
Abstract
The aim of this study was to investigate the effects of krill oil (FJH-KO) in monoiodoacetate (MIA)-induced osteoarthritis in rat models, and H2O2- or lipopolysaccharide (LPS)-treated primary chondrocytes and the SW982 synovial cell line. We found that 150 mg/kg b.w. FJH-KO supplementation increased running speed, stride, and foot pressure in MIA-induced osteoarthritic rats. In the H2O2-treated SW982 synovial cell line and primary chondrocytes, FJH-KO treatment prevented cell death and suppressed matrix degradation by increasing the levels of anabolic factors of cartilage tissue, including aggrecan, collagen type Ⅰ, collagen type Ⅱ, tissue inhibitors of metalloproteinase (TIMP)-1, and TIMP-3, and decreasing those of catabolic factors of cartilage tissue, including phosphorylation of Smad, MMP-3, and MMP-13. In addition, FJH-KO treatment suppressed the activation of inflammation and apoptosis pathways in the LPS-treated SW982 synovial cell line and primary chondrocytes. We suggest that FJH-KO supplementation may help prevent osteoarthritis progression because of its direct effects on inflammation and apoptosis of chondrocytes.
Introduction
Osteoarthritis, commonly known as wear-and-tear arthritis, is a degenerative joint disease that causes cartilage degradation and synovial inflammation, leading to joint pain and stiffness. Factors that may contribute to the development of this disease include age, obesity, musculoskeletal abnormalities, and joint injuries and overuse. However, its exact etiology and mechanism remain unclear. 1 –3 Patients with osteoarthritis face a deterioration in their quality of life due to physical limitations, chronic pain, and poor treatment outcomes, which also affects mental health. 4 Although the prevalence of this disease is expected to increase, medications cannot reverse the joint damage caused by osteoarthritis. 5,6
Degeneration of articular cartilage is directly related to the progressive loss of the extracellular matrix, mainly composed of aggrecan and type II collagen, by an abnormal mechanical stress-induced imbalance between degradation and synthesis. 7 –9 Abnormal mechanical stresses in articular cartilage result in chondrocyte inflammation, apoptosis, and metabolic derangement that cause the degradation of the extracellular matrix through upregulation of metalloprotease, including matrix metalloproteinases (MMPs). 10,11 Several studies have shown that inflammatory mediators, including inflammatory cytokines, prostaglandin E2 (PGE2), and nitric oxide, exacerbate the progression of osteoarthritis. 12,13
Osteoarthritis is usually treated by using a combination of treatments, including medications, physical therapy, injections into the joint, and surgery. Patients with this disease are usually prescribed strong anti-inflammatory drugs or other medications, including acetaminophen (Tylenol) and nonsteroidal anti-inflammatory drugs, to help relieve pain and minimize cartilage damage. 14 Unfortunately, these medications have side-effects such as constipation, diarrhea, heartburn, nausea, vomiting, and dizziness. 15 Therefore, complementary and alternative treatments for the prevention and treatment of osteoarthritis are being investigated. 16
In a previous study, we found that the supplementation with Antarctic krill oil (FJH-KO) suppressed monoiodoacetate (MIA)-induced articular cartilage damage and inflammation in rat models. 17 FJH-KO is extracted from Antarctic krill (Euphausia superba), which are red shrimp-like small crustaceans that are a good source of omega-3 (n-3) long-chain polyunsaturated fatty acids (LC-PUFA). Moreover, the fatty acids in FJH-KO are bound to phospholipids, particularly phosphatidylcholine, which can increase bioavailability of n-3 PUFAs to the body tissues. 18,19
Several animal studies have suggested that FJH-KO supplementation may act as an anti-inflammatory agent; however, the exact mechanism underlying its effects has not been identified. In this study, we investigated the effects of FJH-KO treatment on anabolic and catabolic factors, inflammation, and apoptosis using H2O2- or lipopolysaccharide (LPS)-treated primary chondrocytes and the SW982 synovial cell line.
Materials and Methods
Induction of osteoarthritis in rats and treadmill trials
The experimental protocol was approved by the Institutional Animal Care and Use Committee of Kyung Hee University (KHGASP-19-331). Male Sprague-Dawley rats (6 weeks old) were obtained from Japan SLC, Inc. (Shizuoka, Japan). Rats were induced osteoarthritis and supplied FJH-KO using our previous methods. 17 Three weeks after injecting MIA, we measured running speed, stride, and foot pressure for 10 min using a rat-specific treadmill (Jeollanamdo Institute of Natural Resources Research, Korea).
