaster casts were applied to both hindlimbs of the mice to induce muscular atrophy except in the Normal group (Fig. 1B). Plaster cast reduced the body weight by restricting the movement of the hindlimb, and it was confirmed that the body weights of the Immob, LVM, BAA-835, and A2-165 groups were significantly reduced compared to the normal group (P < .05 for LVM and BAA-835, P < .01 for Immob and A2-165; Fig. 1C, D).
After removing the plaster cast attached for a week, the muscle function was evaluated by measuring the grip strength of the hindlimb. The mouse was placed on a Grip Strength Meter, and grip strength of the hindlimbs was measured by pulling the tail (Fig. 2A). As a result of repeated measurements three times, the grip strength of the Immob, LVM, BAA-835, and A2-165 groups was significantly reduced compared to the normal group (P < .001 for all). In contrast, EB-AMDK19 and EB-FPDK11 showed significantly greater grip strength compared to Immob group, respectively (P < .05 and P < .01, respectively; Fig. 2B).

Grip strength test of hindlimbs.
Changes in muscle atrophy-related markers
To observe changes in hindlimb muscle, the weight of the muscle tissue was measured and the expression level of the relevant marker gene was investigated. As a result of measuring the weights of the tibialis anterior muscle and gastrocnemius muscle, the weights of all treatment groups except for the EB-AMDK19 group were significantly reduced in the tibialis anterior muscle (P < .01 for BAA-835, A2-165, and EB-FPDK11; P < .001 for Immob, LVM; Fig. 3A, B).

Changes in markers related to muscular atrophy due to A. muciniphila and F. prausnitzii in immobilization-induced muscle atrophy mice.
Atrogin-1 and MuRF are important enzymes for ubiquitin-mediated proteolysis in muscle and are used as markers of muscular atrophy. 35 As a result of comparing the expression level of Atrogin-1 in the tibialis anterior muscle, the Immob group was significantly increased compared to the normal group (P < .001), and all treatment groups were significantly decreased compared to the Immob group (P < .001 for all; Fig. 3C). MuRF expression level of the tibialis anterior muscle was significantly increased in the Immob group compared to the normal group (P < .05), but there was no significance in the treatment group (Fig. 3D). In the gastrocnemius, there were no significant changes in the atrogin-1 expression (Fig. 3E). The expression of MuRF in the gastrocnemius muscle of the A. muciniphila or F. prausnitzii oral administration groups was significantly decreased compared to the Immob group (P < .05 for BAA-835, EB-AMDK19, and A2-165; P < .01 for EB-FPDK11; Fig. 3F).
However, there was no significant difference between the normal group and the Immob group. Cathepsin L is a lysosomal endopeptidase enzyme that is involved in the initiation of protein degradation. This enzyme plays a significant role in the process of muscle fiber destruction, and its activity is increased in inflammatory myopathy. 36 Significant changes in cathepsin L were observed only in the tibialis anterior muscle (Fig. 3G). The expression level of CathepsinL was significantly increased in the Immob group compared to the normal group (P < .001), but decreased in the LVM, EB-AMDK19, and F. prausnitzii groups compared to the Immob group (P < .05 for EB-AMDK19 and F. prausnitzii; P < .01 for LVM). Immobilization did not induce significant changes in the expression of cathepsin L in the gastrocnemius among test groups (Fig. 3H).
In muscle, there is a protein synthesis process that is opposite to the protein degradation process, and this synthesis process is performed by the mitochondria. 37,38 PGC-1α is a marker of mitochondrial biogenesis and has been implicated in the regulation of skeletal muscle mass, particularly in the condition of muscle atrophy. 39,40 Elevated PGC-1α levels prevent the induction of autophagy and atrophy-specific ubiquitin ligase. 41 In the tibialis anterior muscle, the expression levels of PGC-1α were increased in all the treatment groups, but a significant induction was observed only in the BAA-835 and EB-AMDK19 groups compared with the Immob group (P < .01 and P < .001, respectively; Fig. 3I). In particular, the PGC-1α level of the EB-AMDK19 group was significantly higher compared with the LVM group. In gastrocnemius muscle, only the EB-AMDK19 and EB-FPDK11 groups showed significantly elevated levels of PGC-1α compared with the Immob group (P < .05, respectively; Fig. 3J).
