
Abstract
Inflammatory bowel diseases, including Crohn's disease and ulcerative colitis, are characterized by chronic gastrointestinal inflammation with continuous relapse-remission cycles. This study aimed to evaluate the protective effect of Bifidobacterium bifidum BGN4 as a probiotic or paraprobiotic against dextran sulfate sodium (DSS)-induced colitis in mice. Ten-week-old female BALB/c mice were randomly divided into five groups. The control (CON) and DSS groups received oral gavage of PBS, whereas the live B. bifidum (LIVE), heat-killed B. bifidum BGN4 (HEAT), and lysozyme-treated B. bifidum BGN4 (LYSOZYME) groups received live B. bifidum BGN4, heat-killed B. bifidum BGN4, and lysozyme-treated B. bifidum BGN4, respectively, for 10 days, followed by DSS supply to induce colitis. The paraprobiotic (HEAT and LYSOZYME) groups had less body weight loss and colon length shortening than the DSS or LIVE groups. The LYSOZYME group exhibited better preserved intestinal barrier integrity than the LIVE group by upregulating gap junction protein expression possibly through activating NOD-like receptor family pyrin domain containing 6/caspase-1/interleukin (IL)-18 signaling. The LYSOZYME group showed downregulated proinflammatory molecules, including p-inhibitor of kappa B proteins alpha (IκBα), cycloxygenase 2 (COX2), IL-1β, and T-bet, whereas the expression of the regulatory T cell transcription factor, forkhead box P3 expression, was increased. The paraprobiotic groups showed distinct separation of microbiota distribution and improved inflammation-associated dysbiosis. These results suggest that B. bifidum BGN4 paraprobiotics, especially lysozyme-treated BGN4, have a preventive effect against DSS-induced colitis, impacting intestinal barrier integrity, inflammation, and dysbiosis.
INTRODUCTION
Inflammatory bowel diseases (IBDs), including Crohn's disease and ulcerative colitis (UC), have become global diseases, possibly due to environmental changes causing disturbances of immune homeostasis. Dysbiosis is considered a crucial factor in the pathogenesis of IBDs because imbalanced microbiota causes failures of immune tolerance, which induce excessive and unnecessary inflammation. 1 Probiotics that possesses the ability to modulate gut microbiota composition have attracted attention as nontoxic alternative therapeutics for IBD management. 2
Interacting with intestinal gut flora and intestinal epithelial cells or immune cells, probiotics exert beneficial effects possibly by balancing the gut microbiota through competitive inhibition of pathogenic bacteria, production of antimicrobial molecules and short-chain fatty acids, improvement of intestinal barrier integrity, and immune modulations. 3,4 However, administration of probiotics to vulnerable populations, such as critically ill patients and neonates, may cause adverse events such as bacteremia. 5 –9 In addition, certain probiotics may transfer antibiotic resistance genes to the host microbial community. 9 Therefore, emerging concerns about the safety of probiotics enhance interest in the inactivated forms of probiotics, termed paraprobiotics.
Paraprobiotics are nonviable microbial cells, or crude cell extracts, which, when administered orally or topically in adequate amounts, confer a benefit to humans or animals. 10 The beneficial effects of paraprobiotics are suggested to be strain specific. 10 Also, paraprobiotic preparation methods influence the efficacy of paraprobiotics, as active substances are damaged or denatured during the inactivation procedure. 11
Heat treatment is the most commonly used inactivation method; however, a great extent of cell membrane damage, nutrient and ion losses, RNA strand breaks, ribosome aggregation, inactivation of essential enzymes, and protein coagulation has been reported. 11 Alternatively, other inactivation methods, including sonication, have been introduced and studies have suggested that intact bacterial cell wall substances or cellular proteins of nonviable probiotics exert beneficial biological activities. 11,12
Bifidobacterium bifidum BGN4, is a Gram-positive, nonmotile, anaerobic lactic acid bacteria that was isolated from the feces of breastfeeding infants. 13 Studies have shown that B. bifidum BGN4 probiotics exert outstanding cell adhesive capacity and immunomodulatory effects. 14 In addition, B. bifidum BGN4 ameliorates intestinal inflammation in a cluster of differentiation (CD)4+CD45high T cell transfer colitis model by suppressing immune cell infiltrate and reducing proinflammatory cytokines. 15
In this study, we compared the efficacy of B. bifidum BGN4 paraprobiotics to B. bifidum BGN4 probiotics in the prevention of murine colitis to extend the application of B. bifidum BGN4. Possible mechanisms of action in association with membrane integrity, T cell differentiation, and microbiome homeostasis were elucidated.