Primary chondrocytes and SW982 synovial cell line culture and treatment
The isolations of chondrocytes from rats were performed by our previous method. 20 The SW982 synovial cell line was obtained from the American Type Culture Collection (Rockville, MD, USA) and cultured according to the manufacturer's protocol. The Antarctic FJH-KO was supplied by Frombio (Suwon-si, Gyeonggi-do, Republic of Korea). Nutritional composition of FJH-KO is shown in Table 1, and the sum of eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA) in FJH-KO was 204.49 ± 2.72 mg/g (Fig. 1). The extracts were stored at −20°C until used, and FJH-KO stock was dissolved in DMSO for treatment in the cells. Acetylsalicylic acid (10 μM) or diverse concentrations of FJH-KO were incubated for 24 h in the H2O2- 200 μM or LPS 50 μg/mL-treated cells.
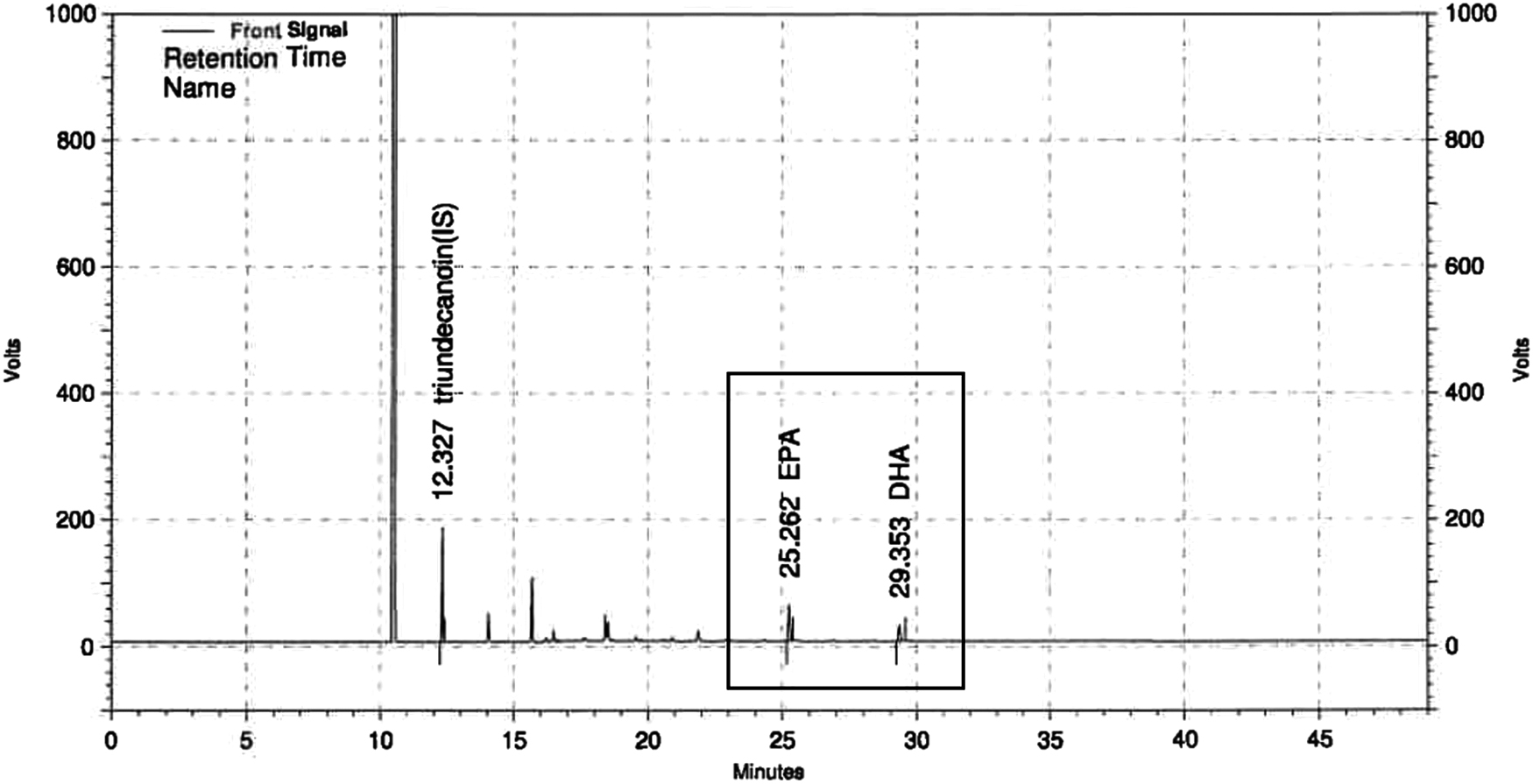
High-performance liquid chromatography analysis of EPA and DHA concentrations in FJH-KO. DHA, docosahexaenoic acid; EPA, eicosapentaenoic acid;
Nutritional Composition of FJH-KO
Cell viability
To evaluate the effect of FJH-KO on viability of the chondrocytes, 3-(4,5-dimethylthiazol-2yl-)-2,5-diphenyl tetrazolium bromide (MTT) was used in the chondrocytes. MTT solution (20 μL, 5 mg/mL in PBS as stock solution) was added to chondrocytes for 3 h at 37°C. To measure the absorbance, supernatants were extracted through suction, and 200 μL of DMSO was added to each well. The plates were read at 560 nm.