Myostatin is a regulatory protein that restricts muscle development and growth, 42 and we measured the level of myostatin in serum for comparison (Fig. 3K). The concentration of myostatin was increased in the Immob group, but significantly decreased in the groups administered with LVM, the A. muciniphila strains, or F. prausnitzii strains (P < .05 for BAA-835; P < .01 for LVM; P < .001 for EB-AMDK19, A2-165, and EB-FPDK11).
Changes in the expression of markers related to gut barrier function and cytokines
Metabolites produced by gut microbiota are key actors in host-microbiota cross-talk. 43,44 These metabolites are activated by binding to G protein-coupled receptors (GPRs), and the activated GPRs convert extracellular signals into intracellular responses. 45,46 To observe whether each strain produces metabolites to activate GPR, the expression levels of GPR43 and GPR119 were observed. No significant change was observed in the expression of GPR43, but a significant increase by EB-AMDK19 was observed in the expression of GPR119 (P < .001) compared with those of the Immob group (Fig. 4A, B).
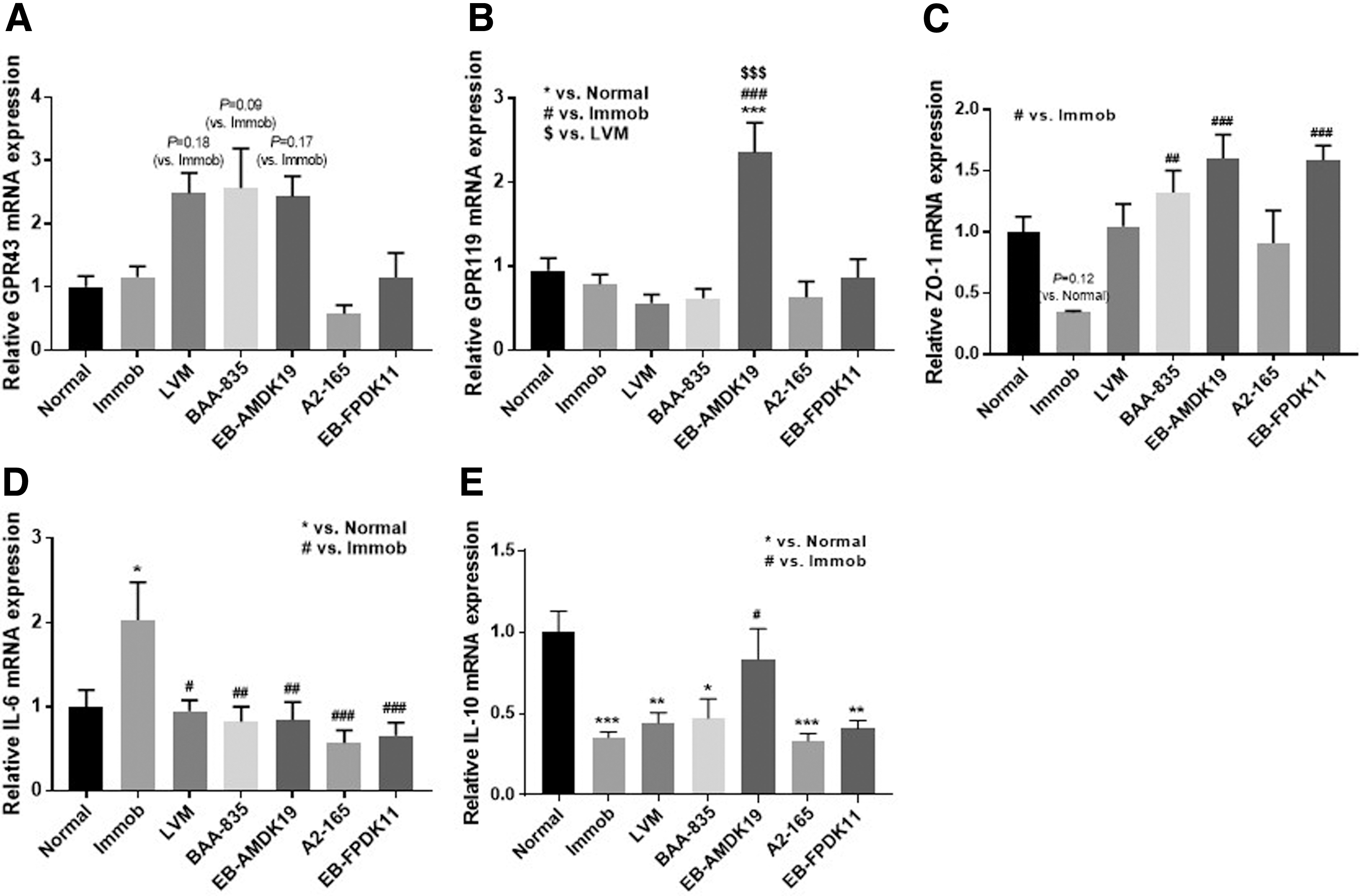
Changes in the expression of markers related to gut barrier function and cytokines by administration of A. muciniphila and F. prausnitzii.
Meanwhile, ZO-1 is responsible for the regulation of the intestinal-barrier function and intestinal permeability. 47 Immobilization resulted in decreased ZO-1 expression, but the expression level of ZO-1 was substantially increased in treatment groups, particularly in groups receiving A. muciniphila strains and EB-FPDK11 compared with that of the Immob group (P < .01 for BAA-835; P < .001 for EB-AMDK19 and EB-FPDK11; Fig. 4C).
In addition, we analyzed the expression levels of cytokines, IL-6 and IL-10, in the large intestine sample to investigate the immune response during immobilization. The expression of IL-6, a pro-inflammatory cytokine, was significantly increased in the Immob group (P < .05). However, the expression level of IL-6 in LVM, BAA-835, EB-AMDK19, A2-165, and EB-FPDK11 was significantly decreased (P < .05 for LVM; P < .01 for BAA-835 and EB-AMDK19; P < .001 for A2-165 and EB-FPDK11; Fig. 4D). The expression level of IL-10, known as an anti-inflammatory cytokine, was decreased in the Immob group, but significantly increased only in the EB-AMDK19 group (P < .05; Fig. 4E).