MATERIALS AND METHODS
Preparation of B. bifidum BGN4 probiotics and paraprobiotics
Bifidobacterium bifidum BGN4 originally isolated from a breastfed infant's fecal sample was provided by Bifido Co., Ltd. 13 Heat-killed BGN4 (HEAT) was prepared by autoclaving live B. bifidum BGN4 at 121°C for 15 min and lysozyme-treated BGN4 (LYSOZYME) was prepared by lysozyme treatment at 37°C for 2 h with shaking followed by sonication (20–25°C, 3 h). All samples were freeze-dried using a Speed-Vac (Ilshin Bio Base, Seoul, Korea). The morphologies of differently prepared paraprobiotics were observed using a field emission scanning electron microscope (FE-SEM, JSM-7500F; JEOL Ltd., Tokyo, Japan) at × 20,000 magnification under an accelerating voltage of 3 kV. Live B. bifidum BGN4 was applied as a control (CON) (Fig. 1).

Electoron microscopies showing different morphologies of paraprobiotics ( × 20,000).
Animals and experimental design
Female 10-week-old BALB/c mice were obtained from Charles River Laboratories (Shin-yokohama, Yokohama, Japan). After 1 week of adaptation, mice were randomized into five groups: CON group (n = 7), dextran sulfate sodium (DSS) group (n = 12), live B. bifidum (LIVE) group (n = 12), HEAT group (n = 12), and LYSOZYME group (n = 12). The CON and DSS groups received oral gavage of PBS (200 μL) for 10 days. For the same period, the LIVE, HEAT, and LYSOZYME groups received oral gavage of live B. bifidum BGN4 (109 colony-forming units (CFUs)/mouse/day), heat-killed B. bifidum BGN4, or lysozyme-treated B. bifidum BGN4 (LYSOZYME) in PBS (200 μL), respectively, followed by supplementation with DSS (36–50 kDa) dissolved in drinking water (2.5% w/w) for 5 days.
An equivalent number of CFUs given to the LIVE group mice was used to prepare HEAT and LYSOZYME sample supplied to the HEAT and LYSOZYME group, respectively. At the end of the experiment, mice were sacrificed, and blood samples were collected. Colon lengths were measured, and colon tissues were stored at −80°C until assayed. All procedures of this study were approved by the Institutional Animal Care and Use Committee of Sookmyung Women`s University (SMWU-IACUC-1806-010).
Disease activity index
The disease activity index (DAI) was used to determine the severity of disease symptoms. The DAI score (0–10) was assessed using three criteria: weight loss (no weight loss = 0, 1–5% loss = 1, 5–10% loss = 2, 10–20% loss = 3, and >20% loss = 4), stool consistency (formed = 0, mild soft = 1, very soft = 2, and watery stool = 3), and rectal bleeding (normal = 0, brown = 1, reddish = 2, and bloody = 3). Stool consistency and bleeding were monitored daily.
Histological analysis
The rectum tissue was fixed in 10% buffered formalin solution and embedded in paraffin. Paraffin blocks were cut into 5 μm sections and then stained with hematoxylin and eosin (H&E). To evaluate the histological score, a light microscope (Olympus PROVIS AX70, Tokyo, Japan), camera (Nikon DS-Ri2, Tokyo, Japan), and NIS-Elements BR Ver4 software (Nikon, Tokyo, Japan) were used. The criteria for histological scoring were inflammatory cell infiltrate, epithelial changes, and mucosal architecture, as previously described. 16 Paraffin blocks were also stained with an Alcian blue pH 2.5 solution using a NovaUltra special stain kit (IHC World, Ellicott City, MD, USA). Mucus layer integrity was determined based on Alcian blue intensity and the number of Alcian blue-positive cells (goblet cells).
Protein extraction and western blot analysis
The rectum tissue was homogenized using the PROPREP™ Protein Extraction Solution (iNtRON Biotechnology, Sungnam, Gyunggi, Korea). A total of 30–50 μg protein from each sample was separated using 4–20% SDS-PAGE and transferred to a polyvinylidene difluoride membrane (GE Healthcare, Chicago, IL, USA). Membranes were then blocked with 4% skim milk (BD Biosciences, Franklin Lakes, NJ, USA) and incubated with respective antibodies (Supplementary Data S1).
Measurement of serum CD14 and interleukin-10 levels
The serum levels of CD14 and interleukin (IL)-10 were determined using ELISA kits (MC140 [CD14] and M1000B [IL-10], R&D systems, Minneapolis, MN, USA), according to the manufacturer's instructions.
Gut microbiota analysis
Fecal bacterial DNA was extracted using a QIAamp DNA stool mini kit (Qiagen, Hilden, Rhine-Westphalia, Germany). PCR amplification was performed using primers targeting the V3-V4 regions of the 16S rRNA gene. For amplification, primers 341F (5′-TCGTCGGCAGCGTCAGATGTGTATAAGAGACAG_CCTACGGGNGGCWGCAG-3′; the underlining sequence indicates the target region primer) and 805R (5′-GTCTCGTGGGCTCGGAGATGTGTATAAGAGACAGGACTACHVGGGTATCTAATCC-3′) were used. Secondary amplification to attach the Illumina NexTera barcode was performed using the i5 forward primer (5′AATGATACGGCGACCACCGAGATCTACACXXXXXXXXTCGTCGGCA GCG TC-3′; X indicates the barcode region) and i7 reverse primer (5′-CAAGCAGAAGAC GGCATACGAGAT-XXXXXXXX-GTCTCGTGGGCTCGG-3′).