Real-time polymerase chain reaction (RT-PCR)
We extracted total RNA from cells, synthesized complementary DNA, and performed real-time polymerase chain reaction (RT-PCR) according to a previously published method 17 using specific primers (Table 2).
Primer Sequences Used in Real-Time Polymerase Chain Reaction Quantification of mRNA
MMPs, matrix metalloproteinases.
Measurement of cytokines, PGE2, and nitric oxide levels
The levels of interleukin-1β (IL-1β), IL-6, tumor necrosis factor-α (TNF-α), and PGE2 were measured using an ELISA kit (R&D Systems, Minneapolis, MN, USA). The concentration of nitric oxide in the serum was measured using the NO ELISA Kit (ENZO Life Sciences, PA, USA) according to the manufacturer's protocol.
Western blot analysis
Total protein from cells was dissolved in 4 × NuPAGE® LDS sample buffer (Life Technologies, Gaithersburg, MD, USA). Protein samples containing 100 μg protein from cells were separated by gel electrophoresis and transferred onto membranes. Membranes blocking, incubation with primary antibodies and secondary antibody, visualization and analysis were performed by our previous methods. 21
Statistical analysis
All data are expressed as means ± standard deviations (SDs); differences among groups were evaluated using one-way analysis of variance and Duncan's multiple range tests implemented using SPSS for Windows (SPSS PASW Statistic 22.0; SPSS, Inc., Chicago, IL, USA). Differences were considered significant at P < .05.
Results
FJH-KO suppressed pain levels in MIA-injected rats
We measured running speed, stride, and foot pressure of rats with MIA-induced osteoarthritis using the treadmill to investigate the effect of FJH-KO on pain levels. The levels of speed, stride, and foot pressure were significantly lower in MIA-injected rats (C) than those in normal rats (NC). However, dietary supplementation with FJH-KO significantly increased speed, stride, and foot pressure compared with those of the MIA-injected rats (P < .05) (Fig. 2).

The effect of FJH-KO on running speed
FJH-KO ameliorated H2O2-induced cell damage
Cell death was induced by treatment with 200 μM H2O2 in the SW982 synovial cell line and primary chondrocytes. FJH-KO pretreatment ameliorated H2O2-induced cell death in both SW982 synovial cell line and primary chondrocytes (P < .05) (Fig. 3).
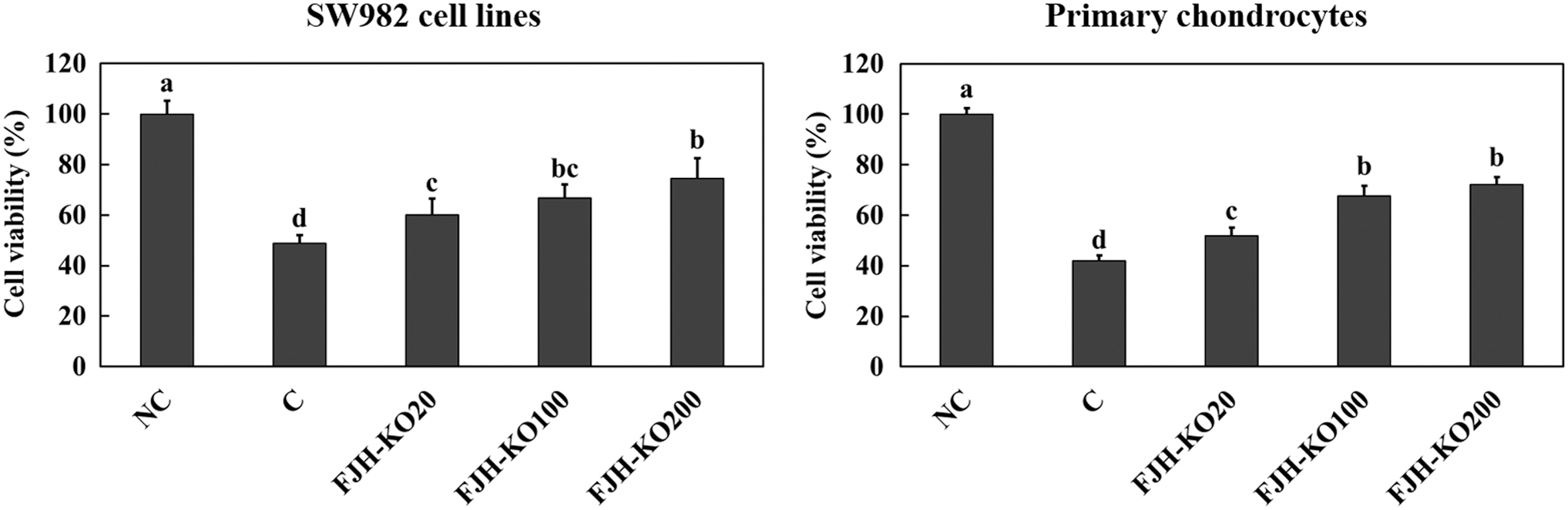
The effect of FJH-KO on H2O2-induced damage of SW982 synovial cells and primary chondrocytes. NC: no treatment; C: H2O2 200 μM treatment; FJH-KO20: H2O2 200 μM and FJH-KO 20 μg/mL treatment; FJH-KO100: H2O2 200 μM and FJH-KO 100 μg/mL treatment; and FJH-KO200: H2O2 200 μM and FJH-KO 200 μg/mL treatment. Values are presented as means ± SDs. Different letters (a>b>c>d) indicate a significant difference with P < .05, as determined using Duncan's multiple range test.