It was necessary to confirm whether the orally administered strain reached the intestine of the mouse. Feces before and after oral administration were collected and used for analysis. To exclude the endogenous A. muciniphila and F. prausnitzii levels, specific primers were designed for the administered EB-AMDK19 (Supplementary Fig. S1A) and EB-FPDK11 (Supplementary Fig. S1B). Levels of EB-AMDK19 and EB-FPDK11 in feces samples were assessed by qPCR with specific primers. Before administration of EB-AMDK19, the genome level of A. muciniphila in feces (5.07 ± 0.23 genome equivalents (log10)/g feces) was close to the limit of detection (5.05). This A. muciniphila level in feces was markedly increased after administration of EB-AMDK19 (Supplementary Fig. S2A). There were no differences between the level of endogenous F. prausnitzii in feces (3.81 ± 0.20 genome equivalents (log10)/g feces) and the limit of detection (3.72). As EB-FPDK11 was administered, the level of F. prausnitzii in feces was significantly increased (P < .05; Supplementary Fig. S2B).
DISCUSSION
A. muciniphila and F. prausnitzii are attracting attention as next-generation probiotics due to their efficacy for mitigating metabolic diseases such as obesity, diabetes, and nonalcoholic steatohepatitis. 25,48 The prevalence of metabolic disorders is also associated with decreased muscle activity, such as a lack of exercise or increased fat mass, which leads to decreased muscle mass. 49,50 In a study related to sarcopenic obesity, it was confirmed that the loss of muscle mass was related to physical functioning problems such as insulin resistance and inflammatory response. 51 EB-AMDK19 and EB-FPDK11 are strains isolated from healthy people and selected by comparing their efficacy. In the study of the efficacy of EB-AMDK19 in metabolic diseases, it was demonstrated that the efficacy differs depending on the subspecies even in the same strain. 30,52 Based on the beneficial effect of A. muciniphila and F. prausnitzii in previous metabolic disease studies, this study was conducted to compare and analyze the efficacy of type strains and selected strains in muscular atrophy.