Mixed amplicons were pooled and sequenced using the Illumina MiSeq Sequencing system (Illumina, San Diego, CA, USA). Reads were normalized to 228,832, and α-diversity indices (Chao and abundance-based coverage estimator [ACE]) were estimated using an in-house code.
The Chao index is a nonparametric method for estimating the number of species in a community developed by Ann Chao. ACE is also a nonparametric method for estimating the number of species using sample coverage, which is defined as the sum of probabilities of observed species. Principal coordinate analysis (PCoA) was used based on the Bray-Curtis distance at the genus level. Linear discriminant analysis effect size was used with the Kruskal–Wallis rank-sum test based on a P-value/P-value (false discovery rate) <0.05 and a linear discriminant analysis (LDA) score >2.0, to identify significantly different taxa between groups. 17 All data were analyzed using the EzBioCloud database (ChunLab, Inc., Seoul, Korea). 18
Statistical analysis
Statistical analysis was performed using the SAS software (version 9.4; SAS Institute, Inc., Cary, NC, USA). Experimental data are expressed as mean ± standard error of means. One-way analysis of variance, followed by Duncan's multiple range test, was used to analyze the difference between five experimental groups. Student's t-test was used to analyze the significance between two groups when there is a tendency of difference without a statistical significance by Duncan's multiple range test. Differences were considered statistically significant at P < .05.
RESULTS
Bifidobacterium bifidum BGN4 paraprobiotics attenuated the symptoms of DSS-induced colitis
To confirm the preventive effect of B. bifidum BGN4 against colitis, body weight loss, colon length shortening, and DAI score were evaluated. Heat-treated paraprobiotic supplementation significantly suppressed body weight loss and reduced colon shortening (Fig. 2A, B). The LYSOZYME groups showed lower DAI scores than the DSS group (Fig. 2C), and the difference was statistically significant.

Effects B. bifidum BGN4 probiotics or paraprobiotics on DSS-induced colitis.
Bifidobacterium bifidum BGN4 paraprobiotics maintained intestinal barrier integrity against the DSS challenge