FJH-KO ameliorated H2O2-induced anabolic factors mRNA expression changes
mRNA expression levels of the anabolic factors, aggrecan (Fig. 4A, F), collagen type Ⅰ (Fig. 4B, G), collagen type Ⅱ (Fig. 4C, H), TIMP-1 (Fig. 4D, I), and TIMP-3 (Fig. 4E, J) were significantly decreased in the H2O2-treated SW982 synovial cell line and primary chondrocytes as compared with those in the normal group (NC). However, acetylsalicylic acid (PC) or FJH-KO treatment significantly increased mRNA expression levels of the anabolic factors in H2O2-treated SW982 synovial cell line and primary chondrocytes (P < .05).

The effect of FJH-KO on H2O2-induced changes in the mRNA expression of aggrecan
FJH-KO ameliorated H2O2-induced catabolic factors expression changes
The mRNA and protein expression levels of catabolic factors, MMP-3 and MMP-13, were significantly increased in the H2O2-treated SW982 synovial cell line and primary chondrocytes compared with those in the normal group (NC). In addition, phosphorylation of Smad, an MMP regulator, was significantly lower in the H2O2-treated SW982 synovial cell line and primary chondrocytes compared with that in the normal group. Treatment of acetylsalicylic acid (PC) or FJH-KO in the H2O2-treated SW982 synovial cell line and primary chondrocytes caused an increase in the mRNA and protein expression levels of MMP-3 and MMP-13, and a decrease in the protein expression level of phosphorylation of Smad compared with those in the control group (C) (Fig. 5).
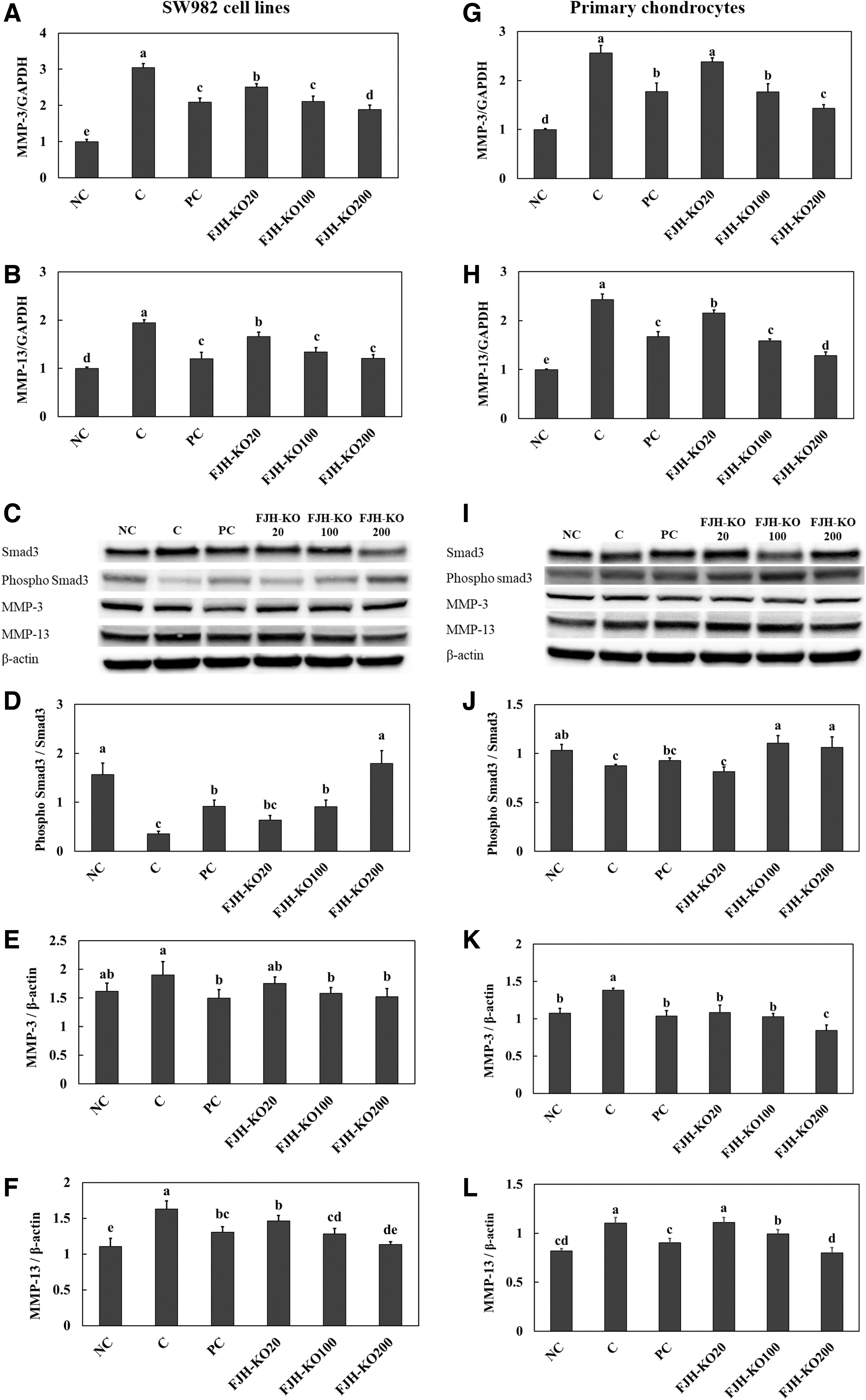
The effect of FJH-KO on H2O2-induced changes in expression changes of catabolic factors, mRNA of MMP-3
FJH-KO ameliorated LPS-induced inflammation
We found that treatment with LPS stimulated secretion of proinflammatory cytokines (IL-1β, IL-6, and TNF-α), PGE2, and nitric oxide in LPS-treated SW982 synovial cell line and primary chondrocytes. However, acetylsalicylic acid (PC) or FJH-KO treatment suppressed secretion of inflammatory molecules in LPS-treated SW982 synovial cell line and primary chondrocytes (Fig. 6) (P < .05).
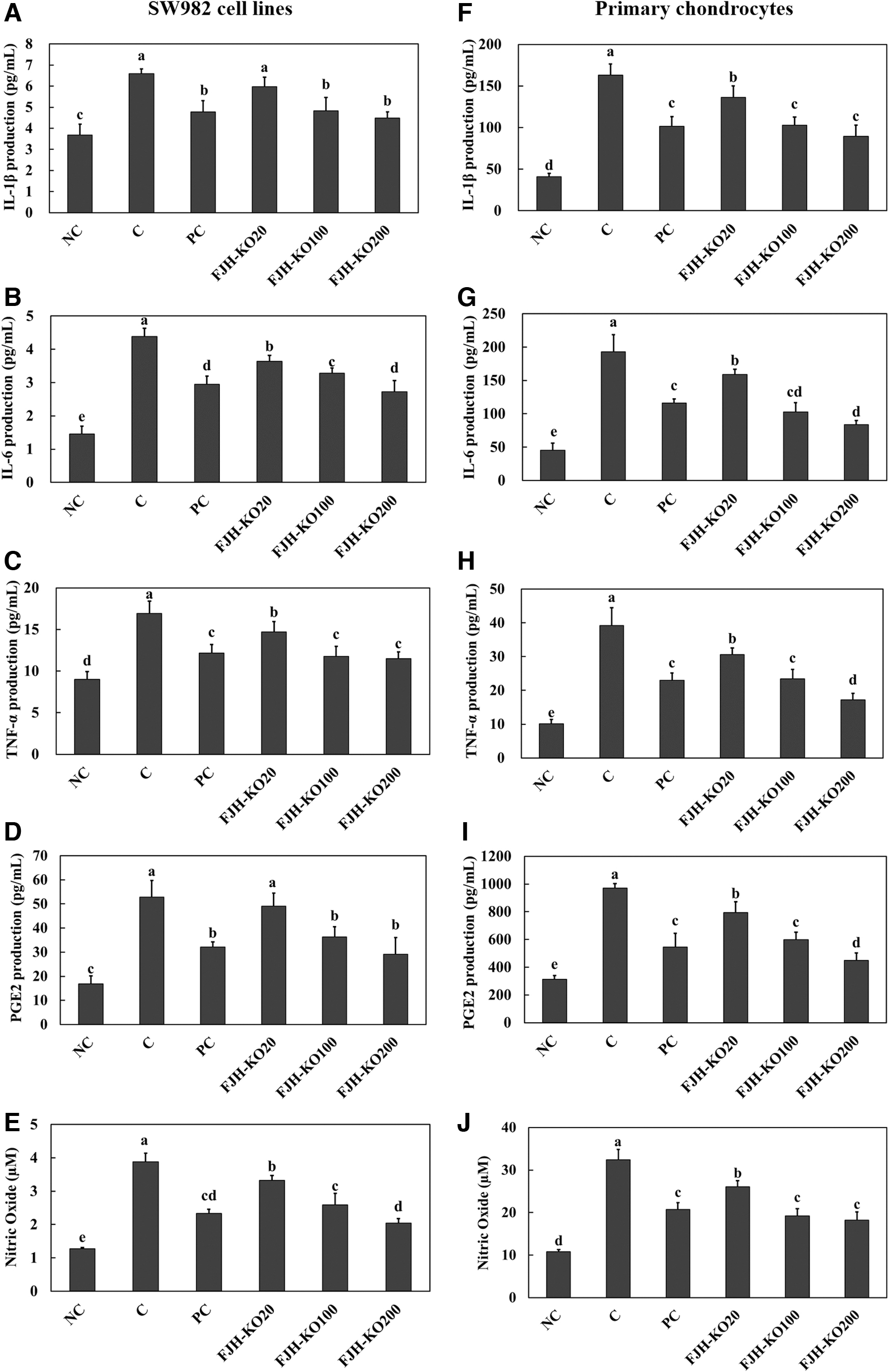
The effect of FJH-KO on LPS-induced secretion of inflammatory mediators, IL-1β