In this study, a mouse model for muscular atrophy was established by plaster casting on the hindlimb. The plaster cast induced protein degradation by limiting muscle movement. This mouse model with immobilization provides a condition similar to muscular atrophy experienced by patients with limited movement due to surgery or patients with prolonged bed rest and represents the symptoms of muscle loss due to disease and aging. 53 –56 Grip strength, as a marker of physical function, was significantly greater in mice orally administered AMDK19 and EB-FPDK11, although muscle mass was not different from controls. This result was measured immediately after removing the plaster cast, and the recovery time may not have been sufficient.
Molecular analysis in muscle revealed partially significant results. Atrogin-1 and CathepsinL were significantly decreased only in the tibialis anterior muscle, and MuRF was significantly decreased only in the gastrocnemius muscle by the administration of EB-AMDK19 and EB-FPDK11. In contrast, PGC-1α was significantly increased by administration of EB-AMDK19 and EB-FPDK11 in both the tibialis anterior and gastrocnemius muscle. LVM, a BCAA agent used as a positive control, significantly decreased the protein levels of Atrogin-1 and MuRF in a previous study performed on a hindlimb suspension-induced muscle atrophy model, suggesting the possibility of protecting against proteolysis. 57
Compared with the type strain, the results of EB-AMDK19 and EB-FPDK11 are expected to regulate the amount of protein in the muscle by inhibiting Atrogin-1, MuRF, and CathepsinL, which are involved in the proteolytic pathway, and by promoting PGC-1α, which is involved in mitochondrial biosynthesis. The ubiquitination process by Atrogin-1, MuRF, and CathepsinL serves as a signal for protein turnover, which leads to rapid muscle mass loss. 58
Conversely, PGC-1α plays a critical role in maintaining muscle metabolic function. 59 Overexpression of PGC-1α, a transcriptional coactivator that induces mitochondrial biogenesis, dramatically prevents lovastatin (treatment for hyperlipidemia)-induced muscle damage and abrogated Atrogin-1. 60 Myostatin, a member of the TGF-β superfamily, is a cytokine that is produced and released by myocytes and that negatively regulates skeletal muscle in humans and animal models. 19 By the administration of A. muciniphila and F. prausnitzii, serum Myostatin levels were significantly decreased compared to the Immob group. Along with increased proteolysis induced by the ubiquitination process, skeletal muscle wasting manifested by myostatin occurs in the suppression of protein synthesis. 61
It was verified that EB-AMDK19 and EB-FPDK11 pass through the intestine with specific primers. Therefore, we tried to analyze how changes in the intestine affect muscles based on the gut-muscle axis. The link between gut microbiota and the production of gut peptides by enteroendocrine cells has been well documented. 62 Enteroendocrine cells act through GPRs and are present in high density in the ileum and colon, where the majority of bacteria reside. 63 GPRs are stimulated and activated by binding to metabolites produced by gut microbiota. GPR43, which is activated by SCFAs, promotes the secretion of GLP-1 and has various physiological actions such as promotion of insulin secretion or inhibition of glucagon production. Studies on GLP-1R agonists reported that it ameliorates muscle wasting by inhibiting myostatin. 64 As such, some studies suggest that GLP-1 based drugs will improve muscle mass and metabolism, but the exact mechanism is not known. 65,66
In metabolic diseases, the A. muciniphila strain promoted the secretion of GLP-1, 30 but no significant results were confirmed in this study on muscle diseases. Since there was no significant reduction of GPR43 in the immobilization model, it was assumed that the plaster casting did not cause any problematic damage to the enteroendocrine cells. Both A. muciniphila and F. prausnitzii are strains that produce SCFAs (GPR43 ligands), so it is necessary to confirm their expression level in an atrophy model established by other methods. GPR119 is stimulated and activated by oleic acid, a long-chain fatty acid (LCFA), and, like GPR43, promotes GLP-1 secretion. 67 The expression level of GPR119 was significantly increased only by EB-AMDK19. It has been reported that oral administration of A. muciniphila increases intestinal 2-oleoylglycerol levels. In addition, a number of phospholipid molecules, including oleoylethanolamide (OEA), act as GPR119 agonists, 68 and OEA supplementation increased the abundance of A. muciniphila bacteria. 69 EB-AMDK19 is considered to play an influential role in the regulation of metabolic diseases through GPRs.