The LYSOZYME group had significantly lower inflammatory cell infiltration concentration and mucosal architecture damage score compared to the DSS group (Table 1, Fig. 3A). The LYSOZYME group exhibited a significantly higher expression of the junction proteins, occludin and E-cadherin (Fig. 3B). No statistical improvement was observed in the expression of claudin 1. To confirm intestinal barrier integrity, serum CD14 concentrations, representing bacteria-originated endotoxin (LPS) infiltration into the blood, were measured. LIVE, HEAT, and LYSOZYE groups showed significantly lower LPS infiltration than animals in the CON group (Fig. 3C).
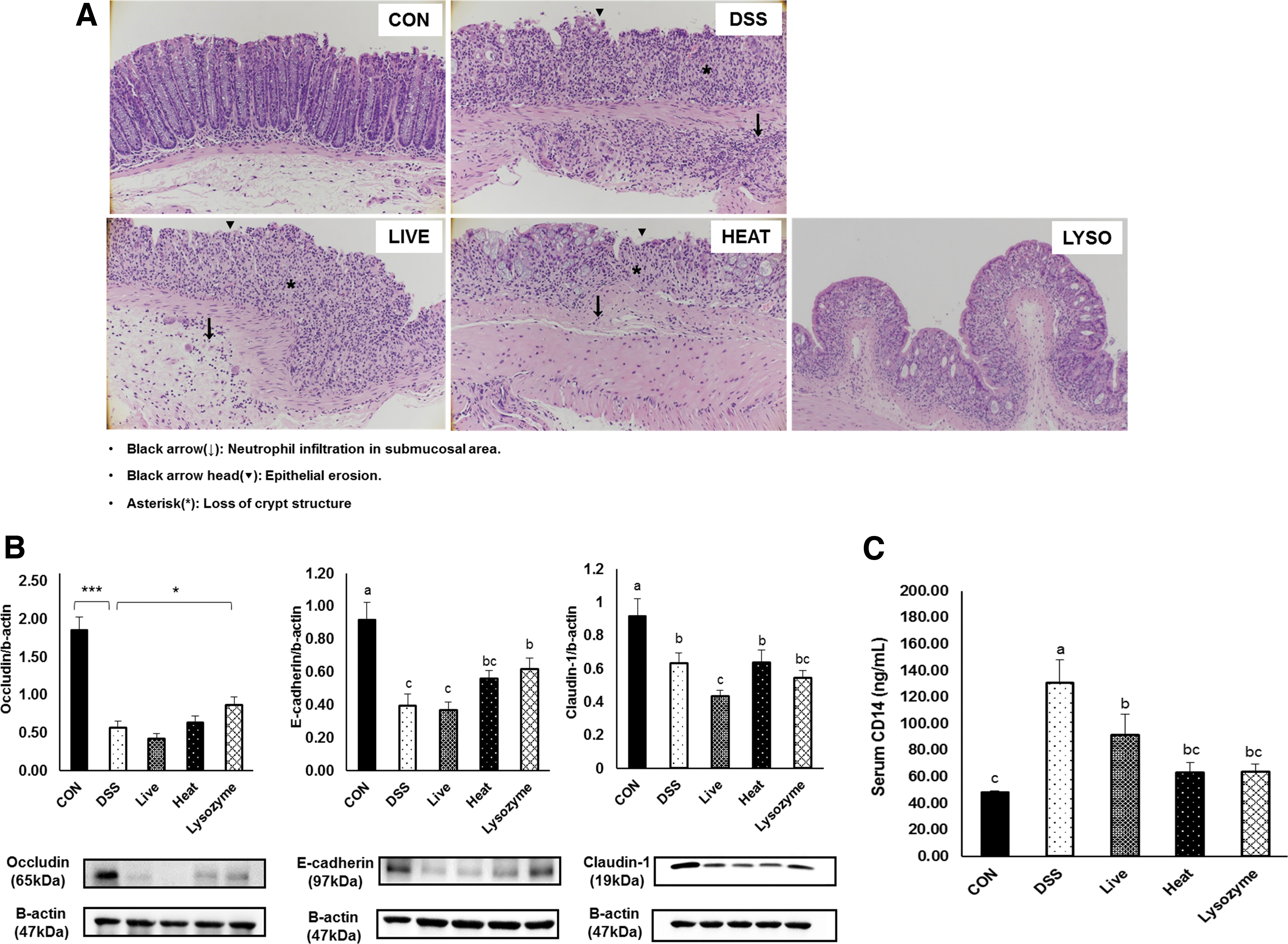
Effects of B. bifidum BGN4 probiotics or paraprobiotics on intestinal barrier integrity and function against DSS challenge
Histological Scores
All data represented means ± SEMs.; n = 7–12 mice per group; a–cMeans with different superscripts are significantly different based on the Duncan's multiple range tests (P < .05).
CON, control; DSS, dextran sulfate sodium; HEAT, heat-killed B. bifidum BGN4; LIVE, live B. bifidum; LYSOZYME, lysozyme-treated B. bifidum BGN4; SEM, standard error of mean.
Bifidobacterium bifidum BGN4 paraprobiotics strengthened the mucus layers through NOD-like receptor family pyrin domain containing 6/caspase-1/IL-18 signaling
Mucus layers were observed by Alcian blue staining (Fig. 4A). The number of goblet cells and the mucus layer area were significantly decreased in the DSS group. The LYSOZYME group showed a significantly larger mucus area than the DSS group, and goblet cell counts were higher in both the HEAT and LYSOZYME groups than the count in the DSS group (Fig. 4B). The DSS group showed significantly lower NOD-like receptor family pyrin domain containing 6 (NLRP6) expression than the CON group. However, the HEAT and LYSOZYME groups exhibited significantly higher NLRP6 expression than the DSS and LIVE groups (Fig. 4C).
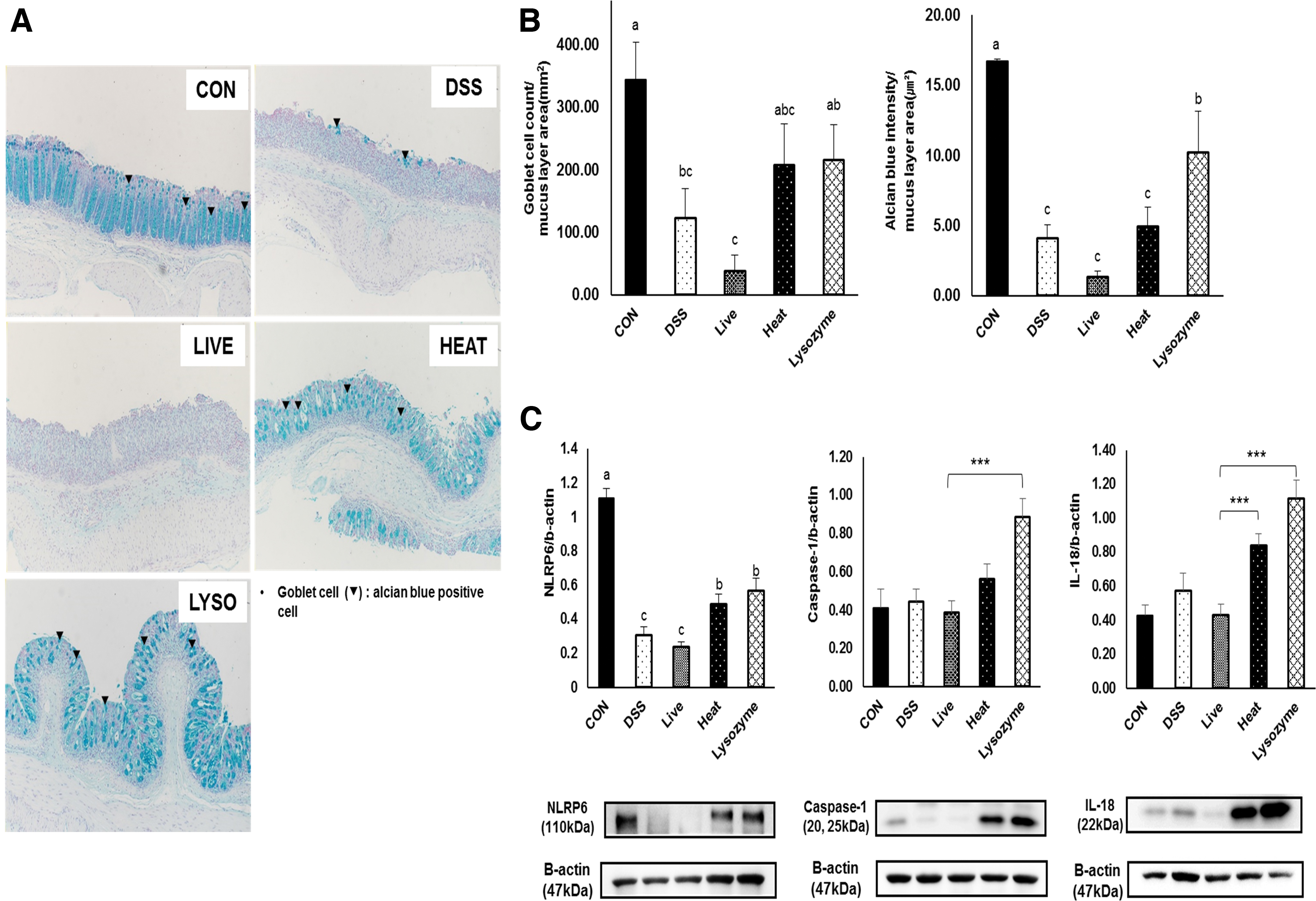
Effects of B. bifidum BGN4 probiotics or paraprobiotics on the intestinal mucosal barrier and NLRP6, caspase-1, and IL-18.
Caspase-1 and IL-18 expression were not affected by DSS administration; interestingly however, paraprobiotic supplementation increased the expression of caspase-1 and IL-18 compared to those of other groups (Fig. 4C).
Bifidobacterium bifidum BGN4 paraprobiotics suppressed the expression of proinflammatory signaling molecules