In addition, LPS treatment increased protein expression levels of phosphorylation of IκB (key regulators of the nuclear factor kappa-light-chain-enhancer of activated B cells [NF-κB] pathway), p65 (nuclear factor NF-κB p65 subunit), and cyclooxygenase-2 (COX-2) in both SW982 synovial cell line and primary chondrocytes. Acetylsalicylic acid (PC) or FJH-KO treatment ameliorated LPS-induced protein expression levels of phosphorylation of IκB and p65 and COX-2 (Fig. 7).

The effect of FJH-KO on LPS-induced activation of inflammatory pathways in SW982 synovial cell line [band images
FJH-KO ameliorated LPS-induced apoptosis
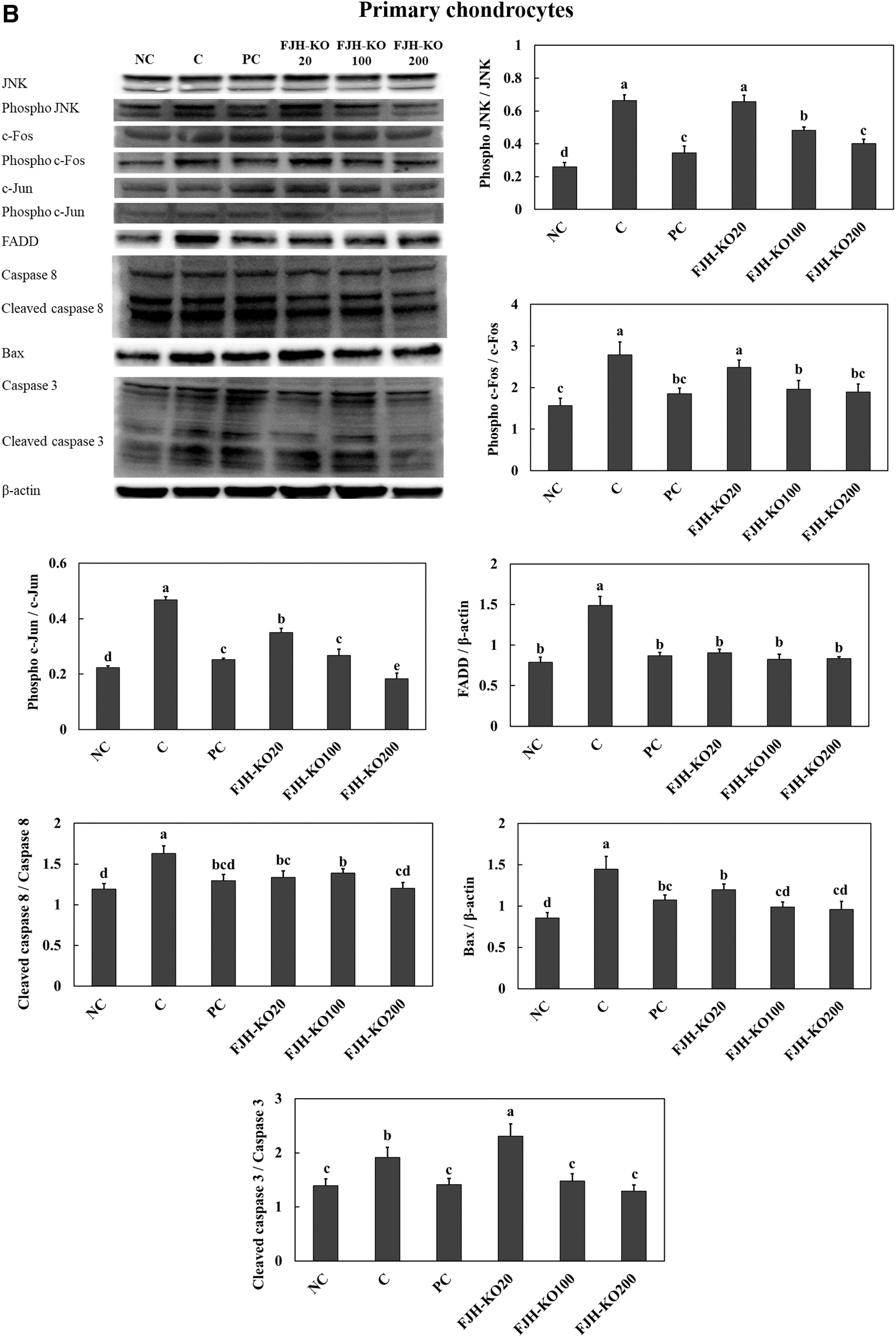
Treatment with LPS stimulated apoptosis signaling pathways, including the c-Jun N-terminal kinase (JNK)/c-Fos and c-Jun pathway and FADD/caspase8/Bax/caspase3 pathway in both SW982 synovial cell line and primary chondrocytes. Treatment with acetylsalicylic acid (PC) or FJH-KO in LPS-induced cells suppressed the expression of proteins involved in the apoptosis pathways in both SW982 synovial cell line (Fig. 8A) and primary chondrocytes (Fig. 8B).