Epithelial tight junction proteins, including ZO-1, are key players in regulating intestinal-barrier function and modulating intestinal permeability 47 to restrict free passage of toxic and infectious molecules from the gut lumen while allowing selective paracellular absorption across the tight junction. 70 The expression level of ZO-1 was decreased by immobilization but was increased by A. muciniphila and F. prausnitzii, particularly in groups receiving EB-AMDK19 and EB-FPDK11.
The integrity of the intestinal epithelial barrier maintained by a tight junction is crucial to protect against stress related to inflammation and infection. Factors related to the alteration of tight junction homeostasis include pro-inflammatory cytokines, pathogenic bacteria, LPS, and pathological conditions. 71 In this study, it was confirmed that the expression level of IL-6 increased and the expression level of IL-10 decreased as the immobilization model was induced. Inflammatory cytokines increase in skeletal muscle after sepsis and cachexia and contribute to maintaining pathological chronic inflammatory conditions. 72 As such, the inflammatory condition plays a vital role in muscle atrophy. 73 Stimulation of inflammatory cytokines induces the expressions of atrogin-1 and MuRF1 ubiquitin ligases and subsequent muscle protein degradation in skeletal muscle cells. 74 In addition, mRNA levels of inflammatory cytokines were increased in skeletal muscle after 14 days of cast immobilization. 75
Recently, several studies have hypothesized that the emergence of sarcopenia occurring with aging may be affected by the composition of gut microbiota. 8,29,76,77 Anabolic resistance (AR) associated with aging is a decrease in the rate of protein synthesis in muscle. The causal mechanism behind AR is related to the gut-muscle axis. 8
The composition of the fecal microbiota was changed in rat models of sarcopenia with age-related muscle mass wasting, and the presence of gut microbiota dysbiosis is associated with several metabolic alterations, involving protein synthesis, release of pro-anabolic mediators, inflammation, and insulin sensitivity. 78 In a study comparing the skeletal muscle from germ-free mice without gut microbiota to skeletal muscle from pathogen-free mice with gut microbiota, the mouse skeletal muscle in germ-free mouse showed atrophy, decreased expression of insulin-like growth factor 1, and reduced transcription of genes associated with skeletal muscle growth and mitochondrial function. In addition, transplanting the gut microbiota from pathogen-free mice into germ-free mice induced an increase in skeletal muscle mass and a reduction in muscle atrophy markers. 79
As research on the correlation between gut microbiota and muscle continues, it is necessary to select candidates for the gut microbiome that are important for improving muscle disease.
In conclusion, we found that EB-AMDK19 and EB-FPDK11 have some potential to prevent muscle atrophy. This suggests that specific strains of the species could be applied to treat metabolic disorders and complications such as muscular atrophy. However, the exact mechanism for intramuscular protein synthesis and catabolism could not be confirmed, and it was difficult to compare with other muscle diseases because it was performed in a model with injury to the muscle. For the study of sarcopenia, aging mice are required, and mice aged 12–20 months are commonly used as aging models. A future study needed to analyze the molecular signal in the muscle by long-term administration of the strains for at least 12 months with aging is required.
Footnotes
AUTHOR DISCLOSURE STATEMENT
No competing financial interests exist.
FUNDING INFORMATION
No funding was received for this article.
SUPPLEMENTARY MATERIAL
Supplementary Figure S1
Supplementary Figure S2
Supplementary Table S1