DSS administration significantly increased the protein expression nuclear factor kappa B (NF-κB) activation signaling-associated molecules, which include p-mitogen associated protein kinase (MAPK) and p-inhibitor of kappa B proteins alpha (IκBα), IL-1β, and cycloxygenase 2 (COX2), possibly causing tissue inflammation. However, the LYSOZYME group had significantly lower expression of p-IκBα, COX2, and IL-1β than the DSS group. Likewise, the HEAT group exhibited significantly (P < .05) lower expression of p-IκBα than the DSS group (Fig. 5A).

Effects of B. bifidum BGN4 probiotics or paraprobiotics on intestinal innate and adaptive immune responses
To investigate the effects of probiotic and paraprobiotic supplementation on T cell differentiation, the expression of signature transcription factors, including T-bet for Th1 cells, GATA binding protein 3 (GATA3) for Th2 cells, and forkhead box P3 (FOXP3) for regulatory T cells (Tregs), was measured. The expression of T-bet was significantly increased in the DSS group and was lowered by paraprobiotic supplementation.
GATA3 expression was significantly increased in the DSS group; however, paraprobiotic supplementation did not exert any change in the expression level. FOXP3 expression was significantly decreased in the DSS group, while probiotic or paraprobiotic supplementation increased its expression, although only the HEAT group showed a statistical significance (Fig. 5B). Figure 5C shows that serum IL-10 concentrations were increased in the DSS group and paraprobiotic supplementation reduced the circulating levels of IL-10.
Bifidobacterium bifidum BGN4 paraprobiotics suppressed DSS-induced dysbiosis
DSS supplementation significantly reduced the gut microbial diversity (Table 2) and altered the microbiota composition (Fig. 6A). DSS-induced colitis significantly altered Bacteroidetes and Verrucomicrobia (Table 3). Beta-diversity analyses using PCoA demonstrated significant segregation of the CON group from the four other DSS-treated groups. Moreover, the probiotic and paraprobiotic groups were separated into distinct groups (Fig. 6B). Many DSS-induced alterations in the proportions of taxa were restored by probiotic or paraprobiotic supplementation (Fig. 6C), and LEfSe analysis indicated that each group showed distinct microbial enrichment (Fig. 6D).