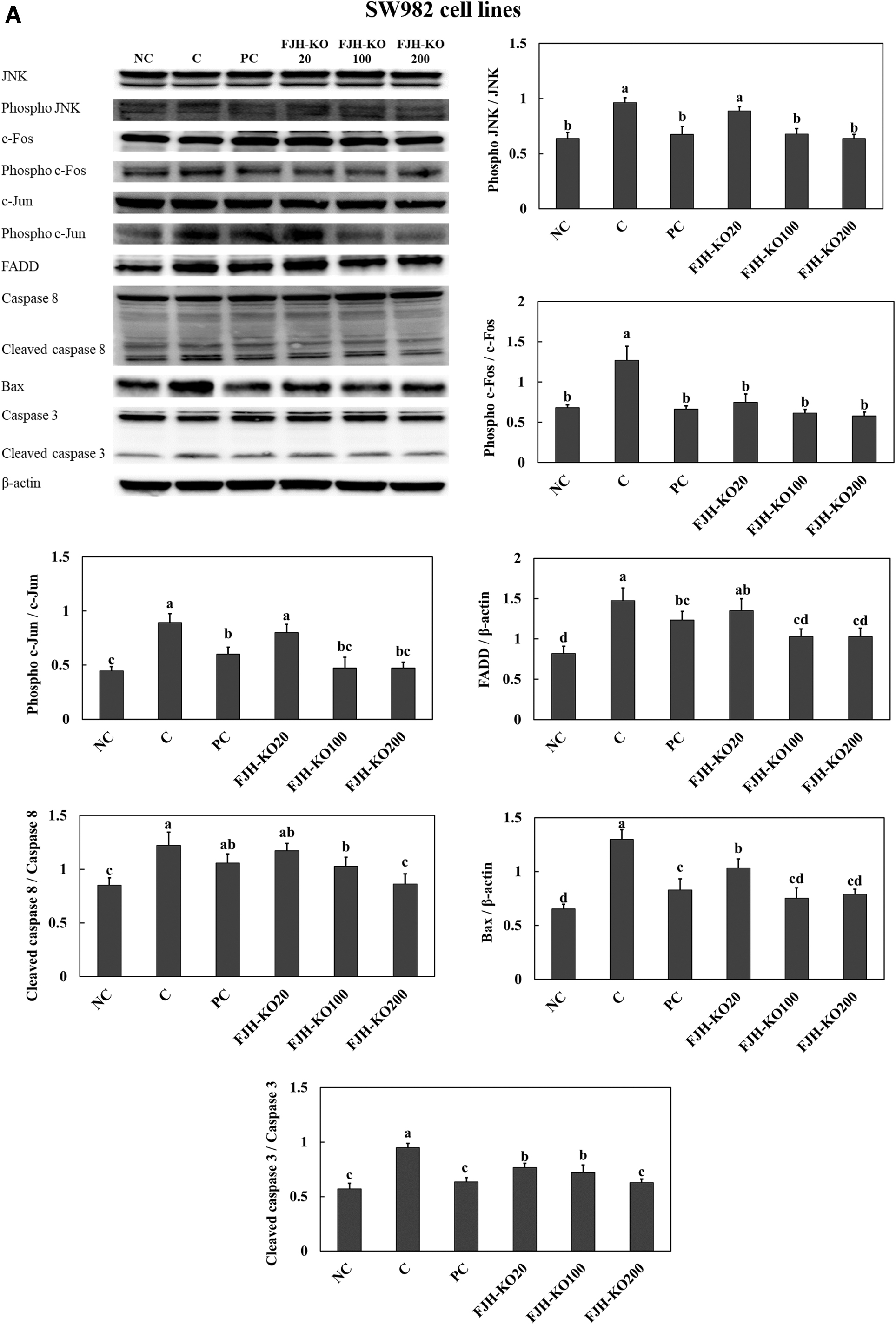
The effect of FJH-KO on LPS-induced activation of the apoptosis pathway in the SW982 synovial cell line
Discussion
After a study by Illingworth et al., which reported on the effect of n-3 LC-PUFA derived from fish oils in reducing LDL-cholesterol, the health benefits of n-3 LC-PUFA gained immense attention in the scientific community. 22,23 Recently, there has been increasing interest in the effect of FJH-KO containing abundant n-3 LC-PUFA. 24 Kim et al. demonstrated that ozonated FJH-KO suppressed proinflammatory mediators in LPS-treated macrophages. 25 In this study, we investigated the beneficial effects of FJH-KO in an H2O2- or LPS-treated SW982 synovial cell line and in primary chondrocytes to provide a scientific basis for utilizing natural agents in osteoarthritis treatment.
Injection of MIA (an inhibitor of glyceraldehyde-3-phosphate dehydrogenase) in articular cartilage induces the development of osteoarthritis caused by cell death. 26 First, we showed that FJH-KO supplementation inhibited the development of osteoarthritis induced by MIA injection in rats using a treadmill assay. Next, the in vitro models in this study used H2O2 and LPS treatment for inducing osteoarthritis-like inflammation in SW982 synovial cell line and primary chondrocytes.
We found that FJH-KO treatment suppressed degradation of the matrix in H2O2-induced chondrocyte through increases in anabolic factors of cartilage tissue, including aggrecan, collagen type Ⅰ, collagen type Ⅱ, TIMP-1, and TIMP-3, and decreases in catabolic factors of cartilage tissue, including phosphorylation of Smad, MMP-3, and MMP-13. Numerous reports suggest that osteoarthritis with degradation of extracellular matrix is associated with inflammation and cell death. 27,28 Thus, we investigated whether FJH-KO treatment could inhibit the inflammation and apoptosis in an LPS-treated SW982 synovial cell line and primary chondrocytes. FJH-KO treatment in LPS-treated cells attenuated the activation of inflammation and apoptosis pathways.
The p65 subunit of NF-κB is known to be activated in response to cytokine stimulation, and is essential for inducing inflammation. NF-kB activation occurs when it is released from IκB and subsequently transferred from the cytoplasm to the nucleus. Regulation of gene expression by NF-κB controls the expression of numerous genes involved in inflammatory and cellular proliferation, and has been found to be constitutively activated in osteoarthritis-like inflammation. 27,29 In this study, we showed that FJH-KO treatment suppressed IκB and p65 phosphorylation in the LPS-treated SW982 synovial cell line and in primary chondrocytes. These data suggest that FJH-KO treatment prevented inflammation in synovial cells and chondrocytes through inhibition of NF-κB signaling pathways.
In addition, FJH-KO treatment suppressed the apoptosis-involved JNK/c-Fos and c-Jun and FADD/caspase8/Bax/caspase3 pathways in the LPS-treated SW982 synovial cell line and in primary chondrocytes. Hwang and Kim reported that degradation of the extracellular matrix in the pathogenesis of osteoarthritis can be caused by apoptosis-induced chondrocytes death.
In addition, they showed that reactive oxygen species, nitric oxide, proinflammatory cytokines, and mechanical stress are involved in chondrocytes apoptosis inducers and stimulate caspases (key molecules in the apoptosis). 28 Therefore, chondrocyte apoptosis is a potential target for pharmacological treatment of osteoarthritis.
FJH-KO supplementation suppressed MIA-induced development of irregular articular cartilage surface and cartilage matrix degradation in the animal model of our previous study. Thus, we suggest that FJH-KO supplementation can prevent the development of osteoarthritis through suppression of apoptosis in chondrocytes. Although there are various causes of osteoarthritis, our previous 17 and present studies used an animal model in which osteoarthritis was forcibly induced by using MIA. Therefore, we need to further confirm the efficacy of FJH-KO in various osteoarthritis models, such as aging, and in females.
In summary, our previous and present studies indicate that natural FJH-KO supplementation has a beneficial effect in the treatment of osteoarthritis as it inhibits inflammation and apoptosis in synovial cells and chondrocytes. This study provides scientific evidence for, and describes the mechanisms underlying anti-inflammatory and anti-osteoarthritis effects of FJH-KO.
Footnotes
Acknowledgment
This research was a part of the project titled “Development and commercialization of functional materials on improving joint health using krill oil”, funded by the Ministry of Oceans and Fisheries, Korea.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This research was funded by the Ministry of Oceans and Fisheries, Korea.