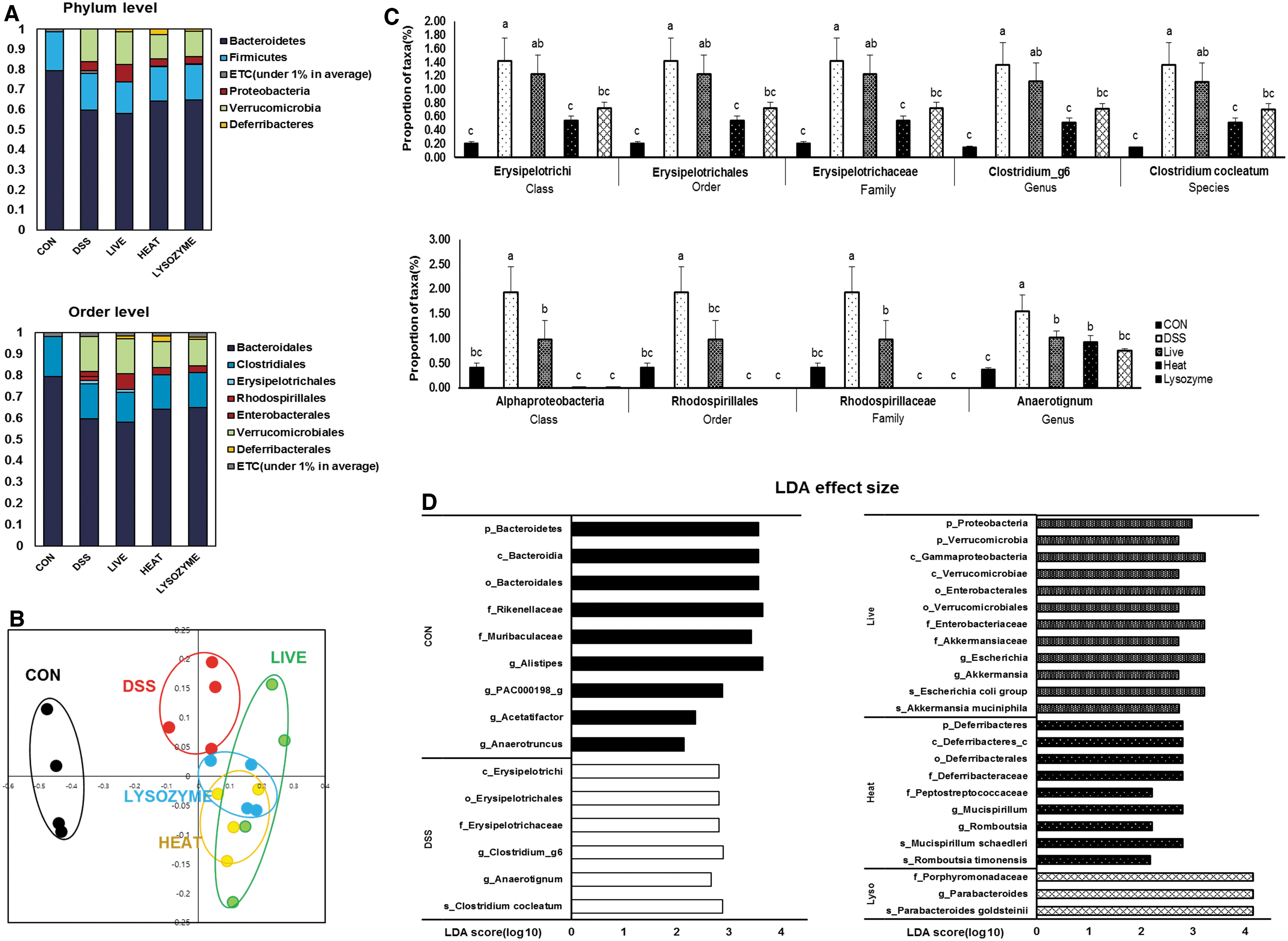
Effects of B. bifidum BGN4 probiotics or paraprobiotics on DSS-induced dysbiosis.
α-Diversity of Gut Microbiota
All data represented means ± SEMs; n = 7–12 mice per group; a–cMeans with different superscripts are significantly different based on the Duncan's multiple range tests (P < .05).
ACE, abundance-based coverage estimator.
Abundances of Certain Taxa at the Phylum Level
All data represented mean ± SEM; n = 7–12 mice per group; a–cMeans with different superscripts are significantly different based on the Duncan's multiple range tests (P < .05).
DISCUSSION
In this study, B. bifidum BGN4 paraprobiotics prevented DSS-induced colon shortening, weight loss, and disease activities. Especially, LYSOZYME effectively maintained E-cadherin and occludin expressions, and mucus layers. Enhanced epithelial barrier function was also evidenced by lower CD14 levels, representing infiltration concentrations of luminal content, especially LPS, in the B. bifidum BGN4 paraprobiotic groups. In addition, B. bifidum BGN4 paraprobiotics suppressed proinflammatory molecules involved in NF-κB activation, which may be linked to the transcriptional regulation of T cell development (Fig. 7).

Supply of paraprobiotics alleviates chemically induced intestinal inflammation. The LYSOZYME preserves intestinal barrier integrity by maintaining mucus layers and by activating NLRP6/caspase-1/IL-18 signaling. It also suppressed inflammatory responses through downregulated p-IκBα, COX-2, IL-1β, and T-bet expression and upregulated FOXP3 expression. The paraprobiotic supplementation exhibits distinct separation of microbiota distribution and lower inflammation-induced dysbiosis. Color images are available online.
IBD is frequently associated with a thinner colonic mucosal layer and increased gut permeability. 19 A recent study showed that inflammatory conditions decrease the expression of the connexin 43-associated junction complex partners, E-cadherin, zonula occludens-1, and β-catenin. 20,21 Although studies on claudin-1 have shown conflicting results, 22 reduced claudin-1 is reported in a DSS-induced colitis model in mice, whereas the onset of colitis and colorectal cancer (CRC) is shown to be associated with increased claudin-1 expression through the inhibition of goblet cell differentiation and interference with the formation of a normal mucus layer. 23
A number of studies have provided evidence that different strains of Bifidobacterium exhibit efficacies in intestinal barrier protection. B. longum, 24 B. infantis, 25 B. bifidum, 26 B. longum LC67, 27 B. adolescentis IM38, 28 B. longum CCM7952, 29 B. bifidum ATCC29521, 30 and B. longum YS108R 31 alleviate intestinal inflammation by tightening the intestinal barrier. However, mechanistic explanations of their efficacies are subtle and studies on their nonviable preparations are beginning to be explored.
Paraprobiotics consisting of bacterial remnants, including lipoteichoic acid (LTA), peptidoglycan, DNA, and muramyl dipeptide, are suggested to be involved in barrier-strengthening signals. 32 NLRP6 is a member of the NOD-like receptor family that recognizes microbial stimuli, such as LTA or other metabolites, including taurine and histamine, 32 and is commonly expressed in the colon, especially in goblet cells.
As an innate immune receptor, NLRP6 manages mucus homeostasis by regulating mucin 2 exocytosis and the production of IL-1β and IL-18. 33 In the DSS-induced colitis model, IL-18 injection before the DSS treatment prevents colitis by improving goblet cell function and intestinal barrier integrity, suggesting NLRP-6 mediation of IL-18 production is an important preventive response to protect epithelial integrity and to maintain homeostasis of immunological responses. 31 Another study indicated NLRP6 deficiency results in reduced IL-18 levels and altered microbial composition that cause exacerbation of chemically induced colitis. 34
As expected, results from this study showed B. bifidum BGN4 paraprobiotic-supplemented groups exhibited significantly higher NLRP6 expression than the DSS and LIVE group. Paraprobiotics may have an advantage over probiotics in terms of epithelial penetration to exert positive effects due to smaller sizes of active components. 23 The relatively lower anti-inflammatory activities of the HEAT group compared to those of the LYSOZYME group suggest that biologically active components in paraprobiotics might be vulnerable to heat treatment to a certain extent.
T cell subset imbalance aggravates inflammatory responses in IBD. 35 Polarization of proinflammatory T cells, such as Th1 and Th2 cells, and their decreased differentiation into anti-inflammatory Tregs, reducing the Treg subset population, have been observed in DSS-induced colitis in mice. 36 In this study, the expression of transcription factors specific to each T-helper cell was determined. B. bifidum BGN4 paraprobiotics significantly decreased T-bet expression, while increasing FOXP3 expression, suggesting a decreased ratio of Th1 to Treg cells. B. bifidum BGN4 probiotics showed a similar trend.
The current results are consistent with previous results obtained using B. bifidum BGN4 probiotics in a T cell transfer colitis model in which BGN4 reduced immune cell infiltrates and downregulated Th1-type cytokines. 15 NF-κB-mediated inflammation might be alleviated by B. bifidum BGN4 paraprobiotics, as well as probiotics, as demonstrated by changes in IκBα, MAPK, COX2, and IL-1β expression.
These results suggest that B. bifidum BGN4 suppressed DSS-induced inflammatory responses, possibly by polarizing T-helper cell differentiation into Tregs; again, paraprobiotics may be effective in driving direct immunomodulatory effects. The lack of significant T cell subset regulatory effects of paraprobioitcs might be due to a short duration of DSS supply in this acute colitis model; although studies have indicated that both acute and chronic colitis models exhibit significantly higher percentages of IFNγ+ and IL-17 cells, the percentage of FoxP3+ cells was decreased. 37
More recent studies have revealed that DSS treatment dramatically alters the gut microbiota composition and triggers dysbiosis. 38 In this study, DSS induced a decreased α-diversity in gut flora and altered the microbial composition. PCoA showed a clear separation between the cluster of probiotic- or paraprobiotic-treated and DSS-treated mice, indicating that probiotics and paraprobiotics transformed the microbial composition.
Microbial composition analysis revealed that B. bifidum BGN4 paraprobiotics restored DSS-induced increases in Erysipelotrichia (class), Erysipelotrichales (order), Erysipelotrichaceae (family), Clostridium_g6 (genus), and Clostridium cocleatum (species). Erysipelotrichaceae is known to be involved in the development of IBD and CRC. 39 B. bifidum BGN4 paraprobiotics and probiotics lowered the proportions of Alphaproteobacteria (class), Rhodospirillales (order), Rhodospirillaceae (family), and Anaerotignum (genus), which are increased in patients with severe UC and in DSS-induced colitis mice. 40
The LDA score also suggested that paraprobiotics partially suppressed dysbiosis. The CON group was abundant in commensal taxa, such as Muribaculaceae, Rikenellaceae, and Alistipes, which belong to Bacteroidetes, and Acetatifactor and Anaerotruncus, which belong to the butyrate-producing bacterial taxa Lachnospiraceae and Ruminococcaceae, respectively. 41 The DSS group possessed inflammation-relevant bacteria in abundance; Erysipelotrichaceae has been reported to be increased in CRC patients, and Clostridium cocleatum increases were reported in colitic mice. 42,43 The LIVE group had abundant mucin-degrading Akkermansia muciniphila and Enterobacteriaceae, which overgrow in the inflammatory state. A previous study showed that IBD patients exhibit a higher proportion of Enterobacteriaceae than the CON group. 44
In addition, Escherichia coli increased in IL-10 knockout mice and chemically induced colitis mice. 45 In this study, the HEAT group had a higher proportion of the commensal bacteria Mucispirillum and Romboutsia. Decreases in both of these commensal bacteria have been reported in colon polyps and cancerous tissues. 46 Finally, the LYSOZYME group had an abundance of Parabacteroides goldsteinii, which has shown antiobesity and anti-inflammatory effects and increases in insulin sensitivity in an obese mouse model. 47 Taken together, B. bifidum BGN4 paraprobiotics may partially suppress dysbiosis following DSS challenge.
In this study, B. bifidum BGN4 probiotics showed relatively minor effects in alleviating inflammation compared to paraprobiotics. Biologically active bacterial remnants produced during the inactivation process may have acted as signaling molecules to exert superior anti-inflammatory activities in this study. Different microbial compositions between probiotic- or paraprobiotic-supplemented animals are another possible explanation for differential efficacies. Interestingly, a previous colitis study using B. bifidum BGN4 probiotics was effective against colitis by suppressing immune cell infiltration and reducing the production of proinflammatory cytokines. 19
In that study, a CD4+CD45RBhigh T cell transfer colitis model, which is suitable for investigating the adaptive immune system, was used; B. bifidum BGN4 exerted beneficial effects through interaction with IECs. The DSS-induced colitis model used in this study was characterized by profound damage to IECs, which might explain the lack of efficacy of probiotics in animals. Although further studies are required to identify the functional components of these paraprobiotics, B. bifidum BGN4 paraprobiotics could be potential agents for preventing colitis and be developed as safer and more effective products than probiotics.
Footnotes
AUTHORS' CONTRIBUTIONS
G.-E.J., M.-S.P., J.-H.P., B.-H.L., and M.-K.S. developed the idea. G.-E.J. and M.-S.P. provided their probiotic strain (B. bifidum BGN4). J.-H.P. and B.-H.L. produced B. bifidum BGN4 paraprobiotics. S.-Y.L. designed detailed experiments models and carried out animal experiments. S.-Y.L. analyzed experiments under the supervision of M.-K.S. M.-K.S. and S.-Y.L. wrote the article. All authors have read and agreed to the published version of the article.
AUTHOR DISCLOSURE STATEMENT
S.-Y.L., B.-H.L., J.-H.P., and M.-K.S. declare no conflicts of interest. G.E.J. and M.S.P. are directly employed by BIFIDO Co., Ltd. G.E.J. and M.S.P. hold BIFIDO Co., Ltd. stocks as a CEO and CTO, respectively.
FUNDING INFORMATION
This research was supported by the Korea Institute of Planning and Evaluation for Technology in Food, Agriculture and Forestry (IPET) through the High Value-added Food Technology Development Program, funded by the Ministry of Agriculture, Food and Rural Affairs (MAFRA-RRB—[317043-03-2-HD 030]) and the Medical Research Center Program through National Research Foundation (NRF) grant funded by the Korean government (MSIP) (no. 2011-0030074).
SUPPLEMENTARY MATERIAL
Supplementary Data